Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Neural engineering wikipedia , lookup
Membrane potential wikipedia , lookup
Neuroplasticity wikipedia , lookup
Action potential wikipedia , lookup
Resting potential wikipedia , lookup
Electroencephalography wikipedia , lookup
Haemodynamic response wikipedia , lookup
End-plate potential wikipedia , lookup
Central pattern generator wikipedia , lookup
Neural coding wikipedia , lookup
Biological neuron model wikipedia , lookup
Activity-dependent plasticity wikipedia , lookup
Nonsynaptic plasticity wikipedia , lookup
Subventricular zone wikipedia , lookup
Holonomic brain theory wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Magnetoencephalography wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Synaptogenesis wikipedia , lookup
Development of the nervous system wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Multielectrode array wikipedia , lookup
Evoked potential wikipedia , lookup
Nervous system network models wikipedia , lookup
Neuroanatomy wikipedia , lookup
Neural oscillation wikipedia , lookup
Chemical synapse wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Pre-Bötzinger complex wikipedia , lookup
Synaptic gating wikipedia , lookup
Optogenetics wikipedia , lookup
Apical dendrite wikipedia , lookup
Metastability in the brain wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Single-unit recording wikipedia , lookup
Electrophysiology wikipedia , lookup
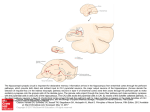
Cellular-synaptic generation of EEG activity György Buzsáki1, Roger D. Traub2 and Timothy Pedley3 1Center for Molecular and Behavioral Neuroscience, Rutgers, The State University of New Jersey, 197 University Avenue, Newark, NJ 07102 2 Department of Physiology, Neuroscience Unit Medical School, University of Birmingham Birmingham B15 2TT, U.K. and 3Department of Neurology Columbia University, New York, NY 10032 Correspondence: György Buzsáki Center for Molecular and Behavioral Neuroscience, Rutgers University 197 University Avenue Newark, NJ 07102 Tel: (973) 353-1080 ext. 3131 Fax: (973) 353-1588 E-mail: [email protected] Key words: EEG, cellular activity, synchrony, extracellular currents, intrinsic oscillations, synaptic activity, current-source density analysis Supported by NIH (NS34994, MH54671) and the Wellcome Trust. 1 Introduction To date, three methods can provide high temporal resolution of neuronal interactions at the network level: electric field recording (EEG), magnetoencephalogram (MEG; 51, 70) and optical imaging (32, 86). Each of these have their advantages and shortcomings. MEG is not practical for experimental work on freely moving subjects due to the large size of magnetic sensors. A major obstacle of the optical imaging method is that its "view" is confined to surface events. Since most of the network interactions occur in the depth of the brain at the level of the synapses, a search for alternative methods is warranted. In addition, research in both MEG and optical imaging fields face the same fundamental questions as those arose decades ago in connection with scalp-recorded EEG: the "reverse engineering" problem of signal interpretation (cf. 10, 31, 63). Membrane currents generated by neurons pass through the extracellular space. These currents can be measured by electrodes placed outside the neurons. The field potential (i. e., local mean field), recorded at any given site, reflects the linear sum of numerous overlapping fields generated by current sources (current from the intracellular space to the extracellular space) and sinks (current from the extracellular space to the intracellular space) distributed along multiple cells. This macroscopic state variable can be recorded with electrodes as a field potential or electroencephalogram (EEG) or with magnetosensors (SQUIDs) as a magnetoencephalogram (MEG). These local field patterns, therefore, provide experimental access to the spatio-temporal activity of afferent, associational and local operations in a given structure. To date, field potential measurements provide the best experimental and clinical tool for assessing cooperative neuronal activity at high temporal resolution. However, without a mechanistic description of the underlying neuronal processes, the scalp or depth EEG simply remains a gross correlate of brain activity rather than a predictive descriptor of the specific functional/anatomic events. The essential experimental tools for the exploration 2 of EEG generation have yet to come. In this chapter we provide a basic description of field potential generation in the mammalian archicortex and neocortex and summarize recent progress and future directions. A straightforward approach to deconvolve the surface (scalp) recorded event is to simultaneously study electrical activity on the surface and at the sites of the extracellular current generation. Electrical recording from deep brain structures using wire electrodes is one of the oldest methods in neuroscience. Local field potential measurements or "micro-EEG" (66) combined with recording of neuronal discharges is the best experimental tool available to study the influence of cytoarchitectural properties, such as cortical lamination, distribution, size and network connectivity of neural elements on electrogenesis. However, large number of observation points combined with decreased distance between the recording sites are required for high spatial resolution and for making interpretation of the underlying cellular events possible. Progress in this field is expected to be accelerated by the availability of micromachined silicon-based probes with numerous recording sites (60). The information obtained from the depth of the brain will then help interpret the surfacerecorded events. Clearly, such a task requires collaborative work among neuroscience, silicon nanotechnology, micromachinery, electric engineering, mathematics and computer science. The stake is high, since interpretation of macrosignals such as those obtained with EEG, MEG, fast MRI, PET or optical imaging methods will still require network (submillimeter) level interpretation of the cellular- synapic interactions. In principle, every event associated with membrane potential changes of individual cells (neurons and glia) should contribute to the perpetual voltage variability of the extracellular space. Until recently, synaptic activity has been viewed as the exclusive source of extracellular current flow or EEG. As will be discussed below, however, synaptic activity is only one of the several membrane voltage changes that contributes to the measured field potential. Progress during the past decade revealed numerous 3 sources of relatively slow membrane potential fluctuations, not directly associated with synaptic activity. Such non-synaptic events may contribute also significantly to the generation of local field potentials. These include calcium spikes, voltage-dependent oscillations and spike afterpotentials observed in various neurons. Sources of extracellular current flow 1. Fast (Na+) action potentials The largest amplitude intracellular event is the sodium-potassium spike, referred to as the fast (Na+) action potential intracellularly and as unit activity extracellularly. Individual fast action potentials are usually not considered to contribute significantly to the scalp recorded EEG, mainly because of their short duration (<2 msec). An additional factor is the high-pass frequency filtering (capacitive) property of the extracellular medium, which attenuates spatial summation of high frequency events. As a result, the amplitude of extracellular unit activity decreases much more rapidly with distance between the cell membrane and the recording site than is the case for slower membrane events. However, when a microelectode is placed close to the cell body layer of cortical structures the recorded field potentials contain both extracellular units and summed synaptic potentials. Furthermore, when action potentials from a large number of neighboring neurons occur within a short time window, e. g., in response to electrical stimulation of afferents, during epileptic activity or even during synchronous physiological patterns, these "population spikes" can be recorded with relatively large size electrodes and in a larger volume (Fig. 1; refs. 4, 9, 25). 2. Synaptic activity In most physiological situations, synaptic activity is clearly the most significant source of extracellular current flow or EEG. The notion that synaptic potentials contribute to 4 the generation of EEG stems from the recognition that for the summation of extracellular currents from numerous individual compartments, the events must be relatively slow (39). The dendrites and soma of a neuron form a tree made of an electrically conducting interior surrounded by a relatively insulating membrane with tens of thousands of synapses on it. Each synapse acts as a small battery to drive current, always in a closed loop. Depending on the chemical nature of the neurotransmitter released in the synaptic cleft, the postsynaptic membrane is depolarized (EPSP) or hyperpolarized (IPSP). Excitatory currents, involving Na+ or Ca2+ ions, flow inwardly at an excitatory synapse (i. e., from the activated postsynaptic site to the other parts of the cell) and outwardly away from it. Such an outward current is referred to as a passive return current from the intracellular milieu to the extracellular space. Inhibitory loop currents, involving Cl- or K+ ions, flow in the opposite direction. The current flowing across the external resistance of the cortex sums with the loop currents of neighboring neurons to constitute a local mean field (Fig. 1). Viewed from the perspective of the extracellular space, membrane areas where current flows into or out of the cells are termed sinks or sources, respectively. The active or passive nature of the sinks and sources are ambiguous, unless the location and types of synapses, involved in the current generation, are identified. Supplementary information may come from simultaneous intracellular recording from neurons dominantly involved in the current generation. Alternatively, extracellular recording of the action potentials and their cross-correlation with the laminar distribution of the field event can provide the necessary clues for the identification of an sink as opposed to a passive return (inward) current of an active inhibitory source (outward). Cross-correlation of the interneuronal discharges with the field potential in question may further decrease the ambiguity regarding the passive versus active nature of the sink-source dipole (16). Identification of synaptic currents in the archicortex 5 Figure 1 illustrates the necessary steps in the identification of network mechanisms of evoked and spontaneous field events. The example is taken from the hippocampus, because it is a simple, three-layered structure consisting of orderly arranged principal cells (pyramidal and granule cells) and interneurons. Therefore, the synaptic interpretation of the extracellular current is much simpler than in multilayered structures. The termination zones of the excitatory paths and the inhibitory connections are also well studied in the hippocampus (10, 84). Activation of the excitatory associational input by indirect, trisynaptic electrical stimulation will depolarize the midapical and basal dendrites of pyramidal cells (shown in blue in Fig. 1). The passive return current will flow out of the cells at the level of the neuronal bodies and distal apical dendrites (shown in red in Fig. 1). This change in voltage is reflected by the characteristic distribution of field potentials in different depths. The extracellular voltage is negative close to the excitatory synapse and positive in the cell body layer. The reason for this is the large depolarization of the dendrite and the gradual decrease of intracellular depolarization towards to soma. This synaptic activity-induced intracellular voltage difference between the dendrites and soma (a "dipole") will result in a current flow across the membrane (arrows in Fig. 1F). Simultaneous events in many neighboring pyramidal cells will linearly summate and produce an extracellular voltage fluctuation which can be measured with closely-spaced electrodes. After determining the impedance characteristics of the extracellular space, the voltage change can be converted into current change (28). Increased afferent discharge also activates interneurons, some of which terminate on the cell bodies of the pyramidal cells. The discharging basket cells release GABA and activate Cl- channels with resulting hyperpolarization of the pyramidal cell somata. Somatic hyperpolarization, in turn, creates a voltage gradient between the soma and dendrites (inhibitory dipole). The created intracellular voltage difference is the driving force of charges across the cell membrane and the consequent spatially distributed current flow in the surrounding extracellular fluid (Fig. 1). Note that the direction of 6 current flow is the same as in the case when the driving force is apical dendritic depolarization (active sink). Since the direction of current flow is identical for dendritic excitation and somatic inhibition, the excitatory and inhibitory currents will sum in the extracellular space, resulting in large amplitude field potentials. The contribution of GABAa-mediated inhibitory currents, however, is believed to be small, because the Cl- equilibrium potential is close to the resting membrane potential. Thus, the change of the transmembrane voltage is limited. However, in actively spiking neurons, when the cell body is depolarized, the transmembrane potential, mediated by GABAa synapses can be large. Another cautionary note is that inhibition may operate also on the dendrites, causing current flow opposite to the direction of excitatory currents. For the identification of excitatory and inhibitory components, represented by the extracellular current flow, a precise knowledge about the anatomical network is essential. Physiological experiments, including recordings from interneurons and pyramidal cells as well as differential pharmacological blockage of the excitatory and inhibitory synapses, can then provide the necessary knowledge for the proper interpretation of the observed sinks and sources. When all this knowledge is in place, the extracellular events can be interpreted unambiguously. Provided that dendritic excitation is strong enough to override somatic inhibition, the cells may discharge. In the simplest case, a Na+ spike will be generated in the initial segment of several pyramidal neurons. The large inward current associated with the spike is reflected by a negative change of the extracellular voltage at the level of the axon initial segment/cell body accompanied by a smaller amplitude, extracellular positive deflection out in the dendritic regions for the same reasons as described above for the EPSPs. However, since the spatial location of this event is opposite to the afferent excitation of dendrites, the direction of the extracellular current flow will also be opposite. The contribution of fast spikes to the extracellular mean field is due to the hypersynchronous discharge of many pyramidal neurons (population spike) as a result 7 of artificial stimulation of an afferent bundle. Interpretation of the extracellular events after the population spike is not straightforward, however, due to complex feedback effects of the network and other non-synaptic events (see below). Once a circuitry, such as shown in Fig. 1, has been "calibrated" by electrically evoked potentials, one can move to the next step: network level description of the generation of spontaneous EEG events. The tutorial example is an intermittently occurring, large amplitude hippocampal sharp wave (SPW). SPWs are present during immobility, consummatory behaviors and slow wave sleep. It is important that the events to be analyzed are clearly separable from other waveforms. After extracting the invariable features of this EEG pattern by averaging or other pattern recognition methods, the simultaneous voltage measurements are converted into a current-source density map (Fig. 1D and E). Note that the distribution of the sinks and sources of SPW is strikingly similar to the potentials elicited by stimulation of the associational/commissural inputs to the pyramidal cells. Indeed, experimental work revealed that SPW in the hippocampus arise from the quasi-synchronous discharge of CA3 pyramidal neurons, the source of associational and commissural afferents to the CA1 region (9, 13). Temporally overlapping activation of converging activity on single CA1 pyramidal cells results in a large depolarization of the dendrites, similar to the depolarization of these cell when the associational pathways are electrically activated. These extra- and intracellular events therefore provide circumstantial evidence that the same neuronal machinery is activated during spontaneously occurring SPWs as during electrical stimulation of the associational afferents. Identification of synaptic currents in the neocortex The strategy described above is, in principle, applicable to any other a priori identified rhythmic or sparse EEG event. Complications arise when several dipoles are involved 8 in the generation of the same EEG patterns, especially when these dipoles are phaseshifted, as is the case in the generation of numerous neocortical patterns (11, 80, 81). Of the neocortical EEG patterns, two conspicuous low-frequency (<15 Hz) rhythms, the physiological sleep spindles and spike-and-wave discharges, associated with petit mal epilepsy, have been studied most extensively (11, 14, 44, 55, 77, 80). It is widely accepted that the source of rhythm generation for both patterns is the interplay between the GABAergic reticular nucleus and corticopetal nuclei of the thalamus (11, 14, 79, 80). It is less clear, however, whether synaptic currents of the thalamocortical afferents can fully account for these rhythms or whether intracortical circuitries are significantly involved in their generation (40). Initially, the "recruiting" response, evoked by repetitive stimulation of intralaminar thalamic nuclei, was thought to be the evoked equivalent of spontaneous spindle waves and spike-and-wave patterns (19, 24, 41, 59, 67). Subsequent studies, however, have suggested that spindle waves are more similar to the "augmenting" response; a pattern evoked by repetitive simulation of sensorimotor thalamic nuclei (58, 75, 76). From the point of EEG generation, this distinction is important since recruiting and augmenting responses have different voltage-versusdepth profiles in the cortex. Thus, a critical issue is the identity of synapses and neurons involved in the generation of these rhythmic patterns. If the thalamocortical synapses are the major source of the extracellular synaptic current then the major sinks are expected to correspond to the anatomical targets of the corticopetal thalamic fibers. Using the approach described above for the hippocampus helped clarify these issues (Fig. 2; 44). The most striking aspects of the experiment shown is Fig. 2 is the general similarity of the spontaneous and evoked field events, independent of the initiating conditions. The spatial position of the major current sinks are sources are similar, independent which thalamic nucleus is being stimulated or which hemisphere. The differences are expressed mainly in the latencies of the large sink-source pairs. Therefore, the similar spatio-temporal distribution of the main sinks and sources 9 suggest that the major current flow derives from the activity of the intracortical circuitry. The neocortex, in essence, functions as a powerful amplifier during these oscillatory events. Because the thalamocortical network is in a metastable state during reduced activities of the brainstem and basal forebrain (55; 77), a weak thalamic or callosal input is capable of recruiting a large population of intracortical neurons. The triggering input may even remain undetectable in the field and the spread of activity reflects primarily the connectivity and excitability of the cortical circuitry rather than the nature of the initiating input (16, 17). The CSD map and the associated multiple-site unit analysis also revealed that at least three dipoles were involved in the generation of the rhythmic field events (Fig. 3; 44). The most consistent dipole was characterized by a major sink in layer IV (dipole 2). When a surface-positive field component was present, it was associated with a major sink in layers V-VI and a source in layers II-III (dipole 1). The third, delayed dipole was represented by a surface-negative spike component and a corresponding sink in layers II-III (dipole 3). The relative strength of these respective sinks varied within single episodes of HVS (Fig. 3). Although the numerous cell types and the complexity of the intracortical circuitry makes identification of the cellular-synaptic origin of neocortical EEG less accessible, these recent findings indicate that the use of simultaneous recording of field and unit activity is a proper method for the revelation of the synapticcellular mechanisms of extracellular current flow in the neocortex. 3. Calcium spikes Beside the fast Na+ spike, an important non-synaptic event in neurons is a wide Ca2+mediated action potential. These Ca2+ spikes are generated in the dendrites and do not propagate to the soma (89). Their major role is believed to boost synaptic inputs and assist plastic modification of synapses (42, 53, 54, 91). Ca2+ spikes represent an inward dendritic current and are large in amplitude (20 to 50 mV). They can occur 10 synchronously with dendritic EPSPs and for this reason they cannot be simply revealed or separated from EPSPs with extracellular recordings. Because Ca2+ spikes are activated by a voltage-dependent mechanism, intradendritic depolarization can trigger them. Figure 4 illustrates in vivo recording from a distal dendrite of a hippocampal CA1 pyramidal neuron during theta oscillation (43). As the dendrite is progressively depolarized by intracellular current injection, the rhythmic synaptic potentials are superimposed by large amplitude Ca2+ events. Are such Ca2+ spikes triggered by physiological stimuli? Recent evidence indicates that it may well be the case. Patterned stimulation of the visual system evoked putative Ca2+ events in layer V pyramidal neurons of area 17 (37). Furthermore, intradendritic recordings during spontaneous SPW bursts revealed that the amount of depolarization, brought about by the converging active presynaptic afferents to CA1 pyramidal cells, is sufficient to trigger voltage-dependent Ca2+ spikes (42). This new information, of course, indicates the need for the reinterpretation of the extracellular events illustrated in Fig. 1. Provided that Ca2+ spikes occur simultaneously in several neurons near the recording electrodes, these large inward currents can significantly contribute to the field sinks observed in the dendritic layers. To date, the quantitative contribution of dendritic Ca2+ spikes to the field EEG has not been determined. They may be quite important in highly synchronous events, such as epilepsy, because synchronous Ca2+ spikes in neighboring neurons may be reflected in the field as large sinks. A complicating factor is that, in contrast to EPSPs, Ca2+ spikes can actively propagate and therefore large dendritic segments and dendritic locations distant from the initiating site, may be also involved. 4. Voltage-dependent intrinsic oscillations 11 Experiments similar to that shown in Fig. 4 revealed that when intradendritic depolarization is sufficiently strong, the resonant property of the membrane may give way to a self-sustained oscillation of the voltage in the theta frequency range, even in the absence of network-driven theta activity. Intrinsic, voltage-dependent slow oscillations and theta frequency resonance have been observed also in somatic recordings of hippocampal pyramidal cells (49), thalamocortical neurons (63), stellate cells of the entorhinal cortex (2) and layer V-VI pyramidal cells of the neocortex (73). In stellate cells, the main driving force of the oscillation is a persistent Na+ current (2), whereas another depolarizing current, (Ih) in conjunction with the low threshold Ca2+ current (IT), is responsible for the maintenance of the cellular rhythm in thalamic neurons (6). Voltage-dependent oscillatory activation of ionic channels has been shown also in the gamma frequency range. The membrane potential of sparsely spiny inhibitory interneurons in cortical layer IV can sustain a 40 Hz oscillation by sequential activation of a persistent sodium current followed by a slowly inactivating K+ conductance (50, 51). Similar intrinsic oscillatory properties have been shown in the intralaminar thalamocortical and GABAergic neurons of the n. reticularis neurons in vivo (80) and in the dendrites of hippocampal pyramidal cells (Penttonen et al., 1988). In most neurons, the voltage-dependent oscillation is subthreshold to trigger action potentials. However, when action potentials do occur, they are phase-locked to the depolarizing portion of the oscillatory cycle. Because these intrinsic, oscillatory membrane fluctuations can occur simultaneously in a number of nearby neurons, their contribution to the extracellular EEG may be substantial. This is perhaps best illustrated in the “low- Ca2+, high Mg2+ “ model of epilepsy, when all synaptic activity is completely blocked and the large rhythmic extracellular field potentials are exclusively due to the voltage-dependent fluctuation of pyramidal cells, coordinated by ephaptic (non-synaptic) transmembrane effects (34). 12 5. Intrinsic spike afterhyperpolarizations: their contribution to cortical delta waves In addition to voltage changes, perturbation of the intracellular concentration of one ion species may trigger influx of other ions by activation of ligand-gated channels. The large Ca2+ influx, in association with a dendritic Ca2+ spike, is followed by the suppression of fast spikes and hyperpolarization of the membrane due to activation of Ca2+-mediated increase of K+ conductance (38, 72). These burst-induced afterhyperpolarizations (AHP) are frequently larger in amplitude and of longer duration than synaptic events. A logical progress of thought is to conclude that they should also be considered as an important source of the extracellularly recorded EEG potential. Large amplitude, slow delta waves (1-4 Hz) are among the most frequently studied neocortical EEG patterns. These irregular, semi-rhythmic or rhythmic patterns are most frequently observed during stage 4 sleep in the normal brain. The rhythmicity of the cortical delta waves is explained by the triggering effect of the periodic quasisynchronous thalamocortical inputs (22, 78). The thalamus can maintain a rhythmic oscillation in the delta range due to the intrinsic properties of thalamocortical neurons and their network connectivity with the GABAergic reticular nucleus (22, 78). In short, the "rhythm generator" of delta waves is the thalamus whereas the "voltage generator" is the neocortex, similar to sleep spindles and spike-and-wave patterns, discussed above. Delta waves occur with largest amplitude in deep (layer V) cortical layers and they are recorded as negative waves on the neocortical surface or the scalp. Depth profile measurements in the neocortex of the cat (15, 40, 71), rabbit (68) and rat (11, 87) revealed that surface negative - deep positive delta waves during slow wave sleep correlate with 13 the suppression or cessation of discharges of layer V pyramidal neurons. Intracellularly, the deep positive waves are correlated with hyperpolarization of pyramidal cells (21). The depth profile of the slow delta waves and the associated unit activity are compatible with the hypothesis that the extracellularly recorded delta waves reflect inhibition of pyramidal cells mediated by GABAergic interneurons (3, 69, 74). GABA released at the somata of layer V pyramidal cells would open the Cl-channels and produce an active outward current whose extracellular spatial summation corresponds to deep positivity. A simultaneously occurring passive inward current at the distal dendrites would set up extracellular (surface) negativity. Indeed, with their widespread action GABAergic interneurons may play an important role in affecting large numbers of pyramidal cells, as discussed earlier. Because subcortical inputs terminate also on GABAergic interneurons (29, 30), the subcortical afferents may globally affect the whole neocortical mantle. A major problem with this `classic` model of delta wave generation is the lack of direct supportive evidence. An explicit prediction of the GABA-interneuron-pyramidal cell model of slow wave generation is that GABAergic cells should fire during the deeppositive delta waves. However, experiments directly addressing this issue failed to find such a correlation in the rat neocortex (11). All putative, physiologically identified, neocortical interneurons decreased their firing rates during the deep-positive slow waves. Although the duration of the GABA effect may outlast the action potentials by tens of msec (23), the effect may be too short for the postulated delta wave-associated GABA-mediated somatic hyperpolarization. GABAB-receptor mediated IPSPs may be a possible candidate. An alternative non-synaptic explanation of the origin of delta wave generation is based on the summation of long-lasting AHPs of layer V pyramidal neurons (11, 77). During sleep, pyramidal cells of the neocortex often fire bursts in response to rhythmic thalamic 14 volleys (22, 78), and these bursts, in turn, can trigger Ca2+-mediated K+-conductance changes. The long-lasting nature of AHP favors the summation of outward somatic currents of individual pyramidal cells resulting in a local positive field in deep layers. Such extracellularly summated currents were hypothesized to form the basis of slow delta EEG waves recorded during sleep (10). The reason why delta waves occur only during slow wave sleep is because subcortical neurotransmitters, such as basal forebrain and brainstem cholinergic neurons, locus coeruleus cells, neurons of the raphe nuclei and hypothalamic histaminergic neurons (1, 5, 11, 33; 77) neurotransmitters are released mostly in the awake brain and the common property of these neurons is to reduce the calcium-mediated potassium conductance (20, 34, 52). These actions of subcortical neurotransmitters at the cellular level therefore result in the blockade of delta waves. Using whole-cell recordings in vivo, Metherate and Ashe (57) could differentiate between IPSPs and AHPs in cortical neurons of the intact brain. First, they showed, by intracellular injection of cesium, that a large part of the delta EEG result from a K+ current. Second, stimulation of the cholinergic nucleus basalis mimicked the cesium effect. Third, cesium injection blocked the nucleus basalis stimulation effect. These findings directly support the suggestion that delta wave-concurrent hyperpolarizations result from the calcium-activated K+ current, rather than by GABAmediated IPSPs. Overall, these examples illustrate that knowledge of the intrinsic properties of the neurons is as important for the identification of sources of the extracellular ion flow as knowledge of synaptic potentials and anatomical circuitry. 6. Other non-synaptic neuronal effects Synchronous discharge of large neuronal populations are often associated with large amplitude extracellular potentials (mV to tens of mV) and steep voltage versus depth gradients. These large field currents, in turn, can influence the activity of nearby neurons by changing their transmembrane voltage (ephaptic effects). Measurement of transmembrane potential changes (as opposed to potentials relative to a distant ground) 15 indicated that such extracellular current loops can depolarize neurons to spike threshold under certain conditions (34, 83). Computer simulations of multiple neurons, embedded in a conductive medium, show that such a mechanism is plausible with observed estimates of extracellular resistivity (85). Importantly, the voltage gradient across pyramidal cell bodies during physiological SPWs and especially during epileptic or interictal spikes is larger than experimentally induced voltage gradients that are known to affect cellular excitability. Although direct experimental support is not available yet, one might expect that ephaptic effects could recruit neurons to fire that are otherwise not or not sufficiently activated by synaptic inputs alone (9, 34). 7. Neuron-glia communication The glial syncytium (astrocytes) is connected through gap junctions, which allow the direct spread of current and the diffusion or transport of small molecules. Although the role of concerted changes in membrane potentials of glia in the generation of extracellular current under physiological conditions has not been studied extensively, recent work on neuron-glia interactions indicate that the glial syncytium my contribute to the slow field patterns in an important way. Intercellular coupling through gap junctions is required for both propagating Ca2+ waves and spreading depression (62). The traveling Ca2+ waves, in turn, can trigger calcium influx into neurons (61, 62). The glia-neuron dialogue in vivo may be responsible for postictal depression (8, 26, 35, 36, 48, 82). The increased [K+]o, resulting from intensive neuronal activity during epileptic afterdischarge, may trigger propagating waves in the astrocytic network reflected by the slowly spreading sustained potentials. In turn, astrocytes at the front of the propagating depolarization wave release more K+ (47, 56), resulting in a large depolarization of neurons. The ensuing depolarization block of spike generation contributes to the termination of the afterdischarge and is regarded as the cause of the consequent "postictal depression" of the EEG (8, 82). 16 DC currents or ultraslow change of the extracellular voltage cannot be recorded with conventional EEG devices with high pass-filtered inputs. Nevertheless, the relatively quick changes in the DC level, such as epilepsy associated spreading depression (8), could be identified mistakenly as slow delta or faster “waves“, due to the differential effect of the high pass-filters. Neuron-glia communication may contribute also to physiological EEG patterns. Sensory evoked responses in scalp recordings with DC amplifiers and non-polarizing electrodes often contain reliable and relatively long-lasting DC changes, usually referred to as Bereitschaftpotenzial (46) or contingent negative variation (88). It remains to be revealed whether and to what extent glial depolarization contributes to these evoked patterns. Ultrafast cortical rhythms SPW-associated depolarization of hippocampal CA1 neurons sets into motion a shortlived, dynamic interaction between interneurons and pyramidal cells. The product of this interaction is an oscillatory field potential (ripple) within stratum pyramidale and a phase-locked discharge of the CA1 network at 200 Hz in the rat (12). SPW-related ripples are also present in higher mammals, including humans (7). The specific synaptic currents, mediating the high-frequency oscillation, are largely mediated by rhythmic, synchronized IPSPs near the soma of CA1 neurons. The mechanism by which highly coherent discharge of pyramidal cells is brought about over the entire dorsal hippocampus during the ripple is not understood (18). Three different hypotheses have been advanced for the explanation of the spatial coherence of fast ripples. The first assumes that the CA3 output produces a voltage-dependent fast discharge in the interneurons and that synchronization of the interneurons is mediated by gap junctions (45). A second explanation is based on the reciprocal connections between the interneuronal and pyramidal cell populations. Fast oscillatory discharges in interneurons would, again, be brought about by the ramp-like depolarizing CA3 17 output. Chance discharge of just a few CA1 pyramidal cells within ~1 ms is hypothesized to reset ongoing oscillatory spiking in the target interneurons and generate a short-lived coherent discharge (90). According to the third hypothesis, zerotime lag synchronization of pyramidal neurons is brought about by assumed gap junctions between their axons (25). Fast field oscillations (300-500 Hz) are also present in the neocortex, in particular in association with sleep spindles and spike-and-wave patterns (Fig. 5; 44). The maximum amplitude of the field oscillation occurs in layer V and the ripple waves reverse in phase in the upper part of layer V. The discharge of pyramidal cells are phase-locked to the ripples. The physiological significance of the fast “ripples“ has yet to be clarified. It may be suggested that the fast oscillation of interneurons during a strong network drive provides a dissipative mechanism to decelerate and limit population synchrony of pyramidal cells and to prevent the "all-or-none" discharge of the activated pyramidal cells by protracting the recruitment process and limiting the number of participating neurons. Because conventional EEG devices are limited in their frequency response, these fast events are often impossible to discern reliably from human scalp recordings (see ENA98). In addition, volume conduction of these fast events is quite limited due to the low-pass filtering properties of lipid membranes, as discussed above. Nevertheless, their detection may be of clinical importance because fast oscillatory events may herald the spread and/or termination of epileptic activity (8, 27). 18 References: 1. Abercrombie ED, Jacobs BL. Single-unit response of noradrenergic neurons in the locus coeruleus of freely moving cats I. Acutely presented stressful and nonstressful stimuli. J Neurosci 1987; 7:2837-2843. 2. Alonso A, Llinas RR. Subthreshold Na+-dependent theta-like rhythmicity in stellate cells of entorhinal cortex layer II. Nature 1989; 342:175-177. 3. Amzica F, Steriade M . Electrophysiological correlates of sleep delta waves. Electroencephalogr Clin Neurophysiol 1998; 107:69-83. 4. Andersen P, Bliss TV, Skrede KK . Unit analysis of hippocampal polulation spikes. Exp Brain Res 1971; 13:208-21. 5. Aston-Jones G, Bloom FE. Norepinephrine-containing locus coeruleus neurons in behaving rats exhibit pronounced responses to non-noxious environmental stimuli. J Neurosci 1981; 1:887-900. 6. Bal T, Von Krosigk M, McCormick DA. Synaptic and membrane mechanisms underlying synchronized oscillations in the ferret lateral geniculate nucleus in vitro. J Physiol (Lond) 1995; 483:641-663. 7. Bragin A, Engel J. Jr, Wilson CL, Fried I, Mathern GW. Hippocampal and entorhinal cortex high-frequency oscillations (100-500 Hz) in human epileptic brain and in kainic acid-treated rats with chronic seizures. Epilepsia 1999; 40: 127137. 8. Bragin A, Penttonen M, and Buzsáki G. Termination of epileptic afterdischarge. J 19 Neurosci 1997; 17:2567-2579. 9. Buzsaki G . Hippocampal sharp waves: their origin and significance. Brain Res 1986; 398:242-252. 10. Buzsáki G, Traub RD. Generation of EEG. In: Engel J Jr, Pedley TA, eds. Epilepsy: a comprehensive textbook. Lippincott-Raven Press, 1996. 11. Buzsaki G, Bickford TG, Ponomareff G, Thal LJ, Mandel RJ, Gage FH. Nucleus basalis and thalamic control of neocortical activity in the freely moving rat. J Neurosci 1988; 8:4007-4026. 12. Buzsaki G, Horvath Z, Urioste R, Hetke J, Wise K. High-frequency network oscillation in the hippocampus. Science 1992; 256:1025-1027. 13. Buzsaki G, Leung L , Vanderwolf CH. Cellular bases of hippocampal EEG in the behaving rat. Brain Res Rev 1983; 6:139-171. 14. Buzsáki G, Smith A, Berger S, Fisher LJ, Gage FH. Petit mal epilepsy and Parkinsonian tremor: hypothesis of a common pacemaker. Neurosci 1990; 36:114. 15. Calvet J, Valvet MC, Scherrer J. Etude stratigraphique corticale de l'activite EEG spontanee. Electroencephal and Clin Neurophysiol 1964; 17:109-125. 16. Castro-Alamancos MA, Connors BW. Spatiotemporal properties of short-term plasticity in sensorimotor thalamocortical pathway in the rat. J Neurosci 1996a; 16:2767-2779. 20 17. Castro-Alamancos MA, Connors BW. Short-term plasticity of a thalamocortical pathway dynamically modulated by behavioral state. Science 1996b; 272:274-277. 18. Chrobak J J, Buzsáki G. High-frequency oscillations in the output networks of the hippocampal-entorhinal axis of the freely moving rat. J Neurosci 1996; 16: 30563066. 19. Clare HM, Bishop GH. Potential wave mechanism in cat cortex. Electroenceph Clin Neurophysiol 1956; 8:583-602. 20. Cole AE, Nicoll RA. Characterization of a slow cholinergic postsynaptic potential recorded in vitro from rat hippocampal pyramidal cells. J Physiol (Lond) 1984; 352:173-188. 21. Creutzfeldt O, Watanabe S, Lux HD. Relations between EEG phenomena and potentials of single cortical cells. I. Evoked responses after thalamic and epicortical stimulation. Electroencephal and Clin Neurophysiol 1966; 20: 1-18. 22. Curro Dossi R, Nunez A, Steriade M. Electrophysiology of a slow (0.5-4 Hz) intinsic oscillation of CAT thalamocortical neurones in vivo. J Physiol (Lond) 1992; 447:215-234. 23. De Koninck Y, Mody I. Noise analysis of miniature IPSCs in adult rat brain slices: properties and modulation of synaptic GABA receptor channels. J Neurophysiol 1994; 71:1318-1335. 24. Demsey EW, Morison RS. The mechanism of thalamo-cortical augmentation and repetition. Amer J Physiol 1942; 138:297-308. 21 25. Draguhn A, Traub RD, Schmitz D, Jefferys JG. Electrical coupling underlies highfrequency oscillations in the hippocampus in vitro. Nature 1998; 394:189-192. 26. Fertziger AP, Ranck JB Jr. Potassium accumulation in interstitial space during epileptiform seizures. Exp Neurol 1970; 26:209-218. 27. Fisher RS, Webber WRS, Lesser RP, Arroyo S, Uematsu S. High frequency EEG activity at the start of seizures. J Clin Neurophysiol 1992; 9:441-448. 28. Freeman JA, Nicholson C. Experimental optimization of current source-density technique for anuran cerebellum. J Neurophysiol 1975; 38:369-382. 29. Freund TF, Antal M. GABA-containing neurons in the septum control inhibitory interneurons in the hippocampus. Nature 1988; 336:170-173. 30. Freund TF, Gulyas A, Acsady L, Gorcs T, Toth K. Serotonergic control of the hippocampus via local inhibitory interneurons. Proc Natl Acad Sci (USA) 1990; 87:8501-8505. 31. Gevins AS, Schaffer RE, Doyle JC, Cutillo BA, Tannehill RL, Bressler SL. Shadows of thought: rapidly changing, asymmetric, brain potential patterns of a brief visuomotor task. Science 1983; 220:97-99. 32. Grinvald A, Frostig R, Lieke E, Hidesheim R. Optical Imaging of Neuronal Activity. Physiol Rev 1988; 68:1285-1366. 22 33. Haas HL, Konnerth A. Histamine and noradrenaline decreas calcium-activated potassium conductance in hippocampal pyramidal cells. Nature (London) 1983; 302: 432-434. 34. Haas HL, Jefferys JGR. Low-calcium field burst discharges of CA1 pyramidal neurones in rat hippocampal slices. J Physiol 1984; 354:185-201. 35. Haglund MM, Schwartzkroin PA. Role of Na-K pump potassium regulation and IPSPs in seizures and spreading depression in immature rabbit hippocampal slices. J Neurophysiol 1990; 63:225-239. 36. Heinemann U, Lux HD, Gutnick MJ. Extracellular free calcium and potassium during paroxysmal activity in cerebral cortex of the cat. Exp Brain Res 1977; 27:237-243. 37. Hirsch JA, Alonso JM, Reid RC. Visually evoked calcium action potentials in cat striate cortex. Nature 1995; 378:612-616. 38. Hotson JR, Prince DA. A calcium-activated hyperpolarization follows repetitive firing in hippocampal neurons. J Neurophysiol 1980; 43:409-419. 39. Humphrey DR. Re-analysis of the antidromic cortical response. I. Potentials evoked by stimulation of the isolated pyramidal tract. Electroenceph Clin Neurophysiol 1968; 24:116-129. 40. Jasper H, Stefanis C. Intracellular oscillatory rhythms in pyramidal tract neurones in the cat. Electroencephal and Clin Neurophysiol 1965; 18:541-553. 23 41. Jasper HH, Drooglever-Fortuyn J. Experimental studies on the functional anatomy of petit mal epilepsy. Res Publ Ass Res Nerv Ment Dis 1947; 26:272-298. 42. Kamondi A, Acsády L, Buzsáki G. Dendritic spikes are enhanced by cooperative network activity in the intact hippocampus. J Neurosci 1998; 18:3919-3928. 43. Kamondi A, Acsády L, Wang X-J, Buzsáki G. Theta oscillations in somata and dendrites of hippocampal pyramidal cells in vivo: activity dependent phaseprecession of action potentials. Hippocampus 1998; 8:244-261. 44. Kandel A, Buzsaki G. Cellular-synaptic generation of sleep spindles, spike-andwave discharges and evoked thalamocortical responses in the neocortex of the rat. J Neurosci 1997; 17:6783-6797. 45. Katsumaru H, Kosaka T, Heizmann CW, Hama K. Gap junctions on GABAergic neurons containing the calcium-binding protein parvalbumin in the rat hippocampus (CA1 region). Exp Brain Res 1988; 72:363-370. 46. Kornhuber HH, Becker W, Taumer R, Hoehne O, Iwase K. Cerebral potentials accompanying voluntary movements in man: readiness potential and reafferent potentials. Electroencephalogr Clin Neurophysiol 1969; 26:439. 47. Kuffler SW, Nicholls JG. Neuroglial cells: physiological properties and potassium mediated effect of neuronal activity on the glial membrane potential. Proc R Soc (Lond) 1966; 168:1-21. 48. Leao AAP. Spreading depression of activity in the cerebral cortex. J Neurophysiol 1944; 7:359-390. 24 49. Leung LS, Yim CY. Intrinsic membrane potential oscillations in hippocampal neurons in vitro. Brain Res 1991; 553:261-274. 50. Llinás RR . The intrinsic electrophysiological properties of mammalian neurons: insight into central nervous system. Science 1988; 242:1654-1664. 51. Llinas RR, Ribary U, Joliot M, Wang X-J. Content and context in temporal thalamocortical binding. In: Buzsaki G, Llinas RR, Singer W, Berthoz A, Christen Y, eds. Temporal Coding in the Brain. Springer-Verlag, 1994. 52. Madison DV, Nicoll RA. Actions of noradrenaline recorded intracellularly in rat hippocampal CA1 pyramidal neurons, in vitro. J Physiol (Lond) 1986; 321: 175177. 53. Magee JC, Johnston D. A synaptically controlled, associative signal for Hebbian plasticity in hippocampal neurons. Science 1997; 275:209-213. 54. Markram H, Lübke J, Frotscher M, Sakmann B. Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 1997; 275:213-215. 55. McCormick DA. Neurotransmitter actions in the thalamus and cerebral cortex and their role in neuromodulation of thalamocortical activity. Prog Neurobiol 1992; 39:337-388. 56. MacVicar BA. Voltage-dependent calcium channels in glial cells. Science 1984; 226:1345-1347. 25 57. Metherate R, Ashe JH. Ionic flux contributions to neocortical slow waves and nucleus basalis-mediated activation: whole-cell recordings in vivo. J Neurosci 1993; 13:5312-5323. 58. Morin D, Steriade M. Development from primary to augmenting responses in the somatosensory system. Brain Res 1981; 205:49-66. 59. Morison RS, Dempsey EW. A study of thalamo-cortical relations. Amer J Physiol 1942; 135:281-292. 60. Nadasdy Z, Csicsvari J, Penttonen M, Buzsaki G. Extracellular recording and analysis of electrical activity: from single cells to ensembles. In: Eichenbaum H, Davis J L, eds. Neuronal ensembles: Strategies for recording and decoding. New York: Wiley-Liss, Pp. 17-55. 61. Nedergaard M. Direct signaling from astrocytes to neurons in cultures of mammalian brain cells. Science 1994; 263:1768-1771. 62. Nedergaard M, Cooper AJ, Goldman SA. Gap junctions are required for the propagation of spreading depression. J Neurobiol 1995; 28:433-44. 63. Nunez PL. Electrical fields of the brain: The neurophysics of EEG. New York: Oxford University Press, 1981. 63. Pedroarena C, Llinas R. Dendritic calcium conductances generate high-frequency oscillation in thalamocortical neurons. Proc Natl Acad Sci (USA) 1997; 94:724-728. 65. Penttonen M, Kamondi A, Sik A, Acsády L, Buzsáki G. Gamma frequency oscillation in the hippocampus: intracellular analysis in vivo. Euro J Neurosci 26 1998; 10:718-728. 66. Petsche H, Pockberger H, Rappelsberger P. On the Search for the Sources of the Electroencephalogram. Neuroscience 1984; 11:1-27. 67. Ralston B, Ajmone-Marsan C. Thalamic control of certain normal and abnormal cortical rhythms. Electroencephal Clin Neurophysiol 1956; 8:559-583. 68. Rappelsberger P, Pockberger H, Petsche H. The contribution of the cortical layers to the generation of the EEG: field potential and current source density analyses in the rabbit's visual cortex. Electroencephal Clin Neurophysiol 1982; 53 :254-269. 69. Ribak CE. Aspinous and sparsely spinous stellate neurons in the visual cortex of rats contain glutamic acid decarboxylase. J Neurocytol 1978; 7:461-478. 70. Ribary U, Ioannides AA, Singh KD, Hasson R, Bolton JPR, Lado F, Mogilner A, Llinás R. Magnetic field tomography (MTF) of coherent thalamo-cortical 40-Hz oscillations in humans. Proc Natl Acad Sci (USA) 1991; 88:11037-11041. 71. Schaul N, Gloor P, Ball G, Gotman J. The electrophysiology of delta waves induced by systemic atropine. Brain Res 1978; 143:475-486. 72. Schwartzkroin PA, Stafstrom CE. Effect of EGTA on the calcium activated afterhyperpolarization in CA3 pyramidal cells. Science 1980; 210:1125-1126. 73. Silva LR, Amitai Y, Connors BW. Intrinsic oscillations of neocortex generated by layer five pyramidal neurons. Science 1991; 251:432-435. 27 74. Somogyi P, Kisvarday ZL, Martin KAC, Whitteridge D. Synaptic connections of morphologically identified and physiologically characterized large basket cells in the striate cortex of the cat. Neuroscience 1983; 10:261-294. 75. Spencer WA, Brookhart JM. Electrical patterns of augmenting and recruiting waves in depth of sensorimotor cortex of cat. J Neurophyiol 1961a; 24:26-49. 76. Spencer WA, Brookhart JM. A study of spontaneous spindle waves in sensorimotor cortex of cat. J Neurophyiol 1961b; 24:50-65. 77. Steriade M, Buzsaki G. Parallel activation of the thalamus and neocortex. In: Steriade M, Biesold D, eds. Brain Cholinergic System. Oxford: Oxford University Press, 1989; (in press). 78. Steriade M, Curro Dossi R, Nunez A. Network modulation of a slow intrinsic oscillation of cat thalamocorticl neurons implicated in sleep delta waves: cortically induced synchronization and brainstem cholinergic suppression. J Neurosci 1991; 11:3200-3217. 79. Steriade M, Deschénes M, Domich L, Mulle C. () Abolition of spindle oscillation in thalamic neurons disconnected from nucleus reticularis thalami. J Neurophysiol 1985; 54(6):1473-1497. 80. Steriade M, McCormick DA, Sejnowski TJ. Thalamocortical oscillations in the sleeping and aroused brain. Science 1993; 262:679-685. 28 81. Steriade M, Gloor P, Llinas RR, Lopes DA Silva FH, Mesulam M-M. Basic mechanisms of cerebral rhythmic activities. Electroencephal Clin Neurophysiol 1990; 76:481-508. 82. Sypert GW, Ward AA. Unidentified neuroglia potentials during propagated seizures in the neocortex. Exp Neurol 1971; 33:239-255. 83. Taylor CP, Dudek FE. Excitation of hippocampal pyramidal cells by an electrical field effect. J Neurophysiol 1984; 52:126-142. 84. Traub RD, Jefferys JGR, Whittington MA. Fast Oscillations in Cortical Circuits. Cambridge, MA: MIT Press, 1999. 85. Traub RD, Dudek FE, Snow RW, Knowles WD. Computer simulations indicate that electrical field effects contribute to the shape of the epileptiform field potential. Neuroscience 1985; 15:947-958. 86. Ts'o D, Frostig R, Lieke E, Grinvald A. Functional Organization of Primate Visual Cortex Revealed by High Resolution Optical Imaging. Science 1990; 249 :333-448. 87. Vanderwolf CH. Cerebral activity and behavior: control by central cholinergic and serotonergic systems. Int Rev Neurobiol 1988; 30:225-340. 88. Walter WG. The contingent negative variation: an electro-cortical sign of sensorimotor reflex association in man. Prog Brain Res 1968; 22:364-377. 89. Wong RK, Prince DA, Basbaum AI. Intradendritic recordings from hippocampal neurons. Proc Natl Acad Sci (USA) 1979; 76:986-990. 29 90. Ylinen A, Bragin A, Nádasdy Z, Jandó G, Szabó I, Sik A, Buzsáki G. Sharp wave associated high frequency oscillation (200 Hz) in the intact hippocampus: network and intracellular mechanisms. J Neurosci 1995; 14:30-46. 91. Yuste, R. and Denk, W. Dendritic spines as a basic unit of synaptic integration. Nature 1995; 375:682-684. 30 Figure legends: Fig. 1. Generation of extracellular field potentials. A. Simultaneously recorded evoked field responses in the CA1-dentate gyrus axis of the rat hippocampus in response to stimulation of the entorhinal input (stim). * Indicate population discharge of the monosynaptically activated granule cells. Their discharge, in turn, activates CA3 pyramidal cells (no shown) whose associational collaterals will depolarize CA1 pyramidal cells and interneurons. This trisynaptic actication of CA1 pyramidal cells is reflected as a late field event (vertical dashed line). B. Spontaneously occurring sharp wave recorded during immobility. The traces are averages of 40 individual events. C. A frontal section of the hippocampus indicating the contact sites of the recording silicon probe (small squares). o, stratum oriens; p, pyramidal layer; r, stratum radiatum; hf, hippocampal fissure; g, granule cell layer; h, hilar region. D and E. Current-source density (CSD) maps of evoked field responses to perforant path stimulatin (D) and of the spontaneous SPW pattern. Sinks (s, inward currents) and sources (so, outward currents) are indicated by cold and warm colors, respectively. iso, zero current flow. The time scale in A and D, B and E are the same. F. Interpretation of the current sinks and sources on the basis of anatomical connectivity. Recorded layers are shown on the right of the pyramidal cell (above) and granule cell (below). Putative active currents are indicated on the right, passive return (re) currents on the left of the pyramidal neuron. Note identical current sinksource distribution of the evoked and spontaneous events in the CA1 region (compare D and E). Sinks in stratum radiatum and oriens reflect excitation of the apical and basal dendrites of CA1 pyramidal cells, respectively by the associational (Schaffer) collaterals of the CA3 region. The large source in the pyramidal layer is a combination of active outward current due to hyperpolarization of the soma by the simultaneously activated basket cells (not shown) and passive return currents from the sinks generated in the basal and apical dendrites. The source in the distal apical dendrites (re) is assumed to represent a passive return current due to the active sink in the middle of stratum radiatum. In addition to EPSPs, dendritic Ca2+ spikes may also contribute to the sinks in strata oriens and radiatum (see Fig. 4). (Reprinted with permission from Buzsaki and Traub, 1996). Fig. 2. Voltage versus depth profiles superimposed onto CSD maps of high voltage spike-and-wave patterns (HVS) and thalamic evoked responses under Ketamine anesthesia and in the awake rat. The 16-site recording probe was located in the 31 somatosensory area. The approximate position of the different layers are indicated left of the CSD maps. Note the overall similarity of the major sinks and sources of the averaged HVS and evoked responses. Major sinks are numbered (1 to 4 in HVS). Vertical dashed lines help identify the earliest sinks/sources. VPLi, primary response; VL and VPLc, augmenting responses 200 msec after the primary response (not shown). In VL, weak early sinks can be indentified in layer VI and layer V, followed by major sinks at similar locations as in the other CSD maps. Stimulating sites are shown in the histological section. The tip of the ipsilateral VPL (VPLi) was two sections (120 µm) posterior to the VL site. An electrolytic lesion was produced at the contralateral VPL (VPLc) stimulating site. Voltage and current calibrations are identical in all panels. iso, baseline isopotential. (Reprinted with permission from Kandel and Buzsaki, 1997). Fig.3. Variations in the voltage-versus-depth profiles and CSD maps of HVSs in the awake rat. A. CSD map of a single HVS episode (3 sec sweep). The superimposed field traces were recorded from layers II, IV and VI, respectively. Note the consistent presence of the layer IV sink but large variability of sinks and sources at other depth locations. B. Selected averages of HVS traces and corresponding CSD maps. All: averages of 200 successive events. ppHVS, average HVS with prominent surface-positive spike component; pHVS, average HVS with less pronounced surface-positive component; nnHVS, average HVS with dominant surface-negative spike component and large sink in layers II-III; nHVS, average HVS with negative spike component in layer IV; DS, average HVS with double spike components at short interspike intervals. Note a prominent delayed sink in layers II-III in ppHVS and nnHVS. Averages from 40 to 50 traces selected from a 5-min recording session. Representative single events of the averages are indicated by vertical lines in A. iso, baseline isopotential. (Reprinted with permission from Kandel and Buzsaki, 1997). Fig. 4. Voltage-dependence of theta-frequency oscillation in a hippocampal pyramidal cell dendrite. Continuous recording of extracellular (extra) and intradendritic (intra) activity in a CA1 pyramidal cell. Holding potential was manually shifted to progressively more depolarized levels by intradendritic current injection (0 to 0.8 nA). The marked epochs (horizontal bars) are shown at faster speed in the bottom records. The recording electrode also contained QX-314 to block Na+ spikes (20 mM). Note large increase of intradendritic theta oscillation amplitude upon depolarization. The relationship of the putative high threshold calcium spikes to the 32 phase of extracellular theta in the CA1 pyramidal layer is indicated by dotted lines. (Reprinted with permission from Kamondi et al., 1998). Fig. 5. HVS-induced fast field oscillation (400-500 Hz ripple). A. Averaged HVS and asssociated unit firing histograms from layers IV to VI. B. Wide-band (a and a'; 1 Hz-5 kHz), filtered field (b and b'; 200-800 Hz) and filtered unit (c and c'; 0.5-5 kHz) traces from layer IV and layer V. C. Averaged fast waves and corresponding unit histograms. The field ripples are filtered (200-800 Hz) derivatives of the wide-band signals recorded from 16 sites. Note sudden phase-reversal of the oscillatory waves (arrow) and phase locked discharges of units in all cortical layers (dashed line). (Reprinted with permission from Kandel and Buzsaki, 1997). 33 Summary Field potential measurements provide an excellent tool for the exploration of network activity in the intact brain. The various rhythms and intermittent EEG potentials can be regarded as time reference points to relate neuronal discharges of single cells. These field potentials (local or global EEG) emerge as a result of synchronous (i. e., simultaneous) changes of the membrane potential of neighboring neurons. Synchronous membrane potential changes can be brought about by synaptic activity (EPSPs and IPSPs), Ca spikes or emerge as a result of intrinsic neuronal patterns (oscillations, burst-induced afterpotentials). The isolated cortical tissue maintains burst discharges of pyramidal cells followed by long-lasting afterhyperpolarization. The synchronous hyperpolarizations in neighboring pyramidal cells can be measured as slow waves in the extracellular space ("synchronization"). In addition, these subcortical neurotransmitters induce a gamma frequency oscillation ("desynchronized" pattern) by activating networks of inhibitory interneurons. 34