Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Caridoid escape reaction wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Aging brain wikipedia , lookup
Single-unit recording wikipedia , lookup
Central pattern generator wikipedia , lookup
Development of the nervous system wikipedia , lookup
Neuroanatomy wikipedia , lookup
Nonsynaptic plasticity wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Activity-dependent plasticity wikipedia , lookup
Biological neuron model wikipedia , lookup
Axon guidance wikipedia , lookup
Optogenetics wikipedia , lookup
Nervous system network models wikipedia , lookup
End-plate potential wikipedia , lookup
Long-term depression wikipedia , lookup
Synaptogenesis wikipedia , lookup
Synaptic gating wikipedia , lookup
Spike-and-wave wikipedia , lookup
Channelrhodopsin wikipedia , lookup
NMDA receptor wikipedia , lookup
Pre-Bötzinger complex wikipedia , lookup
Neurotransmitter wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Signal transduction wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Chemical synapse wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
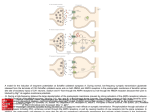
278 Chapter 14 FIGURE 14.4 Transgenic Insertion of a PhotoActivatable Ion Channel that allows the selective stimulation of dopaminergic neurons and their processes. The light-sensitive cation channel, channelrhodopsin-2, tagged with the red fluorescent protein mcherry (ChR2-mcherry) was transgenically expressed from its cDNA in dopaminergic neurons in the mouse brain under the control of the promoter for the dopamine transporter. (A) Dopaminergic neurons in the ventral tegmental area (VTA) of the midbrain expressing the red ChR2-mcherry (left) and an antibody to the dopamine synthetic enzyme tyrosine hydroxylase (TH), counterstained with a green fluorescent secondary antibody (middle). The image on the right shows the left and center pictures merged, and it indicates that ChR2 is restricted to TH-positive neurons. (B) These records were made from a neuron in the nucleus accumbens (which is innervated by dopaminergic fibers from the VTA) in a slice of the midbrain taken from a ChR2-expressing mouse brain. They show short-latency excitatory postsynaptic currents generated when the presynaptic fibers are stimulated by activating the ChR2 with a 5-msec pulse of blue light. The current was inhibited by the glutamate antagonist (DNQX, left side, blue line) but not by the dopamine antagonists SCH23390 + raclopride (SCH/rac, right side). This means that the postsynaptic current did not actually result from the release of dopamine itself, but from the release of glutamate that is stored in the same nerve endings as the dopamine and that activates ionotropic glutamate receptors. (After Stuber et al., 2010). ChR2-mcherry Tyrosine hydroxylase Merge 20 μm (B) 5 ms 5 ms Post DNQX Pre SCH/Rac 20 pA 20 pA Pre DNQX 10 ms Post SCH/Rac 10 ms Visualizing Transmitter-Specific Neurons in Living Brain Tissue 36 Jomphe, C. et al. 2005. J. Neurosci. Methods 146: 1–12. 37 Stuber, G. D. et al. 2010. J. Neurosci. 30: 8229–8233. 38 Gradinaru, V. et al. 2010. Cell 141: 154–165. 39 Kravitz, A. V. et al. 2010. Nature 466: 622–626. 40 Pastrana, E. 2011. Nat. Methods 8: 24–25. 41 Diesseroth, K. 2011. Nat. Methods 8: 26–29. 42 Cooper, J. R., Bloom, F. E., and Roth, R. H. 2003. The Biochemical Basis of Neuropharmacology, 8th ed. Oxford University Press. Certain neurons that contain a transmitter of interest can be color-coded in vivo by using transgenic methods. This can be very helpful for subsequent electrophysiological experiments, especially for neurons that are not very numerous. For example, dopamine-releasing (dopaminergic) neurons in the mesencephalon can be color-coded with the jellyfish green fluorescent protein (GFP, see Box 12.6) by tagging the GFP cDNA onto the cDNA that codes for the promoter region of the synthesizing enzyme tyrosine hydroxylase, then expressing it in mice using transgenic techniques.36 Subsequently, when the mesencephalon is dissected and the cells are isolated and cultured, the dopamine-secreting cells are easily identifiable by their green fluorescence, even though they contribute only a minority of cells in the mixed-cell culture. A similar approach has been used to express a fluorescently tagged, light-activated, cationconducting protein, channelrhodopsin-2, in the dopaminergic neurons37 (Figure 14.4A). Then, dopaminergic neurons in an isolated midbrain slice could be stimulated selectively by a brief flash of blue light, which opened the channelrhodopsin channels to depolarize the neuron (Figure 14.4B). Selectively expressed channelrhodopsins and related proteins are now widely used as a means of stimulating (or inhibiting) neurons,38 particularly for analyzing neural circuits in vivo,39 since light stimulation is easier and more versatile than electrical stimulation of individual neurons through microelectrodes. This field of research has been dubbed “optogenetics.”40,41 Key Transmitters In this section, we highlight some properties and functions of features for a selection of the transmitters previously referred to in Boxes 12.1 and 12.2. Details of chemical structures and metabolic pathways of the small-molecule transmitters are summarized in Appendix B. More detailed information can be obtained in the book by Cooper et al.42 ©2011 Sinauer Associates, Inc. This material cannot be copied, reproduced, manufactured or disseminated in any form without express written permission from the publisher. Neurotransmitters in the Central Nervous System 279 Glutamate This is the fast, excitatory transmitter that is released from neurons throughout the CNS. It activates two types of postsynaptic ionotropic receptors: fast-opening α-amino-3-hydroxy5-methyl-4-isoxazolepropionic acid (AMPA) receptors and slower-opening N-methyl-Daspartate (NMDA) receptors (see Figure 11.12) The AMPA receptors are responsible for normal fast transmission. They are made up of GluA1–4 subunits (see Chapter 5). The speed and duration of the evoked synaptic currents varies substantially between different synapses, depending on the subunit composition of the channel and on the splice-variant (flip or flop43) of the subunit.44 The presence or absence of the GluA2 subunit, and the extent of its mRNA editing45 also affect the calcium permeability of the AMPA receptors.46 One important feature of AMPA receptors is that they are not held in the postsynaptic membrane so tightly as are the nicotinic receptors at the neuromuscular junction, but are highly mobile.47 The lifetime of individual AMPA receptors has been estimated at 10–30 minutes as the receptors recycle between the cell surface and the subsynaptic apparatus.48 Several auxilliary synaptic proteins regulate this trafficking.49 It allows rapid changes in the numbers and subunit composition of synaptic receptors during neural activity, providing the basis for many forms of synaptic plasticity (see Chapter 16). The NMDA receptors are made up of GluN1–3 subunits (see Chapter 5). They differ from AMPA receptors in three main respects: the channels open more slowly; they are blocked by Mg2+ ions and only allow ions to pass when the membrane is depolarized; and they have a much higher permeability to Ca+ ions than most AMPA channels (see Chapter 11). Their voltage-dependence means that they act as coincidence detectors, only allowing current to pass when the neuron is simultaneously depolarized by, for example, highfrequency activation of AMPA channels, ongoing action potential activity, or co-activation of metabotropic glutamate receptors (see below). The consequent entry of Ca2+ ions then induces a variety of downstream effects (see Figure 12.20)—most notably, the induction of long-term potentiation (see Chapter 16). However, there is also a downside, which is that excessive Ca2+ entry through NMDA channels is also neurotoxic (excitotoxic, to give it its original name) and can cause neurodegeneration and ischemic neuron death (stroke).50 Glutamate also activates a distinctive class of G protein–coupled receptors called metabotropic glutamate receptors (mGluRs).51 There are eight of these receptors, mGluR1–8, which are divided into three groups: group I (mGluR1,5), group II (mGluR2,3), and group III (mGluR4,6,7,8) The group I receptors are the main postsynaptic receptors. They couple to the G protein Gq and hence to the phospholipase C–phosphoinositide pathway (see Figure 12.15). Their activation causes a membrane depolarization and increased excitability, mainly by inhibiting Ca2+-activated and M-type, voltage-gated potassium currents. They also release calcium ions from intracellular stores by producing IP3. IP3 synergizes with NMDA receptor activation in inducing long-term potentiation. Thus, mice in which the gene for mGluR1 has been deleted52 show defects in long-term potentiation and learning behavior, while mGluR5 gene deleted mice show deficits in various forms of learning and habituation.53 In contrast, mGluR2, 3, and 4 receptors are more strongly (though not completely) represented in presynaptic glutamatergic fiber terminals. There, they couple to the G protein Go, inhibit the CaV2 Ca2+ channels and so reduce transmitter release. In this way, they mediate a negative feedback control of glutamate release, much like the feedback inhibition of GABA release by presynaptic GABAB receptors (see Figure 11.17). When present postsynaptically, they activate inward rectifier potassium channels, hyperpolarizing the neuron—the opposite effect to that of stimulating mGluR1 or 5 receptors. The mGluR6 receptor hyperpolarizes retinal bipolar cells in a different way: it stimulates cGMP phosphodiesterase, producing a fall in cGMP and closure of cyclic nucleotide-gated cation channels (see Chapter 20). GABA (γ-Aminobutyric acid) and glycine These are the two main inhibitory neurotransmitters in the CNS. They activate chlorideconducting ionotropic GABA or glycine receptors (see Chapter 5) to produce an inhibitory postsynaptic potential (IPSP) in the postsynaptic neuron (see Chapter 11). The increase in chloride conductance tends to inhibit the generation of action potentials by the excitatory postsynaptic potentials (EPSPs). GABA is the predominant inhibitory transmitter 43 Hollman, M., and Heinemann, S. 1994. Annu. Rev. Neurosci.17: 31–108. 44 Geiger, J. R. et al. 1997. Neuron 18: 1009–1023. 45 Seeburg, P. H., and Hartner, J. 2003. Curr. Opin. Neurobiol. 13: 279–283. 46 Cull-Candy, S., Kelly, L. and Farrant, M. 2007. Curr. Opin. Neurobiol. 17: 277–280. 47 Tardin, C. et al. 2003. EMBO J. 22: 4656– 4665. 48 Choquet, D., and Trille, A. 2003. Nat. Rev. Neurosci. 4: 251–265. 49 Nicoll, R. A., Tomita, S., and Bredt, D. S. 2006. Science 311: 1253–1256. 50 Papardia, S., and Hardingham, G. E. 2007. Neuroscientist 13: 572–579. 51 Niswender, C., and Conn, P. J. 2010. Ann. Rev. Pharmacol. Toxicol. 50: 295–322. 52 Aiba, A. et al. 1994. Cell 79: 365–375. 53 Bird, M. K., and Lawrence, A. J. 2009. Trends Pharmacol. Sci. 30: 617–623. ©2011 Sinauer Associates, Inc. This material cannot be copied, reproduced, manufactured or disseminated in any form without express written permission from the publisher. 280 Chapter 14 54 Gold, M. R., and Martin, A. R. 1984. Nature 308: 639–641. 55 Jonas, P., Bischofberger, J., and Sandkühler, J. 1998. Science 281: 419–424. 56 Cobb, S. R. et al. 1995. Nature 378: 75–78. 57 Whittington, M. A., and Traub, R. D. 2003. Trends Neurosci. 26: 676–682. 58 Klausberger, T., and Somogyi, P. 2008. Science 321: 53–57. 59 Buszaki, G. 1984. Prog. Neurobiol. 22: 131–153. 60 Pouille, F., and Scanziani, M. 2001. Science 293: 1159–1163. 61 Betz, H., and Laube, B. 2006. J. Neurochem. 97: 1600–1610. 62 Sivilotti, L. 2010. J. Physiol. 588: 45–58. 63 Olsen, R. W., and Sieghart, W. 2008. Pharmacol. Rev. 60: 243–260. 64 Möhler, H. 2006. Cell Tissue Res. 326: 505–516. 65 Brown, N. et al. 2002. Brit. J. Pharmacol. 136: 965–974. 66 Farrant, M., and Nusser, Z. 2005. Nat. Rev. Neurosci. 6: 215–229. 67 Franks, N. P. 2008. Nat. Rev. Neurosci. 9: 370–386. 68 Nicoll, R. A. et al. 1975. Nature 258: 625–627. 69 Scholfield, C. N. 1978. J. Physiol. 275: 559–566. 70 Scholfield, C. N. 1980. Pflügers Arch. 383: 249–255. 71 Steinbach, J. H., and Akk, G. 2001. J. Physiol. 537: 715−733. 72 Bianchi, M. T. et al. 2009. Epilepsy Res. 85: 212–220. 73 Wallner, M., Hanchar, H. J., and Olsen, R. W. 2003. Proc. Natl. Acad. Sci. USA 100: 15218–15223. 74 Kumar, S. et al. 2009. Psychopharmacology (Berl.) 205: 529–564. 75 Hosie, A. M. et al. 2006. Nature 444: 486–489. 76 Mitchell, E. A. et al. 2008. Neurochem Int. 52: 588–595. 77 Smart, T. G., Hosie, A. M., and Miller, P. 2004. Neuroscientist 10: 432–442. 78 Bowery, N. G. et al. 1980. Nature 283: 92–94. released by neurons in the cortex and midbrain, whereas glycine plays a prominent role in the brainstem and spinal cord. Some neurons have receptors for both transmitters, which act on separate and distinguishable chloride channels,54 and both transmitters can sometimes be released from the same neuron.55 GABA and glycine are concentrated in interneurons. Their primary function is to control the output of the principal excitatory neurons by negative feedback through recurrent inhibitory collaterals from the axons of these neurons. This prevents excessive excitatory discharges from the principal neurons. One example (already mentioned) is the negative feedback inhibition of motoneuron discharges by recurrent axons that innervate glycinergic interneurons (Renshaw cells) in the spinal cord.3 The importance of inhibitory interneurons in the brain is illustrated by the fact that blocking the inhibitory GABA or glycine receptors (with bicuculline or strychnine, respectively) produces convulsions. However, inhibitory interneurons in the cortex also have more subtle functions. Because each interneuron innervates several principal neurons, they serve to coordinate the output of the principal neurons and to synchronize activity within networks of neurons.56–58 In addition to receiving recurrent collaterals from principal neuron axons, these GABAergic interneurons also receive a direct input from collaterals of the afferent fibers to principal neurons, producing what is termed feed-forward inhibition.59 This helps to set the timing of principal neuron’s response to afferent stimuli.60 RECEPTORS FOR GABA AND GLYCINE Ionotropic GABAA and glycine receptors are pentameric (five subunit) receptors homologous with nicotinic ACh receptors but with the difference that their structure favors permeation of the anion chloride instead of cations (see Chapter 5). The glycine receptor is made up of two α-subunits and three β-subunits, arranged as a ring in the order α-β-β-α-β, with four binding sites for glycine at the α-β interfaces.61 Three genetic variants of the α-subunit are known (α1 to α3), but only one form of the β-subunits (β1). Sivilotti eloquently discusses the mechanism whereby glycine opens these channels, as deduced from single channel analysis.62 The GABA receptor has a more complex structure and repertoire of subunits. There are 19 different subunits: 6 α-subunits, 3 β, and 3 γ, one each of δ, ε, π, θ, and three variants of a special ρ-subunit.63 The most common combinations at inhibitory synapses are α1β2γ2 (60%), α2β3γ2 (15%–20%) and α3βnγ2 (10%–15%).64 Because different combinations differ in rates of activation, deactivation and desensitization, their expression is tuned to the optimum requirements for inhibition at different synapses.64 GABA receptors containing the δ-subunit are particularly interesting because these do not desensitize and are activated by unusually low (sub-micromolar) concentrations of GABA.65 Delta subunits, along with α6-subunits, are present in receptors on cerebellar and hippocampal dentate gyrus granule cells. They are tonically activated by resting interstitial GABA concentrations (estimated at 300–600 nM) and so produce a steady component of resting membrane current.66 The ρ-subunits, assembled as a homomeric pentamer, make up another unique type of GABAA receptor present in the retina. The GABAA receptor is also the prime target for a number of important drugs that act allosterically to enhance GABA-mediated currents and IPSPs. These include benzodiazepine compounds, barbiturates, and certain anesthetics such as propafol, etomidate and the steroid anesthetic alphaxolone.64,67 These three groups of drugs bind at different sites on the receptor and have different actions. For example, barbiturates68,69 and alphaxolone70 prolong the postsynaptic current, by slowing the closure of the channels,71 whereas benzodiazepines increase the sensitivity of the channels to GABA (by slowing dissociation) without changing open time.72 Enhancement of GABAA receptor currents is also responsible for some of the effects of ethanol,73,74 which is why one should not mix alcohol with benzodiazepines. Naturally occurring regulators of GABAA receptor activity in the brain include endogenous neurosteroids75,76 and zinc ions.77 GABA B RECEPTORS Like glutamate, GABA can also activate a G protein–coupled receptor. This is termed the GABAB receptor. It was discovered by Norman Bowery and his colleagues in 1980.78 They found that GABA reduced the release of transmitters from peripheral and central neurons by activating a receptor that was not antagonized by GABAA receptor antagonists but could be selectively activated by the GABA analog, baclofen (β-chlorophenyl-GABA). They subsequently identified it as a G protein–coupled ©2011 Sinauer Associates, Inc. This material cannot be copied, reproduced, manufactured or disseminated in any form without express written permission from the publisher. Neurotransmitters in the Central Nervous System 281 Extracellular GABAB1 NH2 GABA Su2 Su1 γ β LIV-BP Presynaptic Ca2+ γ AC α α Postsynaptic K+ γ β α γ β β HOOC 1 – Cytoplasm NH2 GABAB2 COOH 2 + FIGURE 14.5 Schematic Diagram of the GABAB Receptor Dimer. The functional receptor is a dimer composed of GABAB(1) and GABAB(2) monomers joined through a coiled–coil domain at their C-termini. GABA binds to a part of the extracellular domain of the B(1) monomer at a region homologous to the bacterial periplasmic leucine–isoleucine–valine binding protein, or LIV-BP. The B(2) monomer 3 – responds by activating the G proteins Gi and Go. The α-subunit of Gi inhibits adenylate cyclase (pathway 1), while the βγ-subunit activates the postsynaptic G protein gated inward rectifier channel (pathway 2). The βγ-subunit of Go also inhibits the CaV2 class of calcium channels in the presynaptic terminals (pathway 3). (After Couve et al., 2000.) receptor because (among other properties) its action was suppressed by Pertussis toxin. In fact, it turned out to be a very unusual receptor because the active receptor is a dimer of two closely similar receptors termed GABAB(1) and GABAB(2), conjoined through their intracellular C-termini79,80 (Figure 14.5). The B1-subunit has the binding site for GABA whereas the B2-subunit is necessary to activate the G protein. Hence, both subunits have to be expressed and assembled to make a functional receptor.81 GABAB receptors are located both postsynaptically and presynaptically, and have different effects at the two sites.82 Activation of postsynaptic receptors opens inwardly rectifying G protein–gated potassium channels83 (Figure 14.6), which produces a membrane hyperpolarization and delayed postsynaptic inhibition. 82 These postsynaptic GABAB receptors are located around the periphery of the synapse (perisynaptically) and do not readily respond to GABA released by a single afferent stimulus; instead, they are activated by spillover GABA escaping from the synapse during repetitive stimulation or following simultaneous activation of several GABAergic inputs.84 Presynaptic GABAB receptors inhibit voltage-gated calcium channels in the nerve terminal and thereby reduce transmitter release. When GABA is released, it may act on receptors on its own terminal (autoinhibition; see Figure 11.17) or on adjacent terminals that release GABA or another transmitter (i.e., heterosynaptic inhibition). Inhibition of the presynaptic calcium current and its consequent effect on transmitter release is illustrated in the experiment on calyx of Held terminals shown in Figure 14.7A. These terminals, located in the medial nucleus of the trapezoid body, are large enough to be patch clamped.85 When the presynaptic terminal was depolarized by injecting current through the patch electrode, application of baclofen reduced both the resulting inward calcium current and the EPSP (Figure 14.7B). The reduced EPSP could be explained solely by the reduced calcium current since the relationship between inward current and EPSP amplitude was unaltered (Figure 14.7C), However, at some other synapses there may be an additional effect of the GABAB-released G protein βγ-subunits on the transmitter release mechanism itself.86,87 Acetylcholine Cholinergic neurons are present throughout the brainstem, midbrain, and telencephalon and provide a widespread innervation to diverse areas of the brain.88 An important source of cholinergic axons to the cerebral cortex and hippocampus is provided by groups of neu- 79 Couve, A., Moss, S. J., and Pangalos, M. N. 2000. Mol. Cell. Neurosci. 16: 296–312. Bettler, B. et al. 2004. Physiol. Rev. 84: 835–867. 81 Filippov, A. K. et al. 2000. J. Neurosci. 20: 2867–2874. 82 Nicoll, R. A. 2004. Biochem. Pharmacol. 68: 1667–1674. 83 Gahwiler, B. H., and Brown, D. A. 1985. Proc. Natl. Acad. Sci. USA 82: 1558–1562. 84 Scanziani, M. 2000. Neuron 25: 673–681. 85 Takahashi, T., Kajikawa, Y., and Tsujimoto, T. 1998. J. Neurosci. 18: 3138–3146. 86 Wu, L. G., and Saggau, P. 1997. Trends Neurosci. 20: 204–212. 87 Blackmer, T. et al. 2005. Nat. Neurosci. 8: 421–425. 88 Woolf, N. J. 1991. Prog. Neurobiol. 37: 475–524. 80 ©2011 Sinauer Associates, Inc. This material cannot be copied, reproduced, manufactured or disseminated in any form without express written permission from the publisher.