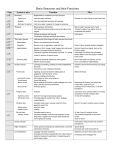
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Affective neuroscience wikipedia , lookup
Multielectrode array wikipedia , lookup
Sensory cue wikipedia , lookup
Neural coding wikipedia , lookup
Human brain wikipedia , lookup
Executive functions wikipedia , lookup
Response priming wikipedia , lookup
Binding problem wikipedia , lookup
Environmental enrichment wikipedia , lookup
History of neuroimaging wikipedia , lookup
Functional magnetic resonance imaging wikipedia , lookup
Aging brain wikipedia , lookup
Neural modeling fields wikipedia , lookup
Neuroesthetics wikipedia , lookup
Psychophysics wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Neuroeconomics wikipedia , lookup
Perception of infrasound wikipedia , lookup
Neuroplasticity wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Optogenetics wikipedia , lookup
Cortical cooling wikipedia , lookup
Synaptic gating wikipedia , lookup
C1 and P1 (neuroscience) wikipedia , lookup
Metastability in the brain wikipedia , lookup
Time perception wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Cognitive neuroscience of music wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Inferior temporal gyrus wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Evoked potential wikipedia , lookup
Spontaneous and Stimulus-Evoked Intrinsic Optical Signals in Primary Auditory Cortex of the Cat MATTHEW W. SPITZER, MICHAEL B. CALFORD, JANINE C. CLAREY, JOHN D. PETTIGREW, AND ANNA W. ROE Vision, Touch and Hearing Research Centre, Department of Physiology and Pharmacology, The University of Queensland, St. Lucia, Queensland 4072, Australia Received 14 June 2000; accepted in final form 3 November 2000 Spitzer, Matthew W., Michael B. Calford, Janine C. Clarey, John D. Pettigrew, and Anna W. Roe. Spontaneous and stimulus-evoked intrinsic optical signals in primary auditory cortex of the cat. J Neurophysiol 85: 1283–1298, 2001. Spontaneous and tone-evoked changes in light reflectance were recorded from primary auditory cortex (A1) of anesthetized cats (barbiturate induction, ketamine maintenance). Spontaneous 0.1-Hz oscillations of reflectance of 540and 690-nm light were recorded in quiet. Stimulation with tone pips evoked localized reflectance decreases at 540 nm in 3/10 cats. The distribution of patches “activated” by tones of different frequencies reflected the known tonotopic organization of auditory cortex. Stimulus-evoked reflectance changes at 690 nm were observed in 9/10 cats but lacked stimulus-dependent topography. In two experiments, stimulus-evoked optical signals at 540 nm were compared with multiunit responses to the same stimuli recorded at multiple sites. A significant correlation (P ⬍ 0.05) between magnitude of reflectance decrease and multiunit response strength was evident in only one of five stimulus conditions in each experiment. There was no significant correlation when data were pooled across all stimulus conditions in either experiment. In one experiment, the spatial distribution of activated patches, evident in records of spontaneous activity at 540 nm, was similar to that of patches activated by tonal stimuli. These results suggest that local cerebral blood volume changes reflect the gross tonotopic organization of A1 but are not restricted to the sites of spiking neurons. Aside from the systematic representation of frequency (Merzenich and Brugge 1973; Reale and Imig 1980), details of the functional organization of auditory cortex remain controversial. Several studies have provided evidence of compartmental organization within the tonotopic representation of primary auditory cortex (A1) in the cat based on clustered or banded distributions of neurons with different response properties (e.g., Imig and Adrian 1977; Mendelson et al. 1993; Middlebrooks and Pettigrew 1981; Phillips et al. 1994; Schreiner and Mendelson 1990). Due to limitations of conventional microelectrode mapping techniques, however, a comprehensive view of the functional organization of A1 has yet to emerge. In a single mapping experiment, it is only possible to characterize effects of a few stimulus parameters from a large number of recording sites. Consequently, it is not yet clear how the various topographies of response properties described by dif- ferent studies relate to one another. A second limiting factor is the lack of anatomical markers. The discovery of histological markers for functionally distinct neuronal subpopulations, most notably cytochrome oxidase, was a considerable aid to elucidating the functional organization of visual cortex (Horton and Hubel 1981; Humphrey and Hendrikson 1983; WongRiley 1979). In auditory cortex, by contrast, the same markers provide little compelling evidence for compartmentalization (Tootell et al. 1985; Wallace et al. 1991). Resolution of this issue in auditory cortex may have implications for our understanding of cortical organization in general. Optical imaging (OI) of intrinsic signals (Grinvald et al. 1986; Ts’o et al. 1990) offers both vastly increased spatial sampling density and the potential to sample cortical responses to a larger stimulus repertoire in a single experiment than is possible with microelectrode mapping, making it a promising alternative approach for studying the physiological organization of A1. This technique uses changes in light reflectance, associated with physiological activity, to visualize the location of cortical tissue activated by a given stimulus. The light reflectance signals derive from multiple sources, including changes in local blood volume, which dominate the signal at wavelengths below 600 nm, and changes in the oxygenation state of hemoglobin and light scattering, which provide a major source of signals at longer wavelengths (Frostig et al. 1990; Malonek and Grinvald 1996; Malonek et al. 1997). Because neuronal responses are measured indirectly, it is essential to verify results obtained by this method with electrophysiological recording. Although most studies of visual cortex have shown a high degree of correspondence between stimulusevoked optical and neurophysiological responses (Crair et al. 1997; Ghose and Ts’o 1997; Grinvald et al. 1986; Roe and Ts’o 1995; Shmuel and Grinvald 1996; Shoham et al. 1997), under certain conditions, the areal extent of over which evoked optical signals are recorded can be several times greater than that of the neuronal spiking response (Das and Gilbert 1995). Recently, several groups have used OI of intrinsic signals to study the topography of stimulus evoked activity in auditory cortex of rodents (Bakin et al. 1996; Harrison et al. 1998; Hess and Scheich 1996). Results of these studies differ from those of single-unit mapping studies of cat A1 (Heil et al. 1994; Phillips Present address and address for reprint requests: M. W. Spitzer, Institute of Neuroscience, University of Oregon, Eugene, OR 97403 (E-mail: spitzer@ uoneuro.uoregon.edu). The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. INTRODUCTION www.jn.physiology.org 0022-3077/01 $5.00 Copyright © 2001 The American Physiological Society 1283 1284 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE et al. 1994) in several respects, including threshold and time course of evoked activity, spatial distribution of tone-evoked activity, and effects of changing sound pressure level (SPL). It is not clear, however, to what extent these apparent differences are due to species differences or recording methodology because in only one case (Bakin et al. 1996) were optical images verified with electrophysiological recording. For a number of reasons, including the large exposed surface area of auditory cortex and the elaborate auditory cortical specializations, most mapping studies of A1 have been performed on the domestic cat (Reale and Imig 1980). As a result, it is possible to formulate relatively high-order questions about physiological organization in this species. On the other hand, all published intrinsic signal-based OI studies of A1 have been conducted on rodents, about which much less is known. The present study was therefore undertaken to characterize stimulus-evoked intrinsic optical signals at different wavelengths in cat A1 and to compare optical responses to neuronal spiking responses elicited by the same stimuli. Preliminary accounts of the results were published in abstract form (Spitzer and Clarey 1998). METHODS Anesthesia and surgical procedures All surgical and experimental procedures were approved by the Animal Experimentation Ethics Committee of the University of Queensland and conformed to the guidelines for use of animals of the National Health and Medical Research Council of Australia. Experiments were performed on cats between 8 wk and 10 mo of age. Stimulus-evoked optical signals at 540 nm, presented in the following text, were obtained from three cats, aged 18, 20, and 27 wk. Young animals were used because it has been reported that the signal-tonoise ratio of optical signals is higher than in older animals (Hubener et al. 1997). Moreover, because response properties of auditory cortical neurons such as latency and threshold mature to adult-like levels within the first three to four postnatal weeks (Brugge et al. 1988), it is unlikely that the present results are compromised by any physical immaturity of auditory cortex. Animals were initially sedated with ketamine hydrochloride and inspected to ensure there were no signs of external ear blockage or middle ear infection. Surgical anesthesia was then induced with a single dose of pentobarbital sodium (25 mg/kg ip). Atropine sulfate (10 mg/kg sc) was administered to minimize bronchial mucous secretion. Anesthesia was maintained with supplemental doses of ketamine hydrochloride (0.5–5 mg 䡠 kg⫺1 䡠 h⫺1 iv, typically starting 2 h after barbiturate injection) throughout the subsequent surgery and recording session. The subject’s heart rate was monitored throughout the recording session. During the optical and electrophysiological recording sessions, anesthetic and supplementary fluids (lactated Ringer solution) were administered by intravenous injection and/or continuous intravenous infusion; anesthetic dose rate was adjusted to maintain a constant heart rate and no signs of skeletal muscle activity (indicated by high-frequency contamination of the electrocardiogram). Anesthetic level was sufficient to attenuate the eye-blink reflex. Throughout all procedures, the animal’s temperature was maintained with a homeothermic heating blanket (Harvard Instruments). Following induction of surgical anesthesia, the trachea was cannulated and an intravenous line was established. A craniotomy was performed to expose the temporal cortical surface with the dura intact. A recording chamber was mounted over the skull opening with acrylic cement. The dura was then reflected to expose the cortical surface, and the recording chamber was filled with low-viscosity silicone oil (Corning) and sealed with a glass cover. Following the optical recording session, the top of the recording chamber was removed to permit introduction of microelectrodes. Dexamethosone was administered (1 mg/kg iv) prior to reflection of the dura and opening of the recording chamber to prevent cerebral edema. During the recording sessions, subjects were situated on a vibration isolation table (Newport Instruments) housed in an anechoic chamber. The cat’s head was stabilized by attachment of the skull to a stereotaxic frame. Following experiments, animals were euthanized by overdose of pentobarbitone sodium (120 mg/kg iv). Optical data acquisition and acoustic stimulation Intrinsic optical signals were recorded using the Imager 2001 video acquisition system (Optical Imaging, Germantown, NY), the details of which have been described previously (Bonhoeffer and Grinvald 1996; Grinvald et al. 1986, 1988; Ts’o et al. 1990). The cortical surface was illuminated with narrowband light, generated by band-pass filtering (band centers, 540, 630, or 690 ⫾ 10 nm; interference filters from Edmund Scientific, Barrington, NJ) the output of a tungsten light source, and conveyed by two fiber-optic light guides. Video images of the illuminated cortex were acquired at 30 frames per second with a CCD camera (Bischke, Japan). The camera was positioned orthogonal to the exposed surface of putative primary auditory cortex (gyral surface between the tops of the anterior ectosylvian and posterior ectosylvian sulci). Data were collected with the camera focused between 350 and 500 m below the cortical surface to minimize artifacts due to the presence of surface blood vessels. A typical imaging session began at least 4 h after induction of anesthesia and continued for 12 h. While 12-bit cameras permit the encoding of absolute reflectance value, the Imager 2001 system employs differential imaging methodology to allow the use of standard 8-bit video technology. In this method, a reference image (average of 64 video frames) is obtained prior to each block of recording trials. This reference image is subtracted from each subsequent recorded image within a block of trials. The reference-subtracted images constitute the stored data files on which further analyses are performed. Such initial reference subtraction allows the encoding of the differential signal within the 8-bit limit and has comparable signal-to-noise resolution as that obtained by storing the absolute reflectance values. Absolute reflectance values can be reconstructed from the reference images. In this study, intrinsic optical signals were acquired by summing video frames captured over 0.5 s for stimulus-evoked signals, or 1 s for spontaneous signals, followed by reference subtraction. The resulting differential image was then amplified, digitized (8 bits), and stored on computer disk. The stored image produced by this process will be referred to hereafter as a data frame. Acoustic stimuli were generated digitally (MALab, Kaiser Instruments, Irvine, CA) and presented via a calibrated piezo-electric transducer (Motorola KSN 1141) positioned 25 cm from the ear contralateral to the imaged cortex and approximately 50° in azimuth from the cat’s midline. Individual acoustic stimuli consisted of trains of five to seven identical tone pips (10-ms cosine rise and fall, 70-ms duration, 730-ms inter-pip interval), as illustrated in Fig. 5. Detailed stimulus parameters are included in figure legends. Stimuli were presented in sets consisting of five or six combinations of frequency and sound pressure level (SPL) and a no-stimulus condition, presented in random order. Acoustically evoked optical signals were recorded by acquiring a sequence of data frames during presentation of each acoustic stimulus. Acquisition of optical data was synchronized to acoustic stimulus presentation, starting one frame (500 ms) prior to stimulus onset and continuing for up to 6.5 s after stimulus offset (see Figs. 3 and 5). Following this data acquisition period, there was a silent interval of 12 s. To remove periodic spontaneous variation of the OPTICAL IMAGING IN AUDITORY CORTEX optical signal (see RESULTS), it was necessary to sum optical data recorded during multiple repeated trials of each stimulus set. Typically, data were collected and stored on-line in blocks of four or five repeated stimulus sets. Acoustically evoked and spontaneous optical signals were visualized by off-line analysis (SumAnal program) (Ghose and Ts’o 1997). For acoustically evoked images, corresponding data frames (i.e., same stimulus and time after stimulation) from multiple blocks of stored data were first summed. Each successive summed frame collected after stimulus onset was then divided by the frame collected prior to stimulus onset within the trial. (n.b. This differential analysis procedure should not be confused with the initial reference image subtraction performed prior to saving data, on-line.) The resulting values indicate the change in light reflectance (⌬R) at each measured pixel following stimulus onset. Further analyses are described in RESULTS. To display ⌬R data as images, images were clipped (top and bottom 19% of the pixel value distribution removed) to eliminate large signals due to blood vessel artifact and fit to a common linear 33 color scale (shown in each figure). Spontaneous optical signals were visualized using a similar procedure of frame-by-frame, temporal analysis of signal development. However, data from individual trials (20-s data acquisition period, no stimulus) were analyzed separately without summation across blocks. The present data were obtained in 10 experiments performed on cats maintained under ketamine anesthesia. An earlier series of experiments was performed on 15 cats maintained under barbiturate anesthesia (pentothal sodium). In general, results obtained using barbiturate anesthesia were inconclusive due to excessive variability. However, a few references are made to results from these experiments, where appropriate. Electrophysiological recording Electrophysiological mapping was performed in four of the experiments. Following the imaging session, a dose of dexamethosone (1 mg/kg iv) was administered, and the lid of the recording chamber was removed. A reference image of the cortical surface collected under illumination with green light (590 nm) was used to guide electrode placement and for later alignment of optical and electrophysiological data. Glass-coated, tungsten microelectrodes (0.5–2.5 M⍀ impedance at 1 kHz) were introduced perpendicular to the cortical surface using a stepping motor microdrive (Herb Adams, Caltech) controlled from outside the anechoic chamber. The electrode signal was amplified (⫻1,000 gain) and filtered 0.3–5 kHz. Recordings of multi- and single-unit activity were obtained by level triggering (MALab). Multiunit recordings were only obtained at sites where action potentials of the constituent units could be clearly distinguished visually using an oscilloscope. Single-unit recordings were also distinguished by visual criteria. In each electrode penetration, an attempt was made to record from the closest site to the cortical surface exhibiting acoustically driven unit activity. In practice, the majority of recordings were obtained at depths between 400 and 800 m. Neuronal responses to the stimulus set used for optical imaging were collected at each recording site. In addition, an attempt was always made to determine the frequency/level combination that gave an optimal response. The response to the “optimal” stimulus was also recorded if it was considerably greater than responses to any of the stimuli used for optical imaging. Slight modifications to the stimulation protocol were made for electrophysiological recording: stimuli were presented as 70-ms-duration tone pips (10-ms cosine rise/fall), 730-ms interstimulus interval, 20 repetitions. The time of occurrence of triggered action potentials and stimulus events were logged (more than 1-s resolution) and stored in a FIFO buffer, from which they were retrieved by the host computer and stored on disk. 1285 RESULTS Spontaneous optical signals Spontaneous changes of light reflectance in auditory cortex were observed in the absence of acoustic stimulation in 9/10 cats. In most cases, the spontaneous signals were manifest as irregular variations of reflectance in the no-stimulus condition but were obscured by averaging signals from multiple trials. To visualize the spontaneous signal directly, optical signals were recorded without averaging from one cat. An example of the spontaneous optical signal recorded under illumination with 540-nm light is shown in Figs. 1 and 2. The change in cortical light reflectance (⌬R) during a 20-s sampling period was measured by recording 21 consecutive 1-s data frames and dividing frame 2 through 21 by frame 1. During the sampling period, the reflectance of 540-nm light increased and decreased relative to the initial frame in an oscillatory manner at a frequency of 0.1 Hz (Fig. 1). The optical signal thus recorded has two components. The first component is a coherent oscillation of reflectance at all points on the cortical surface (Fig. 1C). A second component consists of systematic variation of the amplitude of signals recorded at different sites. The local variations of signal amplitude can be dissociated from the global variations by dividing the reflectance change at each site by the mean reflectance change measured over the entire image. Such analysis (Fig. 1D) reveals that the amplitude of reflectance change at each site oscillates relative to the image mean at a frequency of 0.1 Hz and that the phase of oscillation shifts with changes of position across the cortical surface. The local variation of optical signals can also be visualized by displaying the reflectance change at each measured pixel as an image (individual frames in Fig. 2; see METHODS for details). In this case, the global reflectance changes are removed by fitting the histogram of pixel values of each frame to a common color map, a similar transformation to division by the image mean. When viewed as an image, it becomes apparent that in each frame areas of similar relative reflectance change form distinct patches, several millimeters across. A similar 0.1-Hz oscillation of cortical light reflectance was also recorded using 690-nm light (not shown). Stimulus-evoked optical signals Stimulus-evoked intrinsic optical signals were clearly evident in only 3/10 cats imaged with 540-nm light. A similar success rate has been reported for imaging at 540 nm in chinchilla auditory cortex (Harrison et al. 1998). In two of the unsuccessful cases, normal multiunit responses were recorded after the imaging sessions, indicating that our failure to record optical signals was not a result of a physiologically unresponsive auditory cortex. In successful cases, it was necessary to average responses to large numbers of stimulus presentations to obtain acceptable image quality and to negate the spontaneous signals. An example of the optical signal recorded at 540 nm by averaging responses to 50 presentations of a tone pip train is illustrated in Figs. 3 and 4. Stimulus-evoked optical signals were measured by recording optical data for 10.5 s, starting just prior to stimulation, and dividing data frames collected after stimulus onset by the frame collected before stimulus onset (Fig. 3C). Within 3 s of stimulus onset, reflectance begins to decrease at nearly all points on the cortical 1286 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE FIG. 1. Oscillatory spontaneous optical signals recorded with 540-nm light. A: schematic illustrates the location of the imaged area with respect to the major cortical sulci. AES, anterior ectosylvian sulcus; PES, posterior ectosylvian sulcus; SSS, suprasylvian sulcus; M, medial; A, anterior. B: macroscopic view of the exposed cortical surface. Scale bar ⫽ 5 mm. C: reflectance change was measured in a 162 ⫻ 129 pixel region (box in A and B) by recording video data during a 21-s period as a series of 21 data frames and dividing pixel values in frames 1–20 by the corresponding pixel values in frame 0. Mean reflectance change (⌬R) measured in 5 ⫻ 5 pixel regions (crosses in B) underwent coherent oscillatory change throughout the 20-s sampling period. D: dividing ⌬R values measured at each site by the mean ⌬R value of all pixels of each frame (mean ⌬R) revealed incoherent local oscillations of ⌬R amplitude. surface (Fig. 3D). Reflectance continues to decrease throughout the duration of the 4.8-s stimulus sequence and then returns to prestimulation levels following stimulus offset. The magnitude of the stimulus evoked reflectance changes at most sites is much greater than that of any fluctuations evident in the no stimulus condition (- - -). The absence of large reflectance changes in the no-stimulus condition indicates that signal averaging was sufficient to negate the contribution of spontaneous signals. Local variations of the amplitude of stimulusevoked reflectance changes are clearly evident in Fig. 3D. Division by the image mean reveals a restricted region exhibiting relative decrease of reflectance near the center of presumptive A1 (Fig. 3E). The image representation (Fig. 4A) shows development of an “activated” area (red and orange), corresponding to the location of maximum reflectance decrease, and, presumably, maximum blood volume increase, within 1 s of stimulus onset. The activated area remains relatively stable from 2-s postonset until stimulus offset and then quickly dissipates. Note, also, the late “activation” of posteromedial and anterolateral regions that showed relative increases of reflectance during stimulation. In comparison to the stimulated condition, the unstimulated condition shows only minor and unsystematic fluctuations of reflectance (Fig. 4B). The position of the activated region revealed by optical signals at 540 nm is dependent on stimulus frequency. Optical signals recorded from the same cortex in response to six different frequency-level combinations are shown in Fig. 5. Responses to stimuli presented at 65 dB SPL show a clear progressive shift of the activated region from caudal to rostral as stimulus frequency is incremented from 5 to 20 kHz, in agreement with the known tonotopic organization of cat A1 (Reale and Imig 1980). In contrast to the pattern of neuronal activation, described previously (Heil et al. 1994; Phillips et al. 1994; Schreiner et al. 1992), the position of optical activation evoked by 15-kHz tones remains constant across a 30-dB range of stimulus level. The decaying phase of the activation sequence follows a curious course in responses to the 15-kHz, 80-dB and 20-kHz, 65-dB stimuli. In both cases, following stimulus offset the region of decreased reflectance shifts in a caudal direction. Similar shifts of the activated region after stimulus offset or during the later stages of the stimulation period were observed in all three successful 540-nm imaging experiments. In this particular case, as a result of the shift, the images obtained for over 2 s following offset of the 20-kHz stimulus are very similar to the stimulus-evoked images recorded in response to the 5-kHz stimulus. Thus in this and the other two cases, a topographic relationship of reflectance changes was apparent only for the period during stimulus presentation. Weak stimulus-evoked intrinsic optical signals were also evident in 9/10 cats imaged using 690-nm light and ketamine anesthesia, including those exhibiting stimulus-evoked signals at 540 nm. The magnitude of evoked reflectance changes at 690 nm was approximately one-tenth that observed at 540 nm. In contrast to the optical signals recorded at 540 nm, at 690 nm, reflectance changes were uniformly distributed across the cortical surface, and their topography was uninfluenced by stimulus frequency. Multiunit mapping was performed in four cats OPTICAL IMAGING IN AUDITORY CORTEX 1287 FIG. 2. Temporal progression of spontaneous optical signals recorded at 540 nm from the boxed region in Fig. 1. Numbered images correspond to data frames averaged over successive 1-s sampling periods. Crosses indicate positions sampled in Fig. 1. Regions showing minimum ⌬R values, corresponding to maximum reflectance decrease and, presumably, maximum increase in local blood volume, are shown in red, and maximum ⌬R values are shown in black. ⌬R values were mapped, linearly, onto the indicated color table. ⌬R values less than or greater than the clipped range are shown in red and black, respectively. Scale bar ⫽ 2 mm. exhibiting stimulus evoked signals at 690 nm, and in all cases, revealed normal multiunit responses with stimulus-dependent topography. Cross-condition analyses of the evoked optical signals (e.g., dividing responses to different frequencies and levels, subtracting responses to a single condition from the sum of all conditions, dividing across binaural conditions in barbiturate anesthetized animals), such as those performed in studies of visual cortex (Bonhoeffer and Grinvald 1993; Frostig et al. 1990; Ts’o et al. 1990), did not reveal any stimulus-dependent topography at 690 nm. Finally, similarly uniform and stimulusfeature-independent topography of intrinsic optical signals was observed in barbiturate anesthetized cats using light band-pass filtered at 690 or at 636 nm or long-pass filtered with 650-nm cutoff. Comparison of optical and multiunit responses Multiunit mapping was performed in 2/3 animals that exhibited stimulus-evoked optical signals at 540 nm. Results of one experiment in which optical signals and multiunit electrophysiological data were recorded from the same cortex are shown in Fig. 6. The optical images form a tonotopic sequence with the region of activation shifting progressively from caudal to rostral as stimulus frequency is incremented from 5 to 20 kHz. Multiunit responses were classified as either clear responses, no response, or near-threshold responses (details in legend). In each image, there are a number of multiunit recording sites at which the optical and electrophysiological responses are in apparent agreement (e.g., 5 kHz: 1, 6, 9; 10 kHz: 3, 5, 8, 9; 15 kHz: 4, 5, 6; 20 kHz: 1, 3, 4, 5, 6, 8) and a number at which they clearly disagree (e.g., all stimuli at sites 10 and 11; 10 kHz: 1, 2, 4; 15 kHz: 3; 20 kHz: 7). Multiunit records and optical signals recorded at each site in response to the 20 kHz stimulus are shown in Fig. 7 to permit detailed comparisons. This case was chosen for illustration because it represents the best agreement between optical and multiunit responses. While there appears to be good agreement between optical and multiunit responses at the majority of sites (e.g., 1– 6, and 8), there are also sites where the two measures clearly disagree (10 and 11), and others where the relationship is questionable (7 and 9). Examination of cases where the optical and multiunit re- 1288 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE FIG. 3. Stimulus-evoked optical signals recorded at 540 nm. A: schematic illustrates location of imaged region. Abbreviations and conventions same as Fig. 1. B: macroscopic view of the exposed cortex. Scale bar ⫽ 5 mm. C: stimulus consisted of a train of 7 tone pips (pip duration, 70 ms; rise/fall time, 10 ms; interpip interval, 730 ms) of 10 kHz at 65 dB SPL. Data frames were summed from 50 trials per stimulus. After summation, frames 1–20 were individually divided by frame 0 to calculate ⌬R. D: ⌬R decreased at all sites during the period of stimulus presentation (䡲 above time axis) and returned to prestimulus levels thereafter. Stimulus evoked optical signals (—) were much larger than reflectance changes measured in the unstimulated condition (- - -). E: division by the image mean revealed systematic local variations of amplitude of stimulus evoked signals. sponses disagreed revealed several examples of both false positive (decrease of reflectance relative to image mean, but no multiunit response; e.g., Fig. 6, site 3 at 15 kHz, sites 1 and 4 at 10 kHz) and false negative (clear multiunit response but no reflectance decrease relative to image mean; e.g., Figs. 6 and 7, sites 10 and 11 for all stimuli) optical signals. In experiment 98s11, 55 comparisons of optical and multiunit responses (responses to 5 stimulus conditions compared at 11 sites) revealed 22 clear mismatches of which 9 were false positives and 13 false negatives. Cases where the multiunit response was classified in the threshold category were not counted in this analysis. In experiment 98s5, 95 comparisons (5 stimulus conditions, 19 sites) revealed 30 mismatches of which 8 were false positives and 22 were false negatives. Thus in both experiments in which multiunit data are available, the majority of errors were false negatives. It is not obvious from the preceding qualitative comparisons whether the level of agreement of optical signals and multiunit responses is higher than that expected by chance. To address this issue, quantitatively, the magnitude of optical and multiunit responses was compared at each electrophysiological recording site in two cats. A strong association would be expected to result in a significant negative correlation between these two response measures. Scatter plots relating the magnitudes of optical signals and multiunit responses, recorded at the same sites, in response to all stimuli that elicited clear, stimulus-evoked optical signals, are shown in Fig. 8 (see legend for details). To enable comparison of multiunit responses across recording sites, response magnitude at each site is expressed as a percentage of the response to the optimal stimulus at that site. Sites at which an optimal stimulus could not be determined were excluded from analysis. The level of correlation between magnitudes of optical signals and multiunit responses was assessed separately for each stimulus condition and also for all responses pooled across stimuli within each experiment, by calculating Spearman’s rank order correlation coefficient (Rs; a nonparametric measure was used because the multiunit response data were not normally distributed due to a strong ceiling effect). Correlation values for individual stimulus conditions are shown in Table 1. In only 3 of the 10 cases was a significant (P ⬍ 0.05) correlation detected between optical and multiunit response magnitudes. Furthermore, in one of the three cases showing significant correlation, the direction of relationship was reversed. Due to the small sample sizes, it remains possible that a weak association existed in the remaining cases but was not detected. However, analysis of data pooled across conditions in each experiment (Fig. 8) provided no evidence of a significant correlation. Thus the present data provide little evidence of an association between the magnitude of stimulus-evoked optical and multiunit responses. Relation of stimulus-evoked and spontaneous optical signal Finally, an intriguing similarity between spatial patterns of spontaneous and stimulus-evoked optical signals was observed OPTICAL IMAGING IN AUDITORY CORTEX 1289 FIG. 4. Optical images illustrate the relatively focal topography of stimulus-evoked optical signals recorded at 540 nm. Crosses indicate locations sampled in Fig. 3. Scale bar ⫽ 3 mm. A: in the stimulated condition, a focal region of maximum reflectance decrease develops and is maintained for the duration of stimulus presentation. ⌬R values for each image frame were mapped onto the color table shown such that numerically smallest values, corresponding to maximum reflectance decrease, are red and numerically largest values are black. B: only minor reflectance changes are evident in the unstimulated condition. in the single case for which both forms of data are available. A comparison of spontaneous and stimulus-evoked images obtained at 540 nm from the same cortex, is shown in Fig. 9. The top row of images are single frames (frame 5, 2,000- to 2,500-ms poststimulus onset) taken from sequences of stimulus-evoked activity, which form a tonotopic sequence as shown in Fig. 6. The images in the middle row are single frames, arbitrarily selected, from sequences of spontaneous optical activity recorded from the same cortex. The apparent similarity of patterns of stimulus-evoked and spontaneous optical signals was confirmed by subtracting each image in the middle row from the image above it (results shown on bottom row). The resulting difference images show little overall variation from the mean value, with most of the remaining peaks and troughs corresponding to major blood vessels. The similarity of topography of stimulus-evoked and spontaneous images might appear to suggest that all of the images resulted from spontaneous activity. This suggestion is, however, untenable because it 1290 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE FIG. 5. Optical images recorded at 540 nm show tonotopic organization of stimulus-evoked reflectance changes. Images were collected in response to 6 frequency ⫻ sound pressure level combinations (left) and a no-stimulus condition from the same cortex as in Figs. 3 and 4. The region of peak activation during the stimulation period shifts from posterior to anterior as stimulus frequency is incremented from 5 to 20 kHz. Schematic, at bottom, illustrates timing of stimulus sequence relative to image frames. cannot account for synchronization of reflectance changes to onset of acoustic stimulation, persistence of stimulus-evoked signals despite averaging images from multiple presentations, dependence of the topography of evoked images on stimulus frequency, nor for absence of major reflectance changes in the no-stimulus condition after extensive averaging. An alternative, albeit speculative explanation is that the neuronal or vascular functional units that are activated by acoustic stimulation are also spontaneously active in the absence of stimulation. DISCUSSION The major findings of the present study are that both spontaneously occurring and stimulus-evoked optical signals were observed in auditory cortex of anesthetized cats; the spatial distribution of stimulus-evoked optical signals recorded at 540 nm was consistent with the known tonotopic organization of A1; and despite the stimulus-dependent topography of optical signals, the distribution of maximum reflectance changes was apparently not related to the distribution of extracellularly recorded neuronal activity. The mismatch between the spatial distribution of optical signals and multiunit responses suggests that the optical signal at 540 nm corresponds to local blood volume changes within vascular functional units, recruited in response to physiological activation, whose fine structure is imprecisely matched to that of the multiunit response. The following sections discuss the rationale for these conclusions as well as alternative explanations of our results. Differences in recording depth of optical and multiunit responses Due to constraints imposed by light penetration and scattering, intrinsic optical signals are recorded from the superficial cortical layers (Bonhoeffer and Grinvald 1996). In these experiments, optical signals were recorded at a depth of 500 m below the cortical surface. Because the superficial cortical layers contained few electrically responsive units, multiunit responses were recorded at depths ranging from 320 to 1,200 m, with mean recording depths of 821 and 580 m for experiments 98s5 and 98s11, respectively. The difference in depths at which optical and multiunit responses were recorded raises two issues pertinent to interpretation of the present results and may help to explain the low success rate of the imaging experiments. First, do neuronal response properties vary with depth in A1? In agreement with the general concept of columnar organization, several studies have shown that response properties, including type of response to contralateral ear stimulation and frequency tuning, remain consistent throughout the depth of A1 (Abeles and Goldstein 1970; Imig OPTICAL IMAGING IN AUDITORY CORTEX 1291 FIG. 6. Comparison of multiunit responses and optical images recorded at 540 nm in 1 animal. Multiunit responses were recorded at numbered sites and classified qualitatively as either clear responses, threshold responses, or no response. The threshold category included responses of less than 7 spikes per 20 stimulus presentations and in a few cases where it was difficult to differentiate a putative response from spontaneous activity. Optical images of responses to tone pips at 65 dB SPL were collected as shown in Fig. 4, using identical stimulus parameters. Image frames 4 –7 were summed, and results were median filtered to reduce noise. and Adrian 1977), including the superficial layers (Reser et al. 2000). A major exception is that, under binaural conditions, the influence of ipsilateral ear stimulation may vary with cortical depth (Phillips and Irvine 1979; Reser et al. 2000). Despite the variation of ipsilateral ear influences, however, Clarey et al. (1994) found that preferred azimuth remained homogenous throughout cortical depth in 72% of electrode penetrations. Thus although it is possible that the response to the contralaterally located speaker, used in the present study, would change with depth at some recording sites, such effects would be limited to a minority of sites. Furthermore it is difficult to evaluate the extent to which such depth effects might confound the comparison of multiunit and optical responses because it is unknown whether the optical signals are generated in response to subthreshold neuronal activity in the superficial cortical layers or suprathreshold activity in the middle layers. The depth disparity is only relevant in the latter case. A second issue is whether the 540-nm optical signals could be displaced laterally from their site of origin due to blood flow with an orientation parallel to the cortical surface. This would appear unlikely because the arterioles and venules that supply the cortical parenchyma are oriented orthogonal to the cortical surface (Edvinsson et al. 1993). Reflecting this anatomical arrangement, numerous studies have demonstrated that local blood flow within the cerebral cortex follows a course parallel to that of neuronal columns (e.g., Bryan and Duckrow 1995). Thus optical signals with a hemodynamic basis should remain aligned with their column of origin regardless of recording depth. Finally, the paucity of neuronal responses in the superficial layers of A1, where optical signals are recorded, is an obvious potential cause of the low success rate of the imaging experiments. Future attempts at intrinsic signal based imaging of A1 might benefit from use of anesthetics, such as isofluorane, which are less disruptive to cortical activity patterns, or an awake preparation. Spontaneous optical signals In the present study we observed spontaneous oscillatory changes of reflectance of both red and green light at frequencies close to 0.1 Hz. Spontaneous optical signals with similar properties have been reported previously (Mayhew et al. 1996). Using similar optical recording methods, Mayhew and coworkers observed 0.1-Hz oscillations of reflected dichromatic light (570 and 660 nm) from the surfaces of diverse brain structures in anesthetized rats and unanesthetized cats. Furthermore they demonstrated that the change of intensity of reflected light was highly correlated with changes of local blood flow measured in the same tissue by laser-Doppler flowmetry. Thus it is highly likely that the spontaneous optical signal recorded by Mayhew et al., and in the present study results from vasomotion, a 0.1-Hz oscillation of local cerebral and somatic blood flow that has been widely reported (Dirnagl et al. 1989; Fagrell et al. 1980; Golanov et al. 1994; Meyer et al. 1988; Morita-Tsuzuki et al. 1992). Spontaneous optical signals are a major source of noise that must be overcome to record stimulus-evoked signals. The spontaneous signals measured by Mayhew et al. could be as large as 1–2% of the total reflected light signal; whereas, the “mapping signals” used to define functional architecture by intrinsic signal based optical imaging are typically an order of magnitude smaller (Frostig et al. 1990; Grinvald et al. 1988). At least one other optical imaging study of auditory cortex has reported “spontaneous waves of cortical activity” with a cycle of 5–10 s that acted as a major source of interference (Harrison et al. 1998). In the present 1292 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE FIG. 7. Comparison of optical (540 nm) and multiunit responses to 20 kHz, 65 dB stimulus recorded at numbered sites in Fig. 6. Optical signals recorded in a 5 ⫻ 5 pixel region centered on each multiunit site in the stimulated and unstimulated conditions are shown as — and - - -, respectively. Multiunit responses are shown as poststimulus time histograms and dot rasters below each optical signal. Ordinate axes of histograms are scaled relative to the maximum response recorded at each site. Pip duration ( ) is shown on the horizontal axis of each histogram. study, averaging data from multiple stimulus presentations (more than 16) proved sufficient to remove major contributions of the spontaneous signals. Comparison with previous studies of auditory cortex Results of recent optical imaging studies of auditory cortex in ketamine-anesthetized chinchillas (Harrison et al. 1998) and barbiturate-anesthetized cats (Dinse et al. 1996) are similar to those obtained with 540-nm light in the present study. Harrison et al. (1998) recorded tone-evoked optical signals at 540 nm with similar time course and topography to those reported here. Threshold levels for reliably eliciting optical signals were between 40 and 50 dB SPL. Both previous imaging studies reported monotonic growth of activated regions as stimulus SPL was increased and tonotopic organization of activated regions at suprathreshold SPL. Results obtained with optical imaging at 540 nm differ in several respects from the previously described maps of singleunit (Phillips et al. 1994), and multiunit (Heil et al. 1994; OPTICAL IMAGING IN AUDITORY CORTEX 1293 FIG. 8. Peak amplitude of optical signals recorded at 540 nm is plotted against multiunit response magnitude at each recording site for all stimuli that evoked clear optical responses in experiments 98s11 and 98s5. Stimuli are listed in Table 1. Only stimuli that evoked a clear decrease in reflectance (⌬R) at multiple recording sites, initiated within the first 3 s after stimulus onset, were included in this analysis. Spearman’s rank order correlation coefficient (Rs) are shown atop each plot. In neither case was the correlation significant (P ⬍ 0.05). Optical signals were measured by calculating the reflectance change (⌬R) in a 5 ⫻ 5 pixel window centered at each multiunit recording site and determining the maximum absolute value occurring within 3 s of stimulus onset. Multiunit response magnitude is expressed as percentage of response to the optimum stimulus at each recording site. Stimulus parameters and optical recording procedures are identical to those in Fig. 4. Schreiner et al. 1992), responses to tones in A1. First, singleunit thresholds were often lower than 10 dB SPL, and patches of neighboring sites containing units responding to a given frequency were evident at 20 dB SPL, at least 20 dB below threshold for optical signals. Second, as SPL was increased, the location of patches of active neurons changed such that the location of neurons responding maximally at the highest SPL was usually very different from the location of neurons active near threshold SPL. All maps contained at least some patches of recording sites where neuronal activity decreased as a function of increasing SPL. By contrast, the area of activated cortex indicated by optical recording grows monotonically with increasing SPL, remaining centered on the site showing activation at threshold (Dinse et al. 1996; Harrison et al. 1998). Finally, at high SPL (e.g., at least 60 dB), the distribution of single-unit activity consisted of multiple, discrete, patches of active neurons separated by intervening areas of unresponsive neurons. The largest continuous patches of active neurons were not more than 3 mm across. Optical imaging, on the other hand, typically reveals a single, large region of activation (Dinse et al. 1996; Harrison et al. 1998; present results). In our hands the activated region could span more than 5 mm (Figs. 4 and 5). Finally, the lack of correspondence between multiunit responses and optical signals at 540 nm reported here is unsurprising, given the major discrepancies between the previously reported optical and single-unit maps. It might be argued that the maps of unit activity obtained using contralateral ear stimulation, are not directly comparable to the images obtained in the present study, using free-field stimulation. Certainly, the single-unit maps obtained with different procedures are likely to differ, primarily as a result of differences in the contribution of ipsilateral ear stimulation. However, the effects of SPL on the distribution of active 1. Correlations of ⌬R and multiunit response magnitude for individual stimuli TABLE Experiment n 98s11 11 98s5 19 Stimulus 5 10 10 15 20 5 10 10 10 15 kHz, kHz, kHz, kHz, kHz, kHz, kHz, kHz, kHz, kHz, 65 50 65 65 65 65 50 65 80 65 dB dB dB dB dB dB dB dB dB dB Rs Significance 0.03 ⫺0.43 ⫺0.29 ⫺0.13 ⫺0.63 0.58 ⫺0.35 ⫺0.32 ⫺0.52 0.19 NS NS NS NS P ⬍ 0.05 P ⬍ 0.01 NS NS P ⬍ 0.05 NS neurons primarily reflect the patchy organization of neurons with different forms of sensitivity to SPL (Clarey et al. 1994; Heil et al. 1994; Phillips et al. 1994; Sutter and Schreiner 1995), as opposed to binaural interaction, and are therefore unlikely to be dramatically altered by the difference in stimulation procedures. Two studies, thus far, have reported tone-evoked optical signals at wavelengths greater than 600 nm in auditory cortex. Hess and Sheich (1996) recorded at 610 nm in auditory cortex of unanesthetized gerbils, and Bakin et al. (1994) recorded at 630 nm in barbiturate-anesthetized rats and guinea pigs. In agreement with the present findings, in both studies optical imaging indicated activation of very large, continuous regions of A1. The contiguous and broad distribution of activated regions revealed by optical imaging at 630 nm in rats (Bakin et al. 1996) differs from the patchy distribution of single-unit activity reported by Phillips et al. (1994) in cats. Subsequent multiunit mapping, however, revealed much better agreement between optical and neuronal responses in rats than was evident in the present study. The mapping data presented by Bakin et al. (1994) permit 214 comparisons of optical and multiunit responses (2 stimulus conditions at 107 sites) of which only 33 showed clear disagreement between the two measures. Thus the difference between imaging results in rats and single-unit mapping results in cats may indicate a fundamental difference in the topographic representation of suprathreshold tones in auditory cortex of the two species. In the gerbil, the location of the activated region revealed by optical imaging shifted at later stages of the stimulation period and continued to shift after cessation of stimulation (Hess and Scheich 1996). Similar shifts were observed in all three cats successfully imaged at 540 nm in the present study. Hess and Sheich suggested that shift of the activated area is indicative of a corresponding shift of subthreshold electrophysiological activity. However, in the absence of corroborating electrophysiological data, and in light of the lack of correspondence between optical signals and neuronal responses demonstrated here, it is equally likely that the phenomenon reflects the spatiotemporal properties of the hemodynamic response rather than those of the neuronal response. Comparison with previous studies of visual and somatosensory cortex The findings of the present study differ from those of intrinsic signal-based optical imaging studies of visual and somatosensory cortices in two regards. First, in the present study 1294 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE FIG. 9. Similarity of stimulus-evoked and spontaneous optical images recorded at 540 nm. Top: optical images in response to stimuli of 5 kHz (A), 10 kHz (B), and 20 kHz (C) at 65 dB SPL. Data are the same as in Fig. 6. Middle: individual 1-s frames selected from 3 20-s epochs of spontaneous activity recorded as in Fig. 1. Bottom: results of subtracting images in the middle row from images in the top row (G ⫽ A–D; H ⫽ B–E; I ⫽ C–F). optical signals with stimulus-dependent topography were recorded at 540 nm but not at longer wavelengths. By contrast, in visual cortex, similar functional maps of ocular dominance and orientation preference can be obtained at wavelengths ranging from 480 to 940 nm (Frostig et al. 1990). In rodent somatosensory cortex, stimulation of a single facial vibrissa also evokes optical images with similar spatial location and extent at 550, 610, and 850 nm (Narayan et al. 1994). The wavelength dependence of optical signals in cat auditory cortex suggests that stimulus-evoked changes in local blood volume, oxygen saturation of hemoglobin, and light scattering, which provide the major sources of optical signals recorded at 540, 630, and 690 nm, respectively (Frostig et al. 1990; Malonek and Grinvald 1996; Malonek et al. 1997), are not as well coupled in this cortex as they are other sensory cortices and, presumably, in auditory cortex in other species (Bakin et al. 1996; Harrison et al. 1998; Hess and Scheich 1996). The reason for this apparent difference is not readily apparent. A second difference is that, in the present study, the topographies of optical signals and neuronal spiking responses evoked by the same stimulus were unrelated. By contrast, several studies have demonstrated a tight correspondence between the spatial distribution of optical and spiking responses in visual cortex (Crair et al. 1997; Ghose and Ts’o 1997; Grinvald et al. 1986; Roe and Ts’o 1995; Shmuel and Grinvald 1996; Shoham et al. 1997) and rodent somatosensory cortex (Masino et al. 1993; Peterson et al. 1998). In the following text, we consider three possible explanations for the latter difference. Visualization of subthreshold and interneuron activity Two studies of visual cortex, using spatially restricted stimuli, provide evidence that intrinsic optical signals reflect both sub- and suprathreshold neuronal activity. In contrast to previous imaging studies that used full-field visual stimulation, these studies demonstrated that partial field stimulation evokes optical signals distributed over a much larger cortical area than the corresponding neuronal spiking responses. Using stimuli OPTICAL IMAGING IN AUDITORY CORTEX designed to simulate the receptive field size of a single cortical neuron, Das and Gilbert (1995) reported that the areal extent of evoked optical signals was approximately 20 times greater than that of evoked spiking activity. In a second study, using partial field stimulation, single-unit data provided evidence of subthreshold physiological activity within regions containing evoked optical signals but no spiking neurons (Toth et al. 1996). Thus in visual cortex, stimuli that elicit spatially restricted patterns of spiking activity, as do tones in auditory cortex, reveal large discrepancies between the spatial distribution of optical and unit responses, most likely due to visualization of subthreshold physiological activity. It is therefore possible that visualization of subthreshold activity could be a primary cause of the mismatch between patterns of optical activation and spiking activity observed in the present study. Another potential source of the mismatch between optical and unit activity is the difficulty of recording spiking activity from inhibitory interneurons with extracellular electrodes. Mountcastle et al. (1969) reported that the majority of extracellular recordings obtained with metal electrodes in somatosensory cortex exhibited a type of waveform referred to as “regular spikes.” Only occasionally were recordings obtained from neurons with “thin spikes,” characterized by a comparatively shorter initial negative phase, smaller amplitude, and spatially restricted electrical field. A slightly higher proportion of “thin,” or “fast spiking,” neurons have been recorded in studies using glass micropipette electrodes (e.g., Simons 1978). More recently, intracellular experiments have confirmed Mountcastle’s proposal that regular spiking neurons have pyramidal morphology and thin spiking neurons have aspiny stellate morphology, typical of GABAergic inhibitory interneurons (McCormick et al. 1985). These findings provide strong circumstantial evidence that metal extracellular electrodes record preferentially from pyramidal neurons. The temporal properties of stimulus-evoked spike bursts recorded extracellularly in auditory cortex are consistent with this view (Bowman et al. 1995; Phillips et al. 1996). Consequently a mismatch between multiunit and optical responses could potentially arise in areas where spiking activity of interneurons generates a strong optical signal, while simultaneously inhibiting nearby pyramidal neurons. There is certainly considerable evidence that stimulation with tones at high SPL elicits subthreshold electrophysiological activity throughout a substantial proportion of auditory cortex, including a major component due to inhibitory processes and, presumably, spiking activity in inhibitory interneurons. First, because many A1 neurons have frequency-domain inhibitory side bands (Calford and Semple 1995; Calford et al. 1993; Phillips 1988; Phillips and Cynader 1985; Schreiner and Mendelson 1990; Shamma and Symmes 1985; Shamma et al. 1993), a tonal stimulus of a given frequency will induce widespread inhibition of neurons with higher and lower best frequencies. Second, many A1 neurons exhibit level-dependent suppression (Phillips and Irvine 1981), referred to as “nonmonotonicity,” a substantial proportion of which is likely generated by intracortical inhibitory mechanisms (Barone et al. 1996; Prieto et al. 1994). Because nonmonotonic neurons are distributed in patches (Clarey et al. 1994; Phillips et al. 1994), the spatial pattern of cortical activity induced by tones at high SPL should be expected to include patches of spiking inhibitory interneurons, together with inhibited pyramdial neurons. 1295 Consistent with this contention, a recent voltage-sensitive dyebased imaging study of guinea pig auditory cortex demonstrated that tonal stimulation produces regions of activation, flanked by regions of inhibition, and that the area of excitatory activation could be dramatically expanded by local application of GABA antagonists (Horikawa et al. 1996). Finally, although binaural inhibitory mechanisms would not be expected to have a major influence on the distribution of unit responses to contralaterally presented tonal stimuli, such stimuli might induce subthreshold activation of “predominantly binaural” cortical neurons (Kitzes et al. 1980). In addition to the difference between optical and multiunit responses within A1, failure to distinguish between supra- and subthreshold activity might also account for the difference in the degree of correspondence of optical and unit responses between A1 and visual cortex. In visual cortex, columns of neurons with similar orientation preference are interconnected by a lattice-like array of axonal projections (Gilbert and Wiesel 1983, 1989; Rockland and Lund 1983). As a result, stimulation of a single orientation column gives rise to subthreshold excitatory and inhibitory synaptic inputs preferentially directed to surrounding columns with similar orientation preference (Grinvald et al. 1994; Weliky et al. 1995). Consequently, the spatial pattern of subthreshold electrophysiological activation induced by partial visual field stimulation corresponds very closely to the pattern of suprathreshold activation induced by full-field stimulation. Thus for full-field stimulation, the failure to distinguish between sub- and suprathreshold activation has little effect on the correspondence of optical and unit responses. In auditory cortex, by contrast, it is likely that the patches of activated and actively inhibited pyramidal neurons are spatially separated. Thus visualization of subthreshold physiological activity could have a far greater effect on results in auditory cortex than in visual cortex. The hypothesis that the discrepancy between optical and unit responses in cat auditory cortex results entirely from visualization of subthreshold activity, and/or inability to record spiking activity of interneurons, is inconsistent, however, with the fact that the majority of mismatches that we observed were false-negative errors. In either case, one expects the majority of errors to be false positives. Although our data do not allow us to rule out a contribution of visualized subthreshold activity or interneuronal spiking activity, it is unlikely that either is the major cause of discrepancies between optical and multiunit responses observed here. Timing of auditory cortical neuronal responses The characteristic temporal response properties of neurons in auditory cortex of anesthetized cats might account for the difference in results of optical imaging in auditory and other cortical areas. Most neurons in A1 respond to best-frequency tonal stimulation with one or two spikes synchronized to stimulus onset, followed by suppression of spiking activity for the duration of the stimulus (Fig. 7). In addition, because repeated stimulation at short interstimulus intervals results in decreased responsiveness, it is necessary to use a stimulation duty cycle with a low ratio (⬃1:10) of stimulus-on time to stimulus-off time to obtain maximum responsiveness to individual stimuli (Hocherman and Gilat 1981; Phillips et al. 1989). As a consequence, using optimal stimulus parameters, suprathreshold 1296 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE neuronal responses are only elicited during a very small fraction of the duration of a repetitive tone-pip sequence. By contrast, sustained neuronal responses lasting several seconds are elicited by optimal stimuli in visual cortex of barbiturate anesthetized animals (Hubel and Wiesel 1959). Because of this difference in response properties, discharge rates of optimally stimulated neurons in visual cortex, recorded throughout a 5-s stimulus presentation, may exceed those of auditory cortical neurons by more than a factor of 15 (Roe and Ts’o 1995; Sengpiel and Blakemore 1994). In barrel cortex, periodic vibrissal stimulation elicits sustained spike discharges, with two spike bursts per stimulus period (Peterson et al. 1998). The 4- to 5-Hz stimulation frequencies, typically used in optical imaging studies, would thus elicit discharge rates several times those typically observed in auditory cortex. The gross differences in amount of suprathreshold neuronal responses throughout the period of stimulation in different cortical areas may be reflected in the corresponding optical signals. Specifically, the paucity of neuronal spiking responses may be insufficient to generate a reliable and robust optical signal at 540 nm in auditory cortex. This explanation is consistent with the unreliability of the optical signal, which was only detectable in 3/10 cats tested, and then only after extensive averaging. This hypothesis may, at first, appear inconsistent with the fact that optical imaging has been shown to reveal both suprathreshold and subthreshold activity (Das and Gilbert 1995; Toth et al. 1996), as the paucity of spike discharges in A1 is at least partly due to an abundance of stimulus-evoked inhibitory processes some of which may last beyond the stimulus duration (Volkov and Galazjuk 1991, 1992). It should be noted, however, that optical signals accompanying suprathreshold activity are larger than those accompanying subthreshold activity and can be distinguished on this basis using appropriate analysis (Toth et al. 1996). Furthermore it is not clear how inhibitory activity is reflected by intrinsic optical signals. Thus it remains entirely plausible that the failure of optical signals recorded at 540 nm to reveal physiologically relevant functional topographies in cat auditory cortex is due in large part to the sparseness of neuronal spiking responses. If this is the case, one potential approach to improving the quality of optical signals recorded from auditory cortex would be the use of an awake, behaving preparation. Previous studies have indicated that tones evoke a wider range of response types in A1 of unanesthetized animals, including a greater prevalence of sustained responses (Abeles and Goldstein 1972; Brugge and Merzenich 1973; Pfingst et al. 1977). One previous optical imaging study of auditory cortex used an awake preparation (Hess and Scheich 1996), but in the absence of corroborating physiological data, there is no reason to suppose that the results were any different to those of the present study. Vascular organization Finally, we consider the possibility that the success of optical imaging in visual and barrel cortex depends on specializations of the vascular supply that may be absent in cat auditory cortex. The primary and secondary visual cortices of primates are distinguished by a finely structured modular organization that is demarcated by the pattern of cytochrome oxidase staining (Horton and Hubel 1981; Livingstone and Hubel 1984a; Murphy et al. 1995; Wong-Riley 1979). Neurons within the different cytochrome oxidase defined modules have different response properties (Hubel and Livingstone 1987; Livingstone and Hubel 1984a; Silverman et al. 1989; Ts’o and Gilbert 1988). Furthermore the modularity of functional organization is reflected in the distribution of metabolic activity (Humphrey and Hendrikson 1983; Tootell et al. 1983) as well as the patterns of intrinsic and interareal cortico-cortical connections (Hubel and Livingstone 1987; Levitt et al. 1994; Livingstone and Hubel 1984b, 1987; Rockland and Lund 1983; Yoshioka et al. 1996), and in the distribution of a number of neurochemical markers including CAT-301 (Hendry et al. 1984) and parvalbumin (Blumcke et al. 1990). At least in macaque monkeys, the modular neuronal organization within visual cortex is also reflected by the distribution of small diameter blood vessels (Zheng et al. 1991). The density of capillaries and radial penetrating vessels is greater in cytochrome oxidase-rich blobs and stripes than in the intervening cortical tissue. Similarly, in barrel cortex each histologically defined barrel is delimited by a dense capillary plexus within the barrel core (Patel 1983). There is not, however, a one-to-one relationship between individual barrels and vascular supply, as individual penetrating arterioles give rise to capillaries in several adjacent barrels (Woolsey et al. 1996). Nevertheless, in both visual and barrel cortex, there is evidence that the microvasculature is organized into functional units that bear a close spatial relationship to the anatomically and physiologically defined neuronal functional modules. Because optical imaging relies primarily on blood-borne signals, it is possible that the organization of vascular elements sets constraints on the spatial pattern of activation that can be visualized with this technique. Consistent with this view, the areal extent of activation in barrel cortex produced by nearthreshold deflection of a single vibrissa (Peterson et al. 1998) corresponds very well with the areal extent of the minimal vascular functional unit, consisting of the capillaries supplied by a single penetrating arteriole (Woolsey et al. 1996). If an equivalent complementarity of neuronal and vascular functional units does not exist in auditory cortex, the local blood volume changes, presumably the major source of optical signals recorded at 540 nm, might be spatially decoupled from the pattern of electrophysiological activity. It is difficult to further evaluate this conjecture at present, however, because it is not known whether A1 exhibits modular neuronal organization, comparable to that in visual cortex, and because the microscopic organization of blood supply to A1 has not been described. We thank S. Kaiser, M. Ward, and the staff of Optical Imaging, without whose technical expertise these experiments would not have been possible. We also thank D. Vaney for contributing the vibration isolation table, D. Ts’O for use of the SumAnal program, and K. Keller for help with image analysis. This work was supported by National Health and Medical Research Council of Australia Project Grants 971147 and 96/NHMRC9002G to M. W. Spitzer and J. D. Pettigrew, respectively, and by NHMRC Project Fellowship Grant 961226 to M. B. Calford. Present addresses: M. B. Calford, Faculty of Medicine and Health Sciences, The University of Newcastle, Callaghan, NSW 2308, Australia; J. C. Clarey, Dept. of Otolaryngology, The University of Melbourne, Melbourne, VIC 3010, Australia; A. W. Roe, Dept. of Neurobiology, Yale University School of Medicine, New Haven, CT 06520. OPTICAL IMAGING IN AUDITORY CORTEX REFERENCES ABELES M AND GOLDSTEIN MH JR. Functional architecture in cat primary auditory cortex: columnar organization and organization according to depth. J Neurophysiol 33: 172–187, 1970. ABELES MA AND GOLDSTEIN MH. Responses of single units in the primary auditory cortex of the cat to tones and to tone pairs. Brain Res 42: 337–352, 1972. BAKIN JS, KWON MC, MASINO SA, WEINBERGER NM, AND FROSTIG RD. Suprathreshold auditory cortex activation visualized by intrinsic signal optical imaging. Cereb Cortex 6: 120 –130, 1996. BARONE P, CLAREY JC, IRONS WA, AND IMIG TJ. Cortical synthesis of azimuthsensitive single-unit responses with nonmonotonic level tuning: a thalamocortical comparison in the cat. J Neurophysiol 75: 1206 –1220, 1996. BLUMCKE I, HOF PR, MORRISON JH, AND CELIO MR. Distribution of parvalbumin immunoreactivity in the visual cortex of Old World monkeys and humans. J Comp Neurol 301: 417– 432, 1990. BONHOEFFER T AND GRINVALD A. The layout of iso-orientation domains in area 18 of cat visual cortex: optical imaging reveals a pinwheel-like organization. J Neurosci 13: 4157– 4180, 1993. BONHOEFFER T AND GRINVALD A. Optical imaging based on intrinsic signals: the methodology. In: Brain Mapping: The Methods, edited by Toga AW and Mazziotta JC. San Diego, CA: Academic, 1996, p. 55–97. BOWMAN DM, EGGERMONT JJ, AND SMITH GM. Effect of stimulation on burst firing in cat primary auditory cortex. J Neurophysiol 74: 1841–1855, 1995. BRUGGE JF AND MERZENICH MM. Responses of neurons in auditory cortex of the macaque monkey to monaural and binaural stimulation. J Neurophysiol 36: 1138 –1158, 1973. BRUGGE JF, REALE RA, AND WILSON GF. Sensitivity of auditory cortical neurons of kittens to monaural and binaural high frequency sound. Hear Res 34: 127–140, 1988. BRYAN RM JR AND DUCKROW RB. Radial columns in autoradiographs generated from tracer methods for measuring cerebral cortical blood flow. Am J Physiol Heart Circ Physiol 269: H583–H589, 1995. CALFORD MB, RAJAN R, AND IRVINE DR. Rapid changes in the frequency tuning of neurons in cat auditory cortex resulting from pure-tone-induced temporary threshold shift. Neuroscience 55: 953–964, 1993. CALFORD MB AND SEMPLE MN. Monaural inhibition in cat auditory cortex. J Neurophysiol 73: 1876 –1891, 1995. CLAREY JC, BARONE P, AND IMIG TJ. Functional organization of sound direction and sound pressure level in primary auditory cortex of the cat. J Neurophysiol 72: 2383–2405, 1994. CRAIR MC, RUTHAZER ES, GILLESPIE DC, AND STRYKER MP. Relationship between the ocular dominance and orientation maps in visual cortex of monocularly deprived cats. Neuron 19: 307–318, 1997. DAS A AND GILBERT CD. Long-range horizontal connections and their role in cortical reorganization revealed by optical recording of cat primary visual cortex. Nature 375: 780 –784, 1995. DINSE HR, SCHREINER CE, HILGER T, GODDE B, AND VON SEELEN W. Optical imaging of cat auditory cortex functional topographic organization using intrinsic signals. Assoc Res Otolaryngol Abstr 19: 460, 1996. DIRNAGL U, KAPLAN B, JACEWICZ M, AND PULSINELLI WA. Continuous measurement of cerebral cortical blood flow by laser-Doppler flowmetry in a rat stroke model. J Cereb Blood Flow Metab 9: 589 –596, 1989. EDVINSSON L, MACKENZIE ET, AND MCCULLOCH J. Cerebral Blood Flow and Metabolism. New York: Raven, 1993. FAGRELL B, INTAGLIETTA M, AND ÖSTERGREN J. Relative hematocrit in human skin capillaries and its relation to capillary blood flow velocity. Microvasc Res 20: 327–335, 1980. FROSTIG RD, LIEKE EE, TS’O DY, AND GRINVALD A. Cortical functional architecture and local coupling between neuronal activity and the microcirculation revealed by in vivo high-resolution optical imaging of intrinsic signals. Proc Natl Acad Sci USA 87: 6082– 6086, 1990. GHOSE GM AND TS’O DY. Form processing modules in primate area V4. J Neurophysiol 77: 2191–2196, 1997. GILBERT CD AND WIESEL TN. Clustered intrinsic connections in cat visual cortex. J Neurosci 3: 1116 –1133, 1983. GILBERT CD AND WIESEL TN. Columnar specificity of intrinsic horizontal and corticocortical connections in cat visual cortex. J Neurosci 9: 2432–2442, 1989. GOLANOV EV, YAMAMOTO S, AND REIS DJ. Spontaneous waves of cerebral blood flow associated with a pattern of electrocortical activity. Am J Physiol Regulatory Integrative Comp Physiol 266: R204 –R214, 1994. 1297 GRINVALD A, FROSTIG RD, LIEKE E, AND HILDESHEIM R. Optical imaging of neuronal activity. Physiol Rev 68: 1285–1366, 1988. GRINVALD A, LIEKE E, FROSTIG RD, GILBERT CD, AND WIESEL TN. Functional architecture of cortex revealed by optical imaging of intrinsic signals. Nature 324: 361–364, 1986. GRINVALD A, LIEKE EE, FROSTIG RD, AND HILDESHEIM R. Cortical point-spread function and long-range lateral interactions revealed by real-time optical imaging of macaque monkey primary visual cortex. J Neurosci 14: 2545– 2568, 1994. HARRISON RV, HAREL N, KAKIGI A, RAVEH E, AND MOUNT RJ. Optical imaging of intrinsic signals in chinchilla auditory cortex. Audiol Neurootol 3: 214 – 223, 1998. HEIL P, RAJAN R, AND IRVINE D. Topographic representation of tone intensity along the isofrequency axis of cat primary auditory cortex. Hear Res 76: 188 –202, 1994. HENDRY SH, HOCKFIELD S, JONES EG, AND MCKAY R. Monoclonal antibody that identifies subsets of neurones in the central visual system of monkey and cat. Nature 307: 267–269, 1984. HESS A AND SCHEICH H. Optical and FDG mapping of frequency specific activity in auditory cortex. Neuroreport 7: 2643–2647, 1996. HOCHERMAN S AND GILAT E. Dependence of auditory cortex evoked unit activity on interstimulus interval in the cat. J Neurophysiol 45: 987–997, 1981. HORIKAWA J, HOSOKAWA Y, KUBOTA M, NASU M, AND TANIGUCHI I. Optical imaging of spatiotemporal patterns of glutamatergic excitation and GABAergic inhibition in the guinea-pig auditory cortex in vivo. J Physiol (Lond) 497: 629 – 638, 1996. HORTON J AND HUBEL DH. A regular patchy distribution of cytochromeoxidase staining in primary visual cortex of the macaque monkey. Nature 292: 762–764, 1981. HUBEL DH AND LIVINGSTONE MS. Segregation of form, color, and stereopsis in primate area 18. J Neurosci 7: 3378 –3415, 1987. HUBEL DH AND WIESEL TN. Receptive fields of single neurones in the cat’s striate cortex. J Physiol (Lond) 148: 574 –591, 1959. HUBENER M, SHOHAM D, GRINVALD A, AND BONHOEFFER T. Spatial relationships among three columnar systems in cat area 17. J Neurosci 17: 9270 – 9284, 1997. HUMPHREY AL AND HENDRIKSON AE. Background and stimulus-induced patterns of high metabolic activity in the visual cortex (area 17) of the squirrel and macaque monkey. J Neurosci 3: 345–358, 1983. IMIG TJ AND ADRIAN HO. Binaural columns in the primary field (A1) of cat auditory cortex. Brain Res 138: 241–257, 1977. KITZES LM, WREGE KS, AND CASSADY JM. Patterns of responses of cortical cells to binaural stimulation. J Comp Neurol 192: 455– 472, 1980. LEVITT JB, YOSHIOKA T, AND LUND JS. Intrinsic cortical connections in macaque visual area V2: evidence for interaction between different functional streams. J Comp Neurol 342: 551–570, 1994. LIVINGSTONE MS AND HUBEL DH. Anatomy and physiology of a color system in the primate visual cortex. J Neurosci 4: 309 –356, 1984. LIVINGSTONE MS AND HUBEL DH. Specificity of intrinsic connections in primate primary visual cortex. J Neurosci 4: 2830 –2835, 1984. LIVINGSTONE MS AND HUBEL DH. Connections between layer 4B of area 17 and the thick cytochrome oxidase stripes of area 18 in the squirrel monkey. J Neurosci 7: 3371–3377, 1987. MALONEK D, DIRNAGL U, LINDAUER U, YAMADA K, KANNO I, AND GRINVALD A. Vascular imprints of neuronal activity: relationships between the dynamics of cortical blood flow, oxygenation, and volume changes following sensory stimulation. Proc Natl Acad Sci USA 94: 14826 –14831, 1997. MALONEK D AND GRINVALD A. Interactions between electrical activity and cortical microcirculation revealed by imaging spectroscopy: implications for functional brain mapping. Science 272: 551–554, 1996. MASINO SA, KWON MC, DORY Y, AND FROSTIG RD. Characterization of functional organization withinrat barrel cortex using intrinsic signal optical imaging through a thinned skull. Proc Natl Acad Sci USA 90: 9998 –10002, 1993. MAYHEW JE, ASKEW S, ZHENG Y, PORRILL J, WESTBY GW, REDGRAVE P, RECTOR DM, AND HARPER RM. Cerebral vasomotion: a 0.1-Hz oscillation in reflected light imaging of neural activity. Neuroimage 4: 183–193, 1996. MCCORMICK DA, CONNORS BW, LIGHTHALL JW, AND PRINCE DA. Comparative electro-physiology of pyramidal and sparsely spiny stellate neurons of the neocortex. J Neurophysiol 54: 782– 806, 1985. MENDELSON JR, SCHREINER CE, SUTTER ML, AND GRASSE KL. Functional topography of cat primary auditory cortex: responses to frequency-modulated sweeps. Exp Brain Res 94: 65– 87, 1993. 1298 SPITZER, CALFORD, CLAREY, PETTIGREW, AND ROE MERZENICH MM AND BRUGGE JF. Representation of the cochlear partition on the superior temporal plane of the macaque monkey. Brain Res 50: 275–296, 1973. MEYER JU, BORGSTRÖM P, LINDBOM L, AND INTAGLIETTA M. Vasomotion patterns in skeletal muscle arterioles during changes in arterial pressure. Microvasc Res 35: 193–203, 1988. MIDDLEBROOKS JC AND PETTIGREW JD. Functional classes of neurons in primary auditory cortex of the cat distinguished by sensitivity to sound location. J Neurosci 1: 107–120, 1981. MORITA-TSUZUKI Y, BOUSKELA E, AND HARDEBO JE. Vasomotion in the rat cerebral microcirculation recorded by laser-Doppler flowmetry. Acta Physiol Scand 146: 431– 439, 1992. MOUNTCASTLE VB, TALBOT WH, SAKATA H, AND HYVARINEN J. Cortical neuronal mechanisms in flutter-vibration studied in unanesthetized monkeys. Neuronal periodicity and frequency discrimination. J Neurophysiol 32: 452– 484, 1969. MURPHY KM, JONES DG, AND VAN SR. Cytochrome-oxidase blobs in cat primary visual cortex. J Neurosci 15: 4196 – 4208, 1995. NARAYAN SM, SANTORI EM, AND TOGA AW. Mapping functional activity in rodent cortex using optical intrinsic signals. Cereb Cortex 4: 195–204, 1994. PATEL U. Non-random distribution of blood vessels in the posterior region of the rat somatosensory cortex. Brain Res 289: 65–70, 1983. PETERSON BE, GOLDREICH D, AND MERZENICH MM. Optical imaging and electrophysiology of rat barrel cortex. I. Responses to small single-vibrissa deflections. Cereb Cortex 8: 173–183, 1998. PFINGST BE, O’CONNOR TA, AND MILLER JM. Response plasticity of neurons in auditory cortex of the rhesus monkey. Exp Brain Res 29: 393– 404, 1977. PHILLIPS DP. Effect of tone-pulse rise time on rate-level functions of cat auditory cortex neurons: excitatory and inhibitory processes shaping responses to tone onset. J Neurophysiol 59: 1524 –1539, 1988. PHILLIPS DP AND CYNADER MS. Some neural mechanisms in the cat’s auditory cortex underlying sensitivity to combined tone and wide-spectrum noise stimuli. Hear Res 18: 87–102, 1985. PHILLIPS DP, HALL SE, AND HOLLETT JL. Repetition rate and signal level effects on neuronal responses to brief tone pulses in cat auditory cortex. J Acoust Soc Am 85: 2537–2549, 1989. PHILLIPS DP AND IRVINE DRF. Responses of single neurons in physiologically defined area A1 of cat cerebral cortex: sensitivity to interaural intensity differences. Hear Res 4: 299 –305, 1981. PHILLIPS DP AND IRVINE DR. Methodological considerations in mapping auditory cortex: binaural columns in AI of cat. Brain Res 161: 342–346, 1979. PHILLIPS DP, KITZES LM, SEMPLE MN, AND HALL SE. Stimulus-induced spike bursts in two fields of cat auditory cortex. Hear Res 97: 165–173, 1996. PHILLIPS DP, SEMPLE MN, CALFORD MB, AND KITZES LM. Level-dependent representation of stimulus frequency in cat primary auditory cortex. Exp Brain Res 102: 210 –226, 1994. PRIETO JJ, PETERSON BA, AND WINER JA. Morphology and spatial distribution of GABAergic neurons in cat primary auditory cortex (AI). J Comp Neurol 344: 349 –382, 1994. REALE RA AND IMIG TJ. Tonotopic organization in auditory cortex of the cat. J Comp Neurol 192: 265–291, 1980. RESER DH, FISHMAN YI, AREZZO JC, AND STEINSCHNEIDER M. Binaural interactions in primary auditory cortex of the awake macaque. Cereb Cortex 10: 574 –584, 2000. ROCKLAND KS AND LUND JS. Intrinsic laminar lattice connections in primate visual cortex. J Comp Neurol 216: 303–318, 1983. ROE AW AND TS’O DY. Visual topography in primate V2: multiple representation across functional stripes. J Neurosci 15: 3689 –36715, 1995. SCHREINER CE AND MENDELSON JR. Functional topography of cat primary auditory cortex: distribution of integrated excitation. J Neurophysiol 64: 1442–1459, 1990. SCHREINER CE, MENDELSON JR, AND SUTTER ML. Functional topography of cat primary auditory cortex: representation of tone intensity. Exp Brain Res 92: 105–122, 1992. SENGPIEL F AND BLAKEMORE C. Interocular control of neuronal responsiveness in cat visual cortex. Nature 368: 847– 850, 1994. SHAMMA SA, FLESHMAN JW, WISER PR, AND VERSNEL H. Organization of response areas in ferret primary auditory cortex. J Neurophysiol 69: 367– 383, 1993. SHAMMA SA AND SYMMES D. Patterns of inhibition in auditory cortical cells in awake squirrel monkeys. Hear Res 19: 1–13, 1985. SHMUEL A AND GRINVALD A. Functional organization for direction of motion and its relationship to orientation maps in cat area 18. J Neurosci 16: 6945– 6964, 1996. SHOHAM D, HUBENER M, SCHULZE S, GRINVALD A, AND BONHOEFFER T. Spatio-temporal frequency domains and their relation to cytochrome oxidase staining in cat visual cortex. Nature 385: 529 –533, 1997. SILVERMAN MS, GROSOF DH, DEVALOIS RL, AND ELFAR SD. Spatial-frequency organization in primate striate cortex. Proc Natl Acad Sci USA 86: 711–715, 1989. SIMONS DJ. Response properties of vibrissa units in rat SI somatosensory neocortex. J Neurophysiol 41: 798 – 820, 1978. SPITZER MW AND CLAREY JC. Comparison of intrinsic optical and electrophysiological signals in cat auditory cortex. Soc Neurosci Abstr 24: 357.354, 1998. SUTTER ML AND SCHREINER CE. Topography of intensity tuning in cat primary auditory cortex: single-neuron versus multiple-neuron recordings. J Neurophysiol 73: 190 –204, 1995. TOOTELL RBH, HAMILTON SL, AND SILVERMAN MS. Topography of cytochrome oxidase activity in owl monkey cortex. J Neurosci 5: 2786 –2800, 1985. TOOTELL RBH, SILVERMAN MS, DEVALOIS RL, AND JACOBS GH. Functional organization of the second visual area in primates. Science 220: 737–739, 1983. TOTH LJ, RAO SC, KIM D-S, SOMERS D, AND SUR M. Subthreshold facilitation and suppression in primary visual cortex revealed by intrinsic signal imaging. Proc Natl Acad Sci USA 93: 9869 –9874, 1996. TS’O DY, FROSTIG RD, LIEKE EE, AND GRINVALD A. Functional organization of primate visual cortex revealed by high resolution optical imaging. Science 249: 417– 420, 1990. TS’O DY AND GILBERT CD. The organization of chromatic and spatial interactions in the primate striate cortex. J Neurosci 8: 1712–1727, 1988. VOLKOV IO AND GALAZJUK AV. Formation of spike response to sound tones in cat auditory cortex neurons: interaction of excitatory and inhibitory effects. Neuroscience 43: 307–321, 1991. VOLKOV IO AND GALAZYUK AV. Peculiarities of inhibition in cat auditory cortex neurons evoked by tonal stimuli of various durations. Exp Brain Res 91: 115–120, 1992. WALLACE MN, KITZES LM, AND JONES EG. Chemoarchitectonic organization of the cat primary auditory cortex. Exp Brain Res 86: 518 –526, 1991. WELIKY M, KANDLER K, FITZPATRICK D, AND KATZ LC. Patterns of excitation and inhibition evoked by horizontal connections in visual cortex share a common relationship to orientation columns. Neuron 15: 541–552, 1995. WONG-RILEY MTT. Changes in the visual system of monocularly sutured or enucleated cats demonstrated with cytochrome oxidase histochemistry. Brain Res 171: 11–28, 1979. WOOLSEY TA, ROVAINEN CM, COX SB, HENEGAR MH, LIANG GE, LIU D, MOSKALENKO YE, SUI J, AND WEI L. Neuronal units linked to microvascular modules in cerebral cortex: response elements for imaging the brain. Cereb Cortex 6: 647– 660, 1996. YOSHIOKA T, BLASDEL GG, LEVITT JB, AND LUND JS. Relation between patterns of intrinsic lateral connectivity, ocular dominance, and cytochrome oxidase-reactive regions in macaque monkey striate cortex. Cereb Cortex 6: 297–310, 1996. ZHENG D, LAMANTIA A-S, AND PURVES D. Specialized vascularization of the primate visual cortex. J Neurosci 11: 2622–2629, 1991.