Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Neural oscillation wikipedia , lookup
Subventricular zone wikipedia , lookup
Biochemistry of Alzheimer's disease wikipedia , lookup
Mirror neuron wikipedia , lookup
Electrophysiology wikipedia , lookup
Biological neuron model wikipedia , lookup
Neural coding wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Central pattern generator wikipedia , lookup
Caridoid escape reaction wikipedia , lookup
Multielectrode array wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Synaptogenesis wikipedia , lookup
Pre-Bötzinger complex wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Nervous system network models wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Circumventricular organs wikipedia , lookup
Neural engineering wikipedia , lookup
Synaptic gating wikipedia , lookup
Axon guidance wikipedia , lookup
Optogenetics wikipedia , lookup
Development of the nervous system wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Channelrhodopsin wikipedia , lookup
AMER. ZOOL., 28:1109-1122 (1988)
Structural Repair and Functional Recovery Following Cerebral
Ganglion Removal in the Pulmonate Snail Melampus1
STACIA B. MOFFETT AND RICHARD L. RIDGWAY2
Department of Zoology, Washington State University,
Pullman, Washington 99164-4220
SYNOPSIS. Regeneration of the nervous system of Melampus following cerebral ganglion
removal proceeds through tract, bud, and ganglion stages. Each stage can represent a
terminal condition in some animals. Early events of regeneration appear to include roles
for chemotactic and growth-promoting agents and axonal guidance by preferential adhesion to a connective tissue sheath. This latter proposed mechanism accounts for the
observed sequence in which the neural elements unite in the tract stage and for the pattern
of failures that result when the sheath is disrupted. In the tract stage of regeneration,
communication through the site of the missing ganglion is restored within the central
nervous system, and between neurons of non-excised ganglia and the denervated periphery. Some behavioral recovery results. The bud stage of regeneration is characterized by
neuropil development and associated swelling at the site of confluence of the tracts.
Serotonin immunohistochemistry of bud stage preparations and retrograde dye transport
via bud nerves show tracts and numerous synaptic varicosities, but the neuron somata that
are labeled are located in other ganglia. Ultrastructural examination of late bud/early
ganglion stage tissue reveals the presence of small undifferentiated cells. By six to seven
months postoperative, some snails have clearly reached a ganglion stage of regeneration
characterized by the appearance of differentiated neurons within the bud. The origin of
these new neurons is currently under investigation.
Koritzansky and Hartwig, 1974; Hulsebosch and Bittner, 1981) and in a single
mollusc, the snail Melampus bidentatus (Moffett and Austin, 1982). Why these few
species should be able to replace nerve cells
while other species cannot, is outside the
scope of this brief review (for a summary
of the main hypotheses see Hulsebosch and
Bittner, 1980). However, of the many factors that could contribute to the ease with
which a species would be able to regenerate
neurons, Melampus can claim several: (1) a
primitive evolutionary position among pulmonate gastropods, (2) a long lifespan, (3)
a potential to add neurons into adulthood,
and (4) a propensity for its tissues to respond
to neural induction.
Melampus is a high intertidal salt marsh
snail belonging to the lower order of pulmonate gastropods, the Basommatophora.
It is unusual among pulmonates in retaining the primitive feature of laying large
numbers of small eggs that hatch to a
planktonic veliger stage and eventually
1
From the Symposium on Nervous System Regener- undergo a metamorphosis into the adult
ation in the Invertebrates presented at the Annual Meet- form (Apley, 1970; Russell-Hunter el al.,
ing of the American Society of Zoologists, 27-30
1972). This reproductive strategy is augDecember 1986, at Nashville, Tennessee.
mented
by an iteroparous life cycle, i.e.,
* Present address: Dept. Med. Physiol., HSC, Univ.
one in which the parents live on to reproof Calgary, Alberta T2N 4N1 Canada.
INTRODUCTION
In terms of their capacity to regenerate,
molluscs are probably best known for being
able to replace various body parts. Tentacles, eyes, siphon, penis, and regions of
the foot, mantle, and shell, can all be
regenerated to a degree by most molluscs
(Hyman, 1967). There is evidence of, if not
a requirement for, the influence of neural
factors on regeneration in each of these
cases; damage to the nervous system itself
is typically repaired along with the associated body part. Many animal species are
unable to regenerate central nervous system (CNS) tissues in the absence of body
part replacement and it is especially rare
to encounter the ability to replace severely
damaged or lost neurons in adults (Hulsebosch and Bittner, 1980). In invertebrates
this ability has been documented only in a
few annelid species (Herlant-Meewis, 1962;
1109
1110
S. B. MOFFETT AND R. L. RlDGWAY
duce again and again (Calow, 1978). Melampus bidentatus is a slow growing species with a lifespan of greater than 3 yr in
field populations (Apley, 1970); under controlled laboratory conditions we have
maintained individuals for more than 6 yr.
Primitive features are also evident in the
anatomy of the digestive and reproductive
tracts of Melampus (Morton, 1955; Hubendick, 1978) and, more importantly for this
review, in the nervous system.
The CNS of pulmonate gastropods consists of eleven ganglia (five paired and one
unpaired) plus the commissures and connectives uniting them. A primitive characteristic of the Melampus nervous system
is the unfused ganglia and relatively long
connectives (Price, 1977a, 1979). Each
ganglion consists of a medullary neuropil
surrounded by a cortical rind of neurons
which in turn is encapsulated by a connective tissue sheath. The largest neurons of
gastropod ganglia are usually the most
peripheral and these appear to remain constant in number throughout life. Their
large size is primarily attributable to polyploidy (Lasek and Dower, 1971; Boer et al,
1977). Neuron addition into adulthood has
been noted in the CNS of molluscs, such
as in the opisthobranch gastropod Aplysia
californica, but this addition is apparently
confined to clusters of smaller cells bordering on the neuropil (Coggeshall, 1967).
Recent evidence suggests that this type of
neuron addition also occurs in Melampus
(May etal., 1987).
Melampus is well-adapted to withstand the
extremes of temperature, salinity, and oxygen tension associated with its harsh environment (McMahon and Russell-Hunter,
1981). Moreover, studies in our laboratory
continue to demonstrate the remarkable
regenerative capacity of Melampus tissues,
which includes the ability to produce
supernumerary sensory structures under
the influence of implanted ganglia (Moffett
and Austin, 1981). This demonstrates that
adult animals have the ability to completely
replace body parts, so regeneration of ganglia may simply represent one aspect of a
generalized capacity to access developmental information in response to injury.
In this article we review what is currently
known about cerebral tract and ganglion
regeneration in Melampus and present various possibilities for the mechanisms
involved.
EARLY EVENTS OF GANGLION
REGENERATION
Effects of cerebral ganglion ablations
The paired cerebral ganglia are among
the most important integrative centers in
pulmonate gastropods. Much of the information the animal receives about its environment passes through sensory tracts
associated with these ganglia (Janse, 1974).
Cerebral neurons direct feeding behavior
(Pentreath et al., 1982), locomotion (Snyder and Moffett, 1987), tentacle and head
movements (Lever et al., 1978), and the
whole body withdrawal response (Benjamin et al., 1985). Growth and reproduction
are also under control of cerebral ganglion
neurons (Joosse and Geraerts, 1983). The
survival time is brief when both cerebral
ganglia of Melampus are excised (Price,
19776). However, if both cerebral ganglia are removed and subsequently reimplanted, or removed and replaced by a pair
of cerebral ganglia from a donor, the snails
not only survive but recover behaviors
indicative of regeneration of cerebral circuitry (Moffett and Snyder, 1985; Moffett,
unpublished data). Support of survival and
regenerative growth by a single intact cerebral ganglion or by a pair of implanted
cerebral ganglia suggests that factors produced by these ganglia are critical for nervous system regeneration. This interpretation is supported by Wong and co-workers
(Wong et al., 1984) who have shown that
cultured neurons of the pond snail Helisoma trivolvis require media conditioned
with soluble and surface-active factors from
central ganglia for neurite outgrowth.
The initial tissue response to ganglion
ablation in molluscs is a period of reorganization, lasting about a week, that is characterized by reactive (chromatolytic) cell
bodies in the non-excised ganglia and
evidence of degeneration in the neuropil regions, cut nerves, and connectives
(Borovyagin et al, 1972; Moffett, 1980).
The degenerate tissue gradually disappears as regenerative growth begins. The
challenge then becomes that of directing
GANGLION REGENERATION IN MELAMPUS
1111
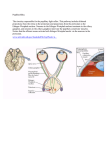
FIG. 1. Diagrammatical representation of the degree and sequence of nervous system regeneration in the
early period following removal of a left cerebral ganglion in Melampus bidentatus. A. Dorsal view of the CNS
with focus on the relative position of neural tracts (numbered structures) severed as a result of left cerebral
ganglionectomy (area delineated by the box). B-D. Examples of tissue regrowth exhibited by animals sacrificed
14 days (B), 28 days (C), and 70 days (D) post-lesion. The sequence of tissue regrowth leading to ganglion
reformation is suggested by the addition of regenerating neural tracts (shaded structures) to the presumptive
ganglion bud (asterisks) in each of the examples. Abbreviations and numbering: left and right cerebral ganglia
(LCe, RCe), cerebral commissure (1), tentacle nerve (2), optic nerve (3), cerebral tube/external peritentacular
nerve (4), anterior (internal) labial nerve (5), median labial nerve (6), labial artery (7), cerebrobuccal connective
(8), cerebropedal connective (9), cerebropleural connective (10). Not shown is the very thin statocyst (or static)
nerve which runs parallel to the cerebropedal connective. Redrawn from Price (19776) with modifications
based on observations of the authors.
the regrowing axons to the area where the
new ganglion will be established.
Axonal guidance by soluble factor gradients
Price (19776) discovered CNS regeneration in Melampus in the course of investigating the effects of ganglion ablation on
reproductive behavior. He observed that
after unilateral cerebral ganglionectomy,
the cut distal end of the medial labial nerve
(MLN; L, of Price, 19776) seemed to serve
as a focal point toward which the regenerating nerves and connectives grew. We
have confirmed these findings as well as
Price's suggestion that the regrowing tracts
arrive at the presumptive ganglion bud site
in a fairly predictable temporal sequence
as shown in Figure 1. The initial bud is a
swelling formed by the fused distal stumps
of the three labial nerves. A thin cerebral
commissure is usually reestablished by 14
days following ganglionectomy (Fig. IB).
Within 30 days the transected cerebropleural and cerebropedal connectives (and
statocyst nerve) fuse to form a single large
tract that joins the developing bud (Fig.
1C). By 70 days post-ablation the last of the
severed tracts, the cerebrobuccal connective and the tentacle and optic nerves have
reconnected (Fig. ID). The points of
attachment of these last structures are
highly variable so that only rarely does the
new ganglion have the same spatial
arrangement of tracts and nerves that is
characteristic of control ganglia.
Price (19776) hypothesized that a diffus-
1112
S. B. MOFFETT AND R. L. RlDGWAY
ible substance might be released from the
cut end of MLN to attract the regenerating
neural tracts. This hypothesis is bolstered
by reports that neurites can indeed be
guided by gradients of soluble molecules
{e.g., nerve growth factor: Letourneau,
1978) and by evidence that in several pulmonate gastropods the sheath of MLN
serves as a neurohaemal area for growth
hormone-producing neurosecretory cells.
In the pond snail Lymnaea stagnalis these
cells are known as "Light-Green Cells"
(LGC) from their appearance when stained
with alcian blue/alcian yellow dyes (Joosse
and Geraerts, 1983). We have shown that
cells probably homologous to the LGC are
present in Melampus and that their axons
likewise project to the sheath of MLN
(Ridgway, 1987). The growth hormone
has yet to be fully characterized, but immunohistochemical and chromatographical
evidence suggest that it may be related to
the vertebrate neuropeptide somatostatin
(Schot et al., 1981; Grimm-Jorgenson,
1983a, b; Ridgway and Moffett, 1987).
Moreover, recent experiments have shown
that exogenous somatostatin can enhance
outgrowth and electrical coupling of
regenerating molluscan neurons, possibly
by lowering intracellular free calcium concentrations (Grimm-Jorgenson, 1987; Bulloch, 1987).
the form and connectivity of the nervous
system in addition to their role in chemical
synaptic transmission (for a recent review
see Kater and Haydon, 1987). An even
broader morphogenetic role for neurotransmitters during regeneration is suggested by studies on transected flatworms.
The trauma of sectioning is thought to
abruptly lower intracellular free calcium
levels within the animal, causing a rise in
serotonin and noradrenaline concentrations (Martelly and Franquinet, 1984). This
sets in motion a cascade of intracellular
molecular events leading to the initiation
of DNA synthesis around 12 hr after transection. Likewise, the elevation of dopamine leads to the initiation of RNA synthesis 18-24 hr after transection (Martelly
and Franquinet, 1984). A similar sequence
of events may underlie the cellular activation observed early in the regeneration
of axotomized neurons of Melampus and
other organisms.
Axonal guidance by substrate adhesion
The tract regeneration following ablation of one cerebral ganglion in Melampus
does not follow the shortest path to the
presumptive bud site, as would be expected
if guidance by a soluble factor gradient were
the primary mechanism. Recent observations performed with the help of Dr. Keith
We are currently investigating the ability Snyder indicate that the initial guidance
of somatostatin and related substances to for many of the regrowing tracts is proact as chemotactic or growth-promoting vided by a connective tissue sheath (capiagents in Melampus CNS regeneration, but tocerebral membrane). It covers the CNS
there are many other substances, such as and fans out like a web between many of
serotonin, dopamine and glutamate, that the nerves. It surrounds much of the
could potentially serve a neurotrophic esophagus and extends into the periphery,
function or provide guidance for regen- providing protection and mechanical superating axons. For example, while gluta- port for the nervous system. We first noted
mate has been shown to enhance neurite the role of the sheath in cerebral commisoutgrowth and electrical synaptogenesis by surotomy regenerates (Figs. 2-6). We found
specific axotomized buccal ganglion neu- that when the sheath is interrupted as little
rons in Helisoma, serotonin and dopamine as possible, regeneration is rapid: a long
can inhibit regenerative activities (Hay- thin connection is usually present within
don et al, 1987; Bulloch and Jones, 1988). 7-10 days (Fig. 2B) and a distinct commisFurthermore, experimental depletion of sure formed by 1 mo postoperative (Fig.
serotonin during embryogenesis alters the 4). The regenerate commissure gradually
morphology and connectivity of identified thickens (and shortens) so that by 1 yr it
Helisoma neurons (Goldberg and Kater, closely resembles control tissue (compare
1985). Taken together, these findings Figs. 3 and 5). When the sheath was cut
strongly suggest that common neurotrans- far anterior, the axons growing out of the
mitters play a significant role in regulating ganglia followed the cut edge forward (Fig.
GANGLION REGENERATION IN MELAMPUS
1113
B
FIG. 2. Position of the cerebral commissure (arrows) and anterior portion of the connective tissue sheath
(shaded areas) as they appear in control preparations (A), in regenerate preparations after transection of only
the cerebral commissure (B), and in regenerate preparations after transection of the cerebral commissure
plus tearing of the connective tissue sheath (C). In both B and C, regenerating axons from the left and right
cerebral ganglia (LCe, RCe) follow the sheath margin to reestablish the commissure.
2C). When the sheath was torn extensively,
the forward-growing tracts often failed to
meet, and the commissure never formed
(Fig. 6). The sheath is usually the substrate
chosen by the earliest regenerating axons
in preference to the underlying esophageal
surface or other tissues adjacent to the ganglia. Subsequent regenerating axons often
appear to select the surfaces of the
"pioneer" axons as a substrate but may also
make use of the sheath.
The importance of the sheath can readily
be extended to the situation following
cerebral ganglion ablation. When a ganglion is removed there are 10 major tracts
that are severed as shown in Figure 7. Six
of these tracts (plus the statocyst nerve) are
held in place and are united by the sheath.
All of these tracts tend to use the sheath
as a substrate for growth toward the ganglion bud site. The other four tracts have
their connection with the sheath, as well
as their CNS connections, severed by ganglionectomy. These tracts are left as free
entities following ganglionectomy and cannot grow along a sheath edge to unite with
the rest of the nervous system. We feel this
is the main reason for the longer times
required for their reunification with the
bud (recall Fig. 1) and the high variability
in their final junction site.
The preference of regenerating tracts in
Melampus for the connective tissue sheath
may relate to differences in the adhesive
properties of surfaces available to axonal
growth cones. Studies in other animals have
demonstrated that the filopodial extensions of growth cones randomly explore
surfaces in the vicinity of the axon tip with
the result being growth in the direction of
greater adhesivity (Letourneau, 1985). It
is impossible to say at present what molecule^) provide the greater adhesivity in the
case of Melampus regeneration because the
constituents of the sheath are still incompletely described. Ultrastructural and histochemical studies have revealed that the
sheath consists primarily of an extracellular matrix (ECM) of collagen and other
products derived from fibroblast-like cells
and possibly glia (Ridgway, 1988). The
"other products" probably include glycoproteins, proteoglycans, and glycosaminoglycans organized into a network having
a specific structure and composition conducive to filopodial adhesion. A similar
hypothesis has been formulated to explain
the rapid and directed outgrowth of neurites from leech neurons cultured on CNS
capsule ECM extract as compared to the
slower, less directed, or unsuccessful
growth exhibited by neurons plated on
concanavalin A, poly-(L-)lysine, fibronectin, or laminin (Chiquet and Acklin, 1986).
The composition of the sheath ECM may
therefore determine the rapidity of early
neurite outgrowth and provide the necessary adhesion while soluble factor gradients and target influences may determine
the specific direction of the outgrowth.
Summary of the early events of
ganglion regeneration
The first response to cerebral ganglion
removal in Melampus is a brief period of
degeneration and reorganization followed
by the outgrowth of axons belonging to
1114
S. B. MOFFETT AND R. L. RlDGWAY
B
/
**.<*
6A
FIGS. 3-6. Examples of neural regeneration in animals where the anterior portion of the connective tissue
sheath was torn during cerebral commissure transection. In each example, A is an osmium tetroxide infiltration
whole mount (plastic embedment) showing the position of the cerebral commissure (CC) whereas B is a crosssection through the same commissure (asterisks) showing the position of the connective tissue sheath (S).
Figure 3 is a control preparation. Figures 4 and 5 are successful cerebral commissure regenerates sacrificed
at 1 mo post-lesion and 1 yr, respectively. Figure 6 is a failed cerebral commissure regenerate sacrificed at 4
mo post-lesion. The large arrowhead in 6A points to a thin, anteriorly directed growth that upon ultrastructural
examination was found to consist of only a very few neurites as shown in 6B. Magnification bars: 3A, 4A,
5A, 6A = 100 /im; 3B, 4B, 5B = 20 Mm; 6B = 1 Mm. Moffett and Ridgway (unpublished data).
neurons having their somata in non-excised
ganglia. Nerves and central tracts united
by incorporation within the connective tissue sheath grow together first, with the
sheath providing the preferred substrate
for initial neuritic growth and the distal
stump of the median labial nerve serving
as a focus for ganglion bud formation. The
result of these early events is reestablishment of central connections through the
site of the missing ganglion and between
remaining central neurons and the denervated periphery. This is the tract stage of
ganglion regeneration.
1115
GANGLION REGENERATION IN MELAMPUS
FIG. 7. Position of neural tracts (numbered structures) relative to the connective tissue sheath (shaded areas)
in situ (A) before and (B) after removal of the left cerebral ganglion (LCe). The sheath normally overlies the
CNS and fans out like a web, incorporating the cerebral commissure (1), labial nerves (5,6, and 7), cerebropedal
connective (9), statocyst nerve (not shown), and cerebropleural connective (10). When the left cerebral ganglion
is removed the sheath holds these tracts in place and provides a substrate for their regrowth (see text). Tracts
not normally incorporated by the sheath, such as the tentacle nerve (2), optic nerve (3), cerebral tube/external
peritentacular nerve (4), and cerebrobuccal connective (8) are left isolated in the blood space as a result of
ganglion removal.
100-r
UJ
O
51
80-
60-•
ID O
40--
S
20-
N
0 -112-24
(n = 10)
6-9
<n=16)
> 24
MONTHS
FIG. 8. Regeneration in snails with one cerebral ganglion removed was evaluated upon sacrifice two to more
than 24 mo after the lesion. The results were arbitrarily grouped into four time periods. The state of repair
was categorized as: no communication through the site of the missing ganglion (N), tract stage (T), bud stage
(B), or ganglion stage (G). The distribution within these four categories was plotted as percent of the total
number (n) of animals examined in each postoperative time period.
1116
S. B . MOFFETT AND R. L . RlDGWAY
FIG. 9. Transmission electron micrographs of regenerate left cerebral ganglia of Melampus. A. Neuropil
region of a 6 mo regenerate ganglion bud with neuritic processes bearing synaptic vesicles and neurosecretory
granules. B. A cluster of small undifferentiated cells in a 6 mo regenerate ganglion bud. C. Small differentiated
cells of a 12 mo regenerate ganglion. D. Large differentiated neuron of a 12 mo regenerate ganglion bearing
numerous cytoplasmic neurosecretory granules. Samples were prepared for electron microscopy as described
by Moffett and Austin (1982). Magnification bars: All bars are equal to 1 nm.
LATER EVENTS IN GANGLION
REGENERATION
Budformation
The morphology of ganglion regeneration has suggested three distinct stages:
tract formation, bud formation and gan-
glion formation (Price, 19776; Moffett and
Austin, 1982; Moffett and Snyder, 1985).
Each stage can represent a terminal con(Fig. 8). The
d i t i o n
in s o m e
anjmais
sequence of events in formation of the tract
stage has been described above. Despite
the loss of the ipsilateral sensory, motor
GANGLION REGENERATION IN MELAMPUS
1117
FIG. 10. Serotonin-like immunoreactivity in whole mounts of normal and regenerate left cerebral ganglia
of Melampus. A. A control cerebral ganglion contains 30-35 immunoreactive cells and a large number of
efferent and afferent processes. B. A 9 mo regenerate ganglion bud with profuse sprouting and growth of
neurites originating from neuron somata located in non-excised ganglia. C. A 15 mo regenerate ganglion
with a cluster of small cell bodies. This large cluster contains at least four immunoreactive cells (large
arrowheads); one cell having an axon (small arrowheads) that could be followed into the adjacent neuropil.
Samples were prepared by the indirect immunofluorescence (A, B) and indirect immunoperoxidase (C) methods as described by Ridgway (1988). Abbreviations: cell bodies (CB), cerebral commissure (CC), cerebropedal
connective (CPeC), median labial nerve (MLN), neuropil (NP), tentacle nerve (TN). Magnification bars: A =
50 fim; B = 50 /mi; C = 10 urn.
and interneuronal elements, a limited
recovery of behaviors mediated via cerebral pathways is seen as early as 3—4 wk
after ganglion removal. This roughly coincides with recovery of the normal extended
appearance of the tentacle on the operated
side, and the ability of this tentacle to
respond to stimulation with withdrawal
(Moffett and Snyder, 1985).
Bud formation results from neuropil
development and associated swelling at the
site(s) of confluence of the regenerating
1118
S. B. MOFFETT AND R. L. RlDGWAY
nerves and central tracts (Moffett and Austin, 1982; Ridgway and MofFett, 1985). This
occurs over a relatively long period and is
the typical condition encountered in animals sacrificed 2-9 mo after ganglionectomy. Ultrastructural examination of bud
stage tissue reveals an organized neuropil
region with many areas rich in neurosecretory granules and synaptic vesicles (Fig.
9A). Axonal projections traced by retrograde dye transport into the bud from bud
peripheral nerves reveal cells in other central ganglia that project through the bud,
but no somata within the bud (Moffett and
Snyder, 1985). Immunohistochemical
studies employing antisera to serotonin
show numerous varicosities ramifying
within the bud neuropil, but again no
labelled cell bodies (Fig. 10B). Nevertheless, the behavior of snails in this stage of
recovery typically includes robust tentacle
and labial withdrawal responses normally
mediated by cerebral neurons. Electrophysiological recordings combined with
acute transections that isolated the bud
from the rest of the CNS indicate that neuropil development within the bud is in some
instances sufficient to mediate reflexes on
the operated side. (Moffett and Snyder,
1985). The bud stage therefore supports
good behavioral recovery while relying on
the growth of neurites and elaboration of
synapses by neurons having their somata
in non-excised central ganglia and the
periphery.
Is the observed behavioral recovery in
the bud stage simply the result of regrowth
of neurons whose axons normally project
into the excised ganglion or are novel projections sent into the bud to compensate
for neurons lost due to ganglion ablation?
In the CNS of control snails, retrograde
dye transport (Kahan and Moffett, 1979)
and serotonin immunohistochemistry
(Ridgway, 1988) have shown that a percentage of the axons in cerebral nerves
normally arise from cells with bilateral or
contralateral somata (Fig. 10A). Thus, the
substrate for some recovery of function is
present after ganglion removal, even if
regeneration consists only of replacement
of normal projections by these cells.
Whether novel projections are made during ganglion regeneration in Melampus is
presently under investigation but studies
in several other gastropod species support
such a possibility. Following removal of the
right pleural ganglion in Lymnaea, regrowing neurites of an identified dopaminergic
neuron of the right pedal ganglion demonstrate highly specific regeneration to
original target areas but also extend into
regions of the CNS not normally occupied
by branches of this cell (Allison and Benjamin, 1985). The longevity, specific morphology (e.g., varicosities), and electrophysiology of these projections suggest that
novel, permanent synaptic connections are
formed (Benjamin and Allison, 1985).
Interestingly, direct damage is not necessary to elicit outgrowth by molluscan neurons (Bulloch, 1984; Allison and Benjamin,
1986; Bulloch and Jones, 1988), though in
such cases the projections are most often
transient. The extent to which these novel
projections can compensate for neurons lost
during ganglionectomy remains unclear.
The ga,7iglion stage
By definition, the regeneration of a ganglion would require the addition of neurons to the bud. In control ganglia, a rind
of neurons just beneath the sheath is visible
as distinct cells and cell clusters that give
the ganglion an overall yellow coloration.
We know from ultrastructural examination of late bud/early ganglion stage
regenerates that the first non-glial cells that
are detected are small, morphologically
undifferentiated cells usually present in
clusters (Fig. 9B). These presumptive
neurons (or neuron precursors) are not
encountered earlier than 5 mo after ganglion removal (Moffett and Austin, 1982).
By 7 mo, a population of differentiated
neurons can be present, characterized by
neurosecretory granules or synaptic vesicles, extensive Golgi apparatus and rough
endoplasmic reticulum (Fig. 9C, D). In
some instances of longer-term regeneration following cerebral ganglionectomy, we
have found small, serotonin immunoreactive somata (Fig. IOC). Thus, new cells
expressing the same phenotype as some of
the lost neurons can arise, indicating that
replacement of specific cells is possible in
Melampus.
The highly successful regenerated gan-
GANGLION REGENERATION IN MELAMPUS
glion appears to have all the features of a
normal, albeit slightly smaller ganglion,
complete with lobes and identifiable cell
clusters (Moffett and Austin, 1982). The
question arises as to how a nervous system
that had adjusted to the absence of a ganglion over a period of 6 mo can accommodate to the addition of newly generated
neurons. The problem can be posed by
considering the situation encountered by
new motor neurons. In order to be effective, these neurons must grow out and
assume control of peripheral structures that
were innervated by other neurons in their
absence, and attract the sensory and interneuronal inputs that had presumably been
concentrated upon the remaining neurons
after the original target cells were removed.
Experiments employing implanted ganglia
have shed light upon the ability of the
Melampus nervous system to accommodate
additional neurons (Moffett, 1980). One or
more cerebral ganglia implanted into the
hemocoel of a snail with an intact nervous
system can remain viable and form connections to the periphery and exchange
information with the host nervous system,
as revealed by recordings from the nerves
and ganglion surface. Implanted ganglia
are able to innervate regions of host tissues
that are already innervated, and the doubly
innervated region tends to enlarge. This is
particularly apparent when a host snail's
tentacle is innervated by an implanted ganglion. Induction of supernumerary sensory
structures appears to represent further evidence of the stimulatory and organizing
effect that the nervous system exerts on
other tissues (Moffett and Austin, 1981).
1119
following implantation of ganglia in
Melampus has shown that increasing the
mass of neural tissue can elicit an increase
in size of the target organ. Cerebral ganglion removal produces a mismatch
between the mass of cerebral nervous tissue and the mass of the tissues that are
normally innervated by cerebral neurons.
This mismatch may set in motion the cellular growth required to create a new balance, e.g., the generation of new ganglion
neurons.
Another possible source of new neurons
could be the neurons or other cell types
present in the non-excised central ganglia.
In annelids, migration of undifferentiated
stem cells from existing ganglia is the
mechanism whereby regenerating ganglia
are populated (Herlant-Meewis, 1962). We
know that there is an increase in the number of cells in certain cell clusters well on
into adulthood in Melampus (May et al.,
1987) and this phenomenon may be common among gastropods and other invertebrates (Hauser and Koopowitz, 1987).
Since glia, pigment cells and some classes
of neurons increase in number during
development, migration via commissure or
connectives would either supply the numbers of post-mitotic neuron precursors
needed or provide a stem cell population
that could increase by mitosis within the
regenerating ganglion.
One observation about ganglion regeneration that is apparent from the data in
Figure 8 is that some snails do not progress
beyond the tract or bud stage even if
allowed more than 2 yr of regeneration
time. The proportion of snails exhibiting
We do not yet know where the cells that regeneration of a ganglion does not change
are destined to become new neurons arise. between the 12-24-mo period and the over
One possibility is that there is a recapitu- 24-mo period, whereas in the 6-9-mo
lation of developmental events, such as period the bud stage rather than the ganthose recently described for Aplysia glion stage predominates. We therefore
(McAllister et al., 1983; Jacob, 1984). Neu- plan to study snails in the 6-9-mo period
ron precursors divide in the body wall and of regeneration to determine the origin(s)
migrate to establish the central ganglia, of the new neurons.
with the cerebral ganglia being the first
formed. These migratory cells are thought Summary of the later events of
to have withdrawn from the cell cycle. ganglion regeneration
However, there is evidence that at least The later stages of regeneration seem to
some post-migration mitosis occurs within be dependent upon the success of earlier
the developing ganglia (see Fig. 9 ofJacob, developments, including first the unifica1984). The growth of sensory structures tion of most or all the central tracts, the
1120
S. B. MOFFETT AND R. L. RlDGWAY
addition of a neuropil to form a bud, and
the population of the bud by neuron somata
to produce a ganglion. In instances in which
the cerebral commissure failed to unite the
regenerating tracts of the left side with the
intact right cerebral ganglion, for instance,
regeneration never has been found to progress beyond the tract or possibly the bud
stage. This implies that physical guidance
is an important mechanism in promotion
of regeneration in Melampus. Neuron
somata have rarely been found in situations
in which unification of nerves and central
tracts had produced an elongated structure
or one with more than one site of enlargement. We therefore think that a favorable
architecture may promote the development of interactions within the regenerating bud that in some way may be required
for more advanced stages of regeneration.
Clearly, replacement of neurons is a longterm process which cannot depend upon
the relatively short-term stimulation of
regenerative growth that is associated with
injury.
CONCLUSION
The phenomenon of CNS regeneration
in Melampus provides us with an opportunity to determine what is required for successful repair of injury involving neuron
loss. Regeneration following ganglion
removal proceeds through a sequence of
stages, and behavioral analysis indicates
that attainment of each successive stage
affords a greater measure of recovery. This
report and others in this symposium have
dealt with factors that stimulate neuron differentiation and growth and are involved
in guidance and the formation of connections. New techniques in the areas of biochemical analysis, cell culture, electrophysiology, and imaging, should allow the
identification of the factors responsible for
repair at each stage of recovery. Besides
being intrinsically interesting, we can hope
that investigations on nervous system
regeneration in invertebrates such as
Melampus will shed light on basic phenomena that may aid in promotion of the process in mammals.
ACKNOWLEDGMENTS
We thank Dr. Keith Snyder for his contributions to the work presented and Dr.
David F. Moffett, Ronald May, and Tamara
Howard for critical reading of the manuscript. We also thank Daniel Austin for
contributing Figure 9B. This research was
supported by grants from the National
Institutes of Health (5 R01 NS 14333 and
1 R01 NS 22896) to S. B. Moffett and by
Grants-in-Aid of Graduate Research from
Sigma Xi and Washington State University
to R. L. Ridgway.
REFERENCES
Allison, P. and P. R. Benjamin. 1985. Anatomical
studies of central regeneration of an identified
molluscan interneuron. Proc. R. Soc. London B
226:135-157.
Allison, P. and P. R. Benjamin. 1986. Stimulation of
neuritic outgrowth in an undamaged molluscan
interneuron. J. Exp. Biol. 122:447-451.
Apley, M. L. 1970. Field studies on life history,
gonadal cycle and reproductive periodicity in
Melampus bidentatus(Pu]monala: Ellobiidae). Malacologia 10:381-397.
Benjamin, P. R. and P. Allison. 1985. Regeneration
of excitatory, inhibitory and biphasic synaptic
connections made by a snail giant interneuron.
Proc. R. Soc. London B 226:159-176.
Benjamin, P. R., C. J. H. Elliott, and G. P. Ferguson.
1985. Neural network analysis in the snail brain.
In A. Selverston (ed.), Model neural networks and
behavior, pp. 87-108. Plenum Press, New York.
Boer, H. H., C. Groot, M. Dejong-Brink, and C. J.
Cornelisse. 1977. Polyploidy in the freshwater
snail Lymnaea stagnalis (Gastropoda, Pulmonata).
A cytophotometric analysis of the DNA in neurons and some other cell types. Neth. J. Zool. 27:
245-252.
Borovyagin, V. L., J. Salanki, and I. Zs.-Nagy. 1972.
Ultrastructural alterations in the cerebral ganglion of Anodonla cygnea L. (Mollusca, Pelecypoda) induced by transection of the cerebro-visceral connective. Acta Biol. Acad. Sci., Hung. 23:
31-45.
Bulloch, A. G. M. 1984. Sprouting and retraction of
neurites by undamaged adult molluscan neurons.
Brain Res. 321:369-373.
Bulloch, A. G. M. 1987. Somatostatin enhances neurite outgrowth and electrical coupling of regenerating neurons in Helisoma. Brain Res. 412:617.
Bulloch, A. G. M. and P. G.Jones. 1988. Modulation
of neurite outgrowth by intact and axotomized
neurons of Helisoma. Amer. Zool. 28:1123-1134.
Calow, P. 1978. The evolution of life-cycle strategies
in fresh-water gastropods. Malacologia 17:351364.
GANGLION REGENERATION IN MELAMPUS
Chiquet, M. and S. E. Acklin. 1986. Attachment to
Con A or extracellular matrix initiates rapid
sprouting by cultured leech neurons. Proc. Natl.
Acad. Sci. U.S.A. 83:6188-6192.
Coggeshall, R. E. 1967. A light and electron microscope study of the abdominal ganglion of Aplysia
californka. J. Neurophysiol. 30:1263-1287.
Goldberg, J. I. and S. B. Kater. 1985. Experimental
reduction of serotonin content during morphogenesis alters morphology and connectivity of
specific identified Helisoma neurons. Soc. Neurosci. Abstr. 11:58.
Grimm-Jorgensen, Y. 1983a. Immunoreactive somatostatin in two pulmonate gastropods. Gen.
Comp. Endocrinol. 49:108-114.
Grimm-Jorgensen, Y. 1983i. Possible physiological
roles of thyrotropin releasing hormone and a
somatostatin-like peptide in gastropods. In J.
Lever and H. H. Boer (eds.), Molluscan neuroendocrinology, pp. 21-28. North-Holland, Amsterdam.
Grimm-Jorgensen, Y. 1987. Somatostatin and calcitonin stimulate neurite regeneration of molluscan neurons in vitro. Brain Res. 403:121 — 126.
Hauser.M.andH. Koopowitz. 1987. Age-dependent
changes in fluorescent neurons in the brain of
Notoplana acticola, a polyclad flatworm. J. Exp.
Zool. 241:217-225.
Haydon, P. G., D. P. McCobb, and S. B. Kater. 1987.
The regulation of neurite outgrowth, growth cone
motility, and electrical synaptogenesis by serotonin. J. Neurobiol. 18:197-215.
Herlant-Meewis, H. 1962. Neurosecretory phenomena during regeneration of nervous centers in
Eiseniafoetida. In H. Heller and R. B. Clark (eds.),
Neurosecretion, pp. 267-273. Academic Press, New
York.
Hubendick, B. 1978. Systematics and comparative
morphology of the Basommatophora. In V. Fretter and J. Peake (eds.), Pulmonates, Vol. 2A, Sys-
1121
pulmonate snail Melampus. Zoomorphologie 94:
81-91.
Kater, S. B. and P. G. Haydon. 1987. Multi-functional roles for neurotransmitters: The regulation of neurite outgrowth, growth cone motility
and synaptogenesis. In A. Vernadakis (ed.), Model
systems ofdevelopment and aging of the nervous system,
pp. 239-255. Martinus Nijhoff, Norwell, Massachusetts.
Koritsanszky, S. and H. G. Hartwig. 1974. The regeneration of the monoaminergic system in the
cerebral ganglion of the earthworm Allolobophora
caliginosa. Cell Tissue Res. 151:171-186.
Lasek, R. J. and W. J. Dower. 1971. Aplysia californka: Analysis of nuclear DN A in individual nuclei
of giant neurons. Science 172:278-280.
Letourneau, P. C. 1978. Chemotactic response of
nerve fiber elongation to nerve growth factor.
Dev. Biol. 66:183-196.
Letourneau, P. C. 1985. Axonal growth and guidance. In G. M. Edelman, W. E. Gall, and W. M.
Cowan (eds.), Molecular bases ofneu ral development,
pp. 269-293. John Wiley and Sons, New York.
Lever, A. J., H. J. Bruins, C. Geigenhuis, W. M. Everts,
and B. S. Fokkema. 1978. The neural organization of the tentacle contraction reflex of the
pond snail Lymnaea stagnalis (L.). Proc. Kon. Ned.
Akad. Wetensch., Ser. C 81:265-278.
Martelly, I. and R. Franquinet. 1984. Planarian
regeneration as a model for cellular activation
studies. Trends Biochem. Sci. 8:468-471.
May, R. H., R. L. Ridgway, and S. B. Moffett. 1987.
Size and number of serotonin immunoreactive
cells change with age in adults of the snail Melampus. Soc. Neurosci. Abstr. 13:1070.
McAllister, L. B., R. H. Scheller, E. R. Kandel, and
R. Axel. 1983. In situ hybridization to study the
origin and fate of identified neurons. Science 222:
800-808.
McMahon, R. F. and W. D. Russell-Hunter. 1981.
The effects of physical variables and acclimation
tematics, evolution, and ecology, pp. 1-47. Academic
on survival and oxygen consumption in the high
Press, London.
littoral salt-marsh snail, Melampus bidentatus Say.
Hulsebosch,C. E. and G. D. Bittner. 1980. Evolution
Biol. Bull. 161:246-269.
of abilities to regenerate neurons in central ner- Moffett, S. 1980. Ultrastructure of regenerating cenvous systems. Am. Nat. 115:276-284.
tral nervous system connections and implanted
Hulsebosch, C. E. and G. D. Bittner. 1981. Regenganglia in the snail Melampus. Soc. Neurosci.
eration of axons and nerve cell bodies in the CNS
Abstr. 6:386.
of annelids. J. Comp. Neurol. 198:77-88.
Moffett, S. and D. R. Austin. 1981. Implanted cereHyman, L. H. 1967. The invertebrates (Vol. 6): Molbral ganglia produce supernumerary eyes and
lusca I. McGraw-Hill, New York.
tentacles in host snails. J. Exp. Zool. 216:321 —
Jacob, M. H. 1984. Neurogenesis in Aplysia californka
325.
resembles nervous system formation in verte- Moffett, S. and D. R. Austin. 1982. Generation of
brates. J. Neurosci. 4:1225-1239.
new cerebral ganglion neurons in the snail
Melampus: An ultrastructural study. J. Comp.
Janse, C. 1974. A neurophysiological study of the
Neurol. 207:177-182.
peripheral tactile system of the pond snail Lymnaea stagnalis (L.). Neth. J. Zool. 24:93-161.
Moffett, S. and K. Snyder. 1985. Behavioral recovery
associated with central nervous system regenerJoosse.J. and W. P. M. Geraerts. 1983. Endocrinolation in ihesmWMelampus.]. Neurobiol. 16:193—
ogy. In A. S. W. Saleuddin and K. M. Wilbur
209.
(eds.), The Mollusca, Vol. 4, Physiology, Part 1, pp.
317—406. Academic Press, New York.
Morton, J. E. 1955. The evolution of the Ellobiidae,
with a discussion on the origin of the Pulmonata.
Kahan, L. B. and S. Moffett. 1979. Cobalt mapping
Proc. Zool. Soc, London 125:127-168.
of neurons with processes in pedal nerves in the
1122
S. B. MOFFETT AND R. L. RlDGWAY
like immunoreactivity in normal and regenerating cerebral ganglia of the snail Melampus. Soc.
Neurosci. Abstr. 11:421.
pods. In N. N. Osborne (ed.), Biology ofserotonergic
transmission, pp. 457-513. John Wiley and Sons, Ridgway, R. L. and S. B. Moffett. 1987. Substance
New York.
P-like and somatostatin-like immunoreactivity in
Price, C. H. 1977a. Morphology and histology of the
the CNS of the snail Melampus. Soc. Neurosci.
central nervous system and neurosecretory cells
Abstr. 13:1071.
in Melampus bidentatus Say (Gastropoda, Pulmo- Russell-Hunter, W. D., M. L. Apley, and R. D. Hunter.
nata). Trans. Am. Microsc. Soc. 96:295-312.
1972. Early life history of Melampus and the significance of semilunar synchrony. Biol. Bull. 143:
Price, C. H. 19776. Regeneration in the central ner623-656.
vous system of a pulmonate mollusc, Melampus.
Cell Tissue Res. 180:529-536.
Schot, L. P. C, H. H. Boer, D. F. Swaab, and S. van
Price, C. H. 1979. Physical factors and neurosecreNoorden. 1981. Immunocytochemical demontion in the control of reproduction in Melampus
stration of peptidergic cells in the pond snail Lym(Mollusca, Pulmonata). J. Exp. Zool. 207:269naea stagnalis with antisera raised to biologically
282.
active peptides of vertebrates. Cell Tissue Res.
Ridgway, R. L. 1987. Alcian blue-alcian yellow
216:273-291.
mapping of neurosecretory cells in the central Snyder, K. A. and S. Moffett. 1987. Loss and recovnervous system of the salt marsh pulmonate snail
ery of locomotor behavior after CNS lesions in
Melampus bidentatus. Comp. Biochem. Physiol.
the snail Melampus bidentatus. Biol. Bull. 172:8387A:295-303.
88.
Ridgway, R. L. 1988. Ultrastructure, histochemis- Wong, R. G., D. L. Barker, S. B. Kater, and D. A.
try, and aspects of regeneration of the cerebral
Bodnar. 1984. Nerve growth-promoting factor
ganglia of the salt marsh snail Melampus bidentatus
produced in culture media conditioned by speSay (Pulmonata, Basommatophora). Ph.D. Diss.,
cific CNS tissues of the snail Helisoma. Brain Res.
Washington State University, Pullman.
292:81-91.
Ridgway, R. L. and S. B. Moffett. 1985. SerotoninPentreath, V. W., M. S. Berry, and N. N. Osborne.
1982. The serotonergic cerebral cells in gastro-