Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extrachromosomal DNA wikipedia , lookup
Messenger RNA wikipedia , lookup
Cancer epigenetics wikipedia , lookup
Epigenetics of neurodegenerative diseases wikipedia , lookup
Gene expression programming wikipedia , lookup
Gene desert wikipedia , lookup
Oncogenomics wikipedia , lookup
Epigenetics of diabetes Type 2 wikipedia , lookup
X-inactivation wikipedia , lookup
Human genome wikipedia , lookup
Epigenomics wikipedia , lookup
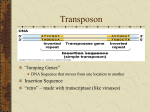
Transposable element wikipedia , lookup
Minimal genome wikipedia , lookup
Genomic imprinting wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
No-SCAR (Scarless Cas9 Assisted Recombineering) Genome Editing wikipedia , lookup
Epigenetics in learning and memory wikipedia , lookup
Deoxyribozyme wikipedia , lookup
Genetic engineering wikipedia , lookup
Epitranscriptome wikipedia , lookup
Gene expression profiling wikipedia , lookup
Non-coding RNA wikipedia , lookup
Short interspersed nuclear elements (SINEs) wikipedia , lookup
Genome evolution wikipedia , lookup
Genome (book) wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Long non-coding RNA wikipedia , lookup
Genome editing wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Designer baby wikipedia , lookup
History of genetic engineering wikipedia , lookup
Non-coding DNA wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Helitron (biology) wikipedia , lookup
Microevolution wikipedia , lookup
Transcription factor wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Point mutation wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics Mutational Dissection Gene Regulation Cell Number: Cancer Sex Determination Complex Pattern Formation NextGen Sequencing Bioinformatics Group Members Neil Brown Andrew Bullen Brian Gibbons Special thanks to Linda Lait for all her help with the figures Basics and Logic of prokaryotic transcription regulation: A great example of regulation of gene transcription comes from prokaryotes and their regulation of metabolic enzymes to break down sugars (lactose, glucose, etc.) if all enzymes where transcribed it would be too energetically costly Therefore the must control transcription based on the presence of what sugar is present. They must have the ability to turn on and off specific groups of genes in response to environmental conditions. Basics of prokaryotic regulation: There are two types of DNA-protein interactions that are necessary for transcription regulation. One being the interaction between RNA polymerase and the promoter region of a gene. the next DNA-protein interaction determines if the transcription caused by the promoter region even occurs Regulation of the lactose system: first look Thanks to the work by Jaques Monond and François Jacob we know a lot about the lactose regulation system and the catabolic repression in the presence of glucose. The metabolism of lactose uses two enzymes: a permease (for lactose transport into the cell) and β-galactosidase (to cleave lactose into glucose and galactose) These enzymes are encoded on a single multigenic mRNA molecule and thus regulation of this mRNA regulates the production of these enzymes. Carabolite Repression of the lac Operon Glucose provides more energy to the cell than other sugars High glucose levels inactivate the enzyme adenylate cyclase which converts ATP to cAMP cAMP is an allosteric effector of CAP (catabolite activator protein) cAMP - CAP complex is an activator which binds to the lac promoter region Hamer et al. 1993 Genetic Aspects of the lac Operon Model Genetic consequences of mutations in various components of the lac operon provide a better understanding of how it works Properties of mutations in structural genes and regulatory elements are quite different Techniques for Mutational Analysis To study the repressor and operator, we consider enzymatic assays for ß-galactosidase (Z) and permease (Y) Genotypes are assayed in the uninduced state (no inducer present) and induced state Uninduced state tests whether genetic circuitry necessary for repression is present Induced state tests whether genetic circuitry necessary to overcome repression is present In all experiments, no glucose is present (no catabolic repression) Natural inducers such as lactose are not used as their concentration decreases over time Synthetic inducers that bind to the allosteric site of the repressor but are not cleaved by ß-galactosidase are used Bacteria heterozygous for desired mutation are created using F' factors carrying the lac region of the genome Results of Genetic Analysis Table 13-1 Table 13-2 Genetic evidence of allostery Repressor mutations are important here. Inhibits transcription when no inducer but permits transcription when the inducer is present. This is the job of a specific location in the repressor. Mutations in this area change the way the repressor acts and essentially voids the action of the inducer when it's present. This "super repressor" is denoted with IS and is dominant to the wild type Mutant IS binds to all operators in the cell and prevents transcription of the lac operon since all inducers are ignored due to the super repressor. This allosteric interaction shows the importance of the repressor being in proper working order Structures of target DNA sites The binding sites in the operon show an interesting pattern. Rotational twofold symmetry (match when spun 180°) Hints to the importance of symmetry and homodimers/homotetramers in the binding of proteins in DNA Summary The lac operon allows bacteria to adjust the way they obtain energy based on what's available in their surroundings. Works via the binding of repressor protein to the DNA, preventing transcription. Repressor activity is controlled by the presence of both glucose catabolites CAP-cAMP) and lactose sugar RNA polymerase requires both a shortage of catabolite and the presence of lactose sugar. Only produces the proper enzymes when needed and useful, avoids being wasteful. The inducer repressor system is an example of negative control, or repression. Expression is normally blocked. The CAP-cAMP system is an example of activation control where expression requires activation. The Arabinose Operon An example of dual positive and negative control. The protein AraC can assume two different conformations: 1. When bound to arabinose, the AraC-arabinose complex binds to the AraI and acts an activator. 2. In the absence of arabinose, AraC acts as a repressor, binding to AraI and AraO, forming a DNA loop which prevents transcription. The arabinose operon also uses the CAP-cAMP complex to regulate metabolism in the presence of glucose. Eukaryotic Regulation Same basic tasks as prokaryotes, but done differently and more intricately Respond to changes in time (development), space (tissue differentiation) and conditions (physiology) Uses a lot of the same mechanisms as prokaryotes, but their greater complexity makes it necessary to have more complex controls as well. Eukaryotic genomes are also larger and typically have many more genes Greater regulation is also necessary to achieve finer control, this therefore increases the number of necessary regulatory sites. Cis-Acting sequences Up until now we have talked about regulatory mechanisms that act in trans with their gene of interest. RNA synthesis requires polymerases to act on the strand. Key areas of interactions have been identified in many genes, these can be promoter or silencer sites and they have different effects Promoters increase transcription rates Silencers decrease transcription rates They are usually classed by their proximity to the gene of interest There are two main classes of promoter, promoter-proximal elements and distance independent. Promoter proximal elements are generally very close to the transcription start site (within 200 bp) and can have very profound effects Usually requires the activation of multiple proximal elements to have a very large effect Distance independent elements can act many base pairs away from their target, 100s-1000s of base pairs away This distance is usually though to be covered by the creation of "folds" in the DNA strand Enhancer or silencer sites act as binding sites for proteins which can greatly affect the transcription rate of the mRNA. Trans Control of Transcription As in prokaryotes, trans-acting regulatory proteins in eukaryotes bind to specific target DNA sequences. Before initiation of transcription by RNA polymerase II, an initiation complex consisting of regulatory proteins is formed at the promoter (TATA box). Transcription factor IID (TFIID) is the first protein to bind to the promoter. It contains a subunit called TBP which binds to the TATA box, and more than 8 additional subunits (TAFs). Two additional regulatory proetins, TFIIA and TFIIB, bind to the TFIID promoter complex before the binding of a preformed RNA polyermase II-TDIIF complex. Finally, TFIIE, TFIIH, and TFIIJ must be added to the complex, in that order, for transcription to be initiated. The TFII complexes are oftern referred to as basal or general transcription factors, are they are required for RNA polymerase II to initiate transcription. Tissue Specific Regulation of Transcription Conferred in one of two ways: 1. The activator for an enhancer may only be present in some types of cells. 2. A tissue-specific repressor can bind to a silencer element near the enhancer, making the enhancer inaccessible to its transcription factor. In some genes, regulation can be controlled by a simple set of enhancers: In female Drosophila, vitellogenins (egg yolk proteins) are made in the ovary and fat body. Enhancers located within a few hundred base pairs of the promoter regulate the vitellogenin gene, one for the ovary and one for the fat body. In other genes, regulation can be very complex, controlling complex patterns of gene expression: The dpp (decapentaplegic) gene in Drosophila encodes a protein which mediates cell-cell communication. The number of enhancers for this gene may number in the hundreds, dispersed along a 50 000 base pair interval of DNA. Each of these enhancers regulates dpp gene expression at a different site in the developing animal. Dissecting Eukaryotic Regulatory Elements In order to find exact locations of specific regulatory elements enhancers of a cloned gene of interest are indentified via reporter genes Reporter genes are pieces of cis-regulatory DNA that are fused to a transcription unit that can express the reporter protein- often producing an observable effect (color, glow etc.) As a example LacZ is easily detected as the addition of X-gal, will be converted to a blue product by β-glactosidase. Via these means the DNA sequence of the enhancer can be identified by whittling down the piece of cis-regulatory DNA to smaller and smaller fragments. Regulatory Elements and Dominant Mutations We have already seen the importance of regulatory sites, it is not unsurprising to imagine the problems that could happen were they to be mutated When the regulatory elements of one gene are moved to another by mutations there can be great consequences In development we see this a lot, while such an extreme change is often fatal to the developing embryo, it can lead to interesting phenotypes The Tab (transabdominal) mutation in drosophila is a great example of a non-lethal mutation. In this case part of the thorax causes part of the thorax of the fly to develop like the sixth abdominal segment. Tab is associated with a chromosomal inversion. One of the break points is at the enhancer region of the similar sr (striped) gene and the other is near the transcription unit of the Abd-B (Abdominal-B) In the chromosomal breakage the inversion causes the abdominal genes to be turned on wherever the striped genes normally would. Structure and function of transcription factors Some of these factors need to travel long distances from their site of origin to their eventual target Factors need to be specific but also flexible in their approaches to binding As seen by the cis acting sequences, the sequence specificity is a very important part of this binding process. Many interacting proteins have core elements bound to DNA binding elements In these proteins certain elements fit into the major or minor grooves of the DNA helix to facilitate binding The Helix-Turn-Helix motif is a common one in these proteins as well. It allows a stable adapter between the DNA sequence and the protein The Zinc finger motif is also an important binding pattern in DNA protein interactions. They can interact with highly specific sections of sequence There are also sections which interact in their activities to produce a greater effect then they would singly. When enough of these enhancing proteins join together they form an enhanceosome Enhanceosomes can greatly increase the output of mRNAs for a particular gene of interest Paramutation In this phenomenon, seemingly normal alleles, termed paramutable alleles are irreversibly changed after being present in the same genome as paramutagenic alleles. In corn, the B-I gene produces individuals with normal pigmentation, but the B' mutation produces nearly colourless individuals. A heterozygous individual is nearly colourless, thus one would think B' is dominant to B-I. However, a cross between two heterozygous individuals does not produce a 1:3:1 progeny. All offspring are colourless. Parental Imprinting Another form of epigenetic inheritance is Parental imprinting Imprinted genes are expressed as if they where hemizygous (one copy present) even though there are two copies of the genes present. There are no changes to the DNA but are extra methylated at certain bases; active genes are less methylated then inactive ones. Because the allele from one parent is inactivated the other will appear dominant, and is often the cause of genetic disease such as Prader-Wili syndrome (PWS) X-Inactivation in Mammals In mammals, the amount of most X chromosome gene products must be made equal to the single X chromosome in males. Accomplished by inactivating one of the X chromosomes in each cell at an early stage, propogating this inactive state to all progeny cells. Two aspects of this phenomenon reflect imprinting: 1. Selection of the X chromosome is not always random. Marsupials always inactivate parental X. All other mammals make the decision randomly at the 64-cell stage of development. 2. The selection is "remembered" as stated above. This memory is correlated to methylation of the inactive X, less acetylation of a histone subunits on the inactive X, and a special RNA called Xist. Xist is synthesized by the inactive X chromosome and is thought to create an inactivation centre. The RNA appears to spread along the chromosome. The mechanisms behind these systems is yet to be determined. Interesting applications of these ideas These transcription factors can have great effects on the endocrine system in the human body, they are important regulatory factors and messengers The hormonal messengers produced by the endocrine glands are great examples of how transcription factors can act across great distances and have a drastic effect on the host organism The steroid hormones (estrogen, testosterone and glucocorticoids) for example can travel long distances to their region of interest where they can then pass through the cell membranes and regulate transcription factors in the nucleus When we look at anabolic steroids, the "artificial" performance-enhancing drug derived from natural compounds These drugs stimulate muscle growth, however the long term effects are very detrimental to human health, including increased cancer rates and potentially reproductive complications. The drug acts on muscle building genes, however it also has a suite of other effects In Genomics we can seek to find a "transcriptome" for an organism under various conditions. This can range from heat shocks to toxicological studies. These genomics studies often look at specific tissues and their expression of various genes over the whole genome to get a perspective on whole organism response to stresses Enter supporting content here Steve Carr (instructor: [email protected]) *** Linda Lait (TA: [email protected]) Dept of Biology, Memorial University of Newfoundland, St John's NL A1B3X9