Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Signal transduction wikipedia , lookup
Cognitive neuroscience of music wikipedia , lookup
Cortical cooling wikipedia , lookup
Central pattern generator wikipedia , lookup
Convolutional neural network wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Neural engineering wikipedia , lookup
Animal echolocation wikipedia , lookup
Response priming wikipedia , lookup
Synaptic gating wikipedia , lookup
Sensory cue wikipedia , lookup
Optogenetics wikipedia , lookup
Time perception wikipedia , lookup
Neuroethology wikipedia , lookup
Sensory substitution wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
C1 and P1 (neuroscience) wikipedia , lookup
Biological neuron model wikipedia , lookup
Microneurography wikipedia , lookup
Nervous system network models wikipedia , lookup
Development of the nervous system wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Psychophysics wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Perception of infrasound wikipedia , lookup
Neural coding wikipedia , lookup
Evoked potential wikipedia , lookup
Efficient coding hypothesis wikipedia , lookup
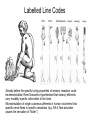
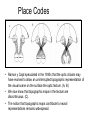
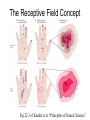
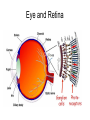
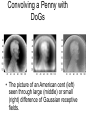
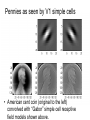
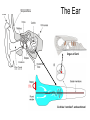
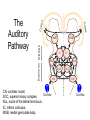
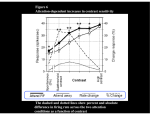
Principles of Sensory Neuroscience Systems Biology Doctoral Training Program Physiology course Prof. Jan Schnupp [email protected] HowYourBrainWorks.net Transduction Fig 2 of http://www.masseyeandear.org/ research/ent/ent-investigators/eatock/ Labelled Line Codes Already before the specific tuning properties of sensory receptors could be demonstrated, Rene Descartes hypothesized that sensory afferents carry modality specific information to the brain. Microstimulation of single cutaneous afferents in human volunteers links specific nerve fibres to specific sensations (e.g. RA-II fibre activation causes the sensation of “flutter”) Place Codes • Ramon y Cajal speculated in the 1930s that the optic chiasm may have evolved to allow an uninterrupted topographic representation of the visual scene on the surface the optic tectum. (A, B) • We now know that topographic maps in the tectum are discontinuous. (C). • The notion that topographic maps contribute to neural representations remains widespread. The Receptive Field Concept Fig 22.3 of Kandel et al “Principles of Neural Science” A Better Receptive Field Concept… … describes the behaviour of a sensory neuron quantitatively in terms of a “transfer function” y=f(x) which maps a mathematical description of the stimulus x (location, intensity, frequency, colour, temperature, recent history …) onto a measure of the neuron’s “output” y (depolarization, firing probability, response latency). Rate (Hz) Rate Codes 150 100 50 0 0 1 2 Weight (g) 3 • Classic experiments performed by Adrian in the 1920s on frog muscle stretch receptors established that sensory afferents use changes in spike rate to signal the intensity of a stimulus. • Adrian also found that many sensory neurons “adapt”, i.e. they do not maintain very high firing rates for long if stimuli are held constant. Quantifying Rate Codes: The Post Stimulus Time Histogram Source: http://www.frontiersin.org/Journal/10.3389/fnsyn.2010.00017/full Eye and Retina Centre –Surround Receptive Fields Photoreceptors ++ + + -- - - Horizontal Cell - Bipolar Cell Retinal Ganglion Cell Difference of Gaussians Model of Retinal Ganglion Cells • The centre-surround structure of Retinal Ganglion Cells turns them into “spatial frequency filters”. Larger RGC receptive fields are tuned to “coarsely grained” structure in the visual scene, while smaller RFs are tuned to fine grain structure. Convolving a Penny with DoGs • The picture of an American cent (left) seen through large (middle) or small (right) difference of Gaussian receptive fields. Seeing Lines The Gabor Filter Model of V1 Simple Cells LGN - + - + -+ - Cortex + - + - + - + Retina->LGN->V1 simple cell: linear Linear Filters in Visual Cortex? Movshon, Tolhurst and Thompson 1978 • The “F0/F1” ratio is often used to distinguish simple (approximately linear) V1 neurons from complex (nonlinear) ones. • Responses are recorded to sinusoidal contrast gratings. If the cell is linear, the output should contain only the input frequency F0. • Fourier analysis is performed on the post stimulus time histogram to measure the amplitude ratio of the fundamental (1st harmonic, F1) to the “zero frequency” (i.e. sustained, “DC” response) F0. • Some complex cells have “on” and “off” responses which manifest themselves as F2=2·F1 components - a “quadratic” (... +c·sin(x)2 +...) non-linearity. Pennies as seen by V1 simple cells • American cent coin (original to the left) convolved with “Gabor” simple cell receptive field models shown above. The Ear Organ of Corti Cochlea “unrolled” and sectioned “Gammatone Filter Bank” Auditory Nerve Fibers behave like Rectified Gammatone Filters Auditory Neuroscience Fig 2.12 Based on data collected by Goblick and Pfeiffer (JASA x Corte x Corte MGB Brainstem Midbrain The Auditory Pathway MGB IC IC NLL CN, cochlear nuclei; Cochlea SOC, superior olivary complex; NLL, nuclei of the lateral lemniscus; IC, inferior colliculus; MGB, medial geniculate body. NLL SOC CN SOC CN Cochlea Linear Neural Filters In Auditory Cortex? From work by Shihab Shamma and colleagues Freq. channel Measuring Frequency-Time (Spectro-Temporal) Receptive Fields with Reverse Correlation time Binaural Frequency-Time Receptive Field Linear Prediction of Responses Input “i vector” 16 Frequency [kHz] 4 1 16 4 1 Latency response r(t) = i1(t-1) w1(1) + i1(t-2) w1(2)+ ... + i2(t-1) w2(1) + i2(t-2) w2(2)+ ... + i3(t-1) w3(1) + i2(t-2) w3(2)+ ... FTRF “w matrix” 1 0.5 0 200 ms 100 0 -5 0 5 10 dB Predicting Space from Spectrum Left and Right Ear Frequency-Time Response Fields a Virtual Acoustic Space Stimuli 16 Frequency [kHz] 4 1 d Elev[deg] [deg] Elev 4 1 c Schnupp et al Nature 2001 -5 0 5 10 dB 1 0 -60 -180 -120 -60 100 0 e 0.5 0 200 ms 60 rate (Hz) response b C81 16 0 f 60 120 180 Azim [deg] 200 0 0 100 ms 200 Are Neurons “Noisy” Rate Coders? Or Precision Spike Timers? Mainen & Sejnowski, Science 1995 • • • Many nervous in the central auditory system seem to fire only short bursts of action potentials at the onset of a stimulus. For such neurons, the response latency may vary as a function of certain stimulus parameters (e.g. intensity, sound source position … ) and could therefore encode that parameter. Nelken et al, “Encoding stimulus information by spike numbers and mean response time in primary auditory cortex” J Comput Neurosci (2005) Sound source azimuth () What about Spike Latency Codes? How about Spike Interval Codes? The discharges of cochlear nerve fibres to low frequency sounds are not random; they occur at particular times (phase locking). The spike time intervals therefore encode temporal features of the stimulus (sound periodicity). Evans (1975) Who reads the neural code, and how do they do it?