Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Proprioception wikipedia , lookup
Patch clamp wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Nonsynaptic plasticity wikipedia , lookup
Electromyography wikipedia , lookup
Development of the nervous system wikipedia , lookup
Membrane potential wikipedia , lookup
Single-unit recording wikipedia , lookup
Synaptic gating wikipedia , lookup
Nervous system network models wikipedia , lookup
Signal transduction wikipedia , lookup
Action potential wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Resting potential wikipedia , lookup
Node of Ranvier wikipedia , lookup
Biological neuron model wikipedia , lookup
Neurotransmitter wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Electrophysiology wikipedia , lookup
Microneurography wikipedia , lookup
Neuroregeneration wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Chemical synapse wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Synaptogenesis wikipedia , lookup
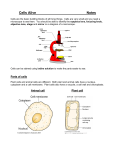
Name_______________________________ EXCITABLE TISSUES LECTURE MANUAL (BY2202) 2011/2012 SF (2nd yr) Biology Students Professor Kumlesh K. Dev Department of Physiology This Manual is produced in conformity with College’s policy on accessibility and may be requested in other formats (e.g. electronic, large print) Note. In the event of any conflict or inconsistency between the general regulations published in the University Calendar and information contained in this Handbook, the provisions of the General Regulations will prevail. I 1 NOTES 1. Lessons There are 5 Lessons that will cover the information given in the Lecture and Lab. You are expected to read the Lessons 1‐5 before the lab. There will be a 15 min lab assessment at the end of the lab and the questions will be related to the Lessons and the Lab. 2. Lab You should only come to the laboratory during the time of your group. Please be aware of your group and come to the laboratory on the correct time and day. Note that there will be a 15 min assessment at the end of each lab group and you are expected to have read the Lessons 1‐5 before the lab. 3. Lectures There will be 2 Lectures associated with the Lessons and Labs. Please refer to your timetable for details. I 2 LESSON 1: EXCITABLE NERVE TISSUES 1. Objectives To be able to: a) describe structure of processes, fibres, synapses of nerve cells b) outline stages of communication between nerve cells (neurotransmission) c) describe myelinated and nonmyelinated nerve axons 2. Introduction Muscle & Nerve: together the Excitable Tissues, characterised by response to external stimuli; part of response is electrical. Nerve cells are adapted to conduct electrical impulses (action potentials). Transmission between one nerve terminal and another or an end‐organ is by chemical transmission across the synaptic gap. It is important to distinguish nerve cell bodies, nerve processes (axons, dendrites), bundles of nerve fibres and nerve fibres. The nerve cell body and its processes comprise the neuron. The function of nervous tissue is to integrate stimuli, generate and transmit action potentials. Nerve cells (neurones) are very variable in size and shape, but essentially are relatively large cells with processes. The main process is the axon, smaller processes are dendrites. Axons contact an end‐organ (effector) or another neuron. Dendrites contact other neurons. Motoneurons have up to 1000 dendritic connections each. Neurons typically have relatively large, pale nuclei, each with a prominent nucleolus. Often only the ‘cell body’ is seen in section. The cytoplasm contains many granules which are clusters of free ribosomes (the high RNA content makes them basophilic). The prominent nucleolus and abundant free ribosomes together indicate a high level of synthesis. Nuclei are long‐lived and often have long axons, so they may have a large mass of cytoplasm to maintain; hence the need to have a capacity for synthesis of cellular components. A typical motor neurone receives about 1000 connections, reflecting the general function of neurones of integrating inputs and giving a single output, the action potential. A neurone’s connections may be relatively short and non‐myelinated (‘dendrites’) or longer (axons). Glial cells provide structural and physiological support for neurones and lie between blood vessels and neurone. About 90% of CNS cells are glial. 3. Nerve Fibres The vast majority of nerve fibres in the body are very thin (<2µm) and invisible with the light microscope. These extensions of the cell body may be very long as in the case of those from motor neurones to the lower limb. Thicker axons become myelinated, that is, they acquire a wrapping of Schwann cell cytoplasm giving an insulating layer. (The Schwann cell is a special type of glial cell.) Thinner axons (the vast majority) remain nonmyelinated. Myelin is an effective insulator and assists the rapid conduction of the action potential from node to node. The larger the axon, the thicker the myelin and the faster the speed of conduction. I 3 All neurons are have associated glial cells which have a very close metabolic relationship and are ‘supporting’ (trophic) cells. The glial cells cover all the surface of the nerve cell body. Glial cells have important immunological functions. 4. Synapses The junction between a neuron and an effector or other neuron is a synapse which has a gap across which the stimulus is passed by a chemical transmitter. This means that there is a synaptic delay and the synapse acts as a filter or gate for the stimulus. This is an important part of the cleaning up of the signal. I 4 LESSON 2: NERVE CELLS 1. Introduction There are three types of neurons. Motor neuron, which runs from CNS to effector such as muscle. Sensory neuron, which runs from a receptor to the CNS. Interneuron, which link other neuron within the CNS (they are often short but can be very complicated especially in the brain). The basic characteristics of neurons are listed below. A neuron contains 1. Cell body (soma) 2. Dendrites (dendritic tree) that carry signals to soma 3. Axon hillock 4. Axon that may be more than 1m long and carry signals away from soma, except in sensory fibres where it is reverse 5. Colaterals 6. Axonal branches and terminals Neurons contain the usual organelles 1. Nucleus with nucleolus 2. Mitochondria, which are also found in the axon terminals. 3. Endoplasmic reticulum with ribosomes (rough ER) 4. Golgi apparatus 5. Neurofibrils that add support 6. microtubules which are important for axonal transport 2. Supporting cells Only about 10% of brain cells are neurons. A large numbers of supporting cells known as glial (or neuroglia) are present. There are 4 types of these cells, these include astrocytes, microglia, oligodendrocytes and ependymal cells.. Astrocytes: take up and recycle certain neurotransmitters and growth factors, and their perivascular feet surround blood vessels & help maintain blood‐brain barrier. Oligodendrocytes (in CNS) and Schwann cells (in periphery) that myelinate neurons. Microglia are of the macrophage lineage (they are fixed macrophages) and act as phagocytic scavengers for defence. Ependymal cells line cavities in brain and spinal cord. 3. Nerve Conduction 1. Nerve Coding The nervous system only has one unit of code ‐ the Action Potential (AP). Nervous information must convey information about the nature of the signal (eg light, temperature, pressure etc) and the strength of the stimulus (eg how bright the light, how high the temp). Nature of the signal (modality) depends on anatomy. Nerve impulses from eye are interpreted as light (even if caused by some other method ‐ punch in eye). Nerve impulses to muscle contract muscle even if elicited electrically. Strength is encoded as frequency of AP ‐ the faster the frequency the stronger the signal and the number of fibres used (recruitment). I 5 2. Action potential A change in electrical potential in a neuron can stimulate voltage gated Na+ channels to open, which cause an influx of Na+ into the neuron causing a further potential change across the membrane opening more channels. Eventually with a strong enough stimulus (supra‐threshold) there is positive feedback and a large number of Na+ channels open (Na+ Gate). In this state the Na+ permeability is some 20 times greater than potassium permeability. Since Na+ carries a positive charge into the cell the inside of the membrane becomes positive. The potential rises to almost Na+ equlibrium potential (+60mV) to about +40mV. This sudden depolarisation of the membrane opens more K+ channels (also voltage gated). K+ permeability increases (K+ gate) and at the same time Na+ permeability returns to below normal (to inactivated state). Thus cell membrane repolarises. In fact the potential actually drops slightly below the resting membrane potential, for a few milliseconds ‐ hyperpolarisation. This whole process of events is known as the Action Potential. It lasts about 1 ms. in nerve (longer in muscle). It is important to note that very few ions actually move so many APs can occur in rapid sucession up to 200/sec for many sec before any serious depletion occurs. The action potential is propergated along the axon. This action is an ALL OR NONE event. 3. Refractory periods During and shortly after an AP the membrane cannot be excited again. This is the Absolute Refractory Period . The membrane then gradually returns to normal Relative Refractory Period. Early in the RRP the membrane can be excited but it requires a much higher voltage (stimulus). 4. Myelin sheath Many neurones (but NOT all) are insulated with a layer of Myelin, which is created by Oligodendrocytes in the CNS and by Schwann cells in peripheral nerves. The glial cell wraps itself around the axon and the fatty Myelin is secreted in the folds. Oligodendrocytes add Myelin sheaths to several axons. Schwann cells to only one. There are gaps in the myelin sheath known as Nodes of Ranvier 5. Un‐myelinated fibres Currents flow around the depolarised area. These flow through the next bit of the membrane and alter its permeability to Na+ ions. The next bit of membrane depolarises. Every tiny increment of the membrane must be depolarised, and depolarisation is a relatively slow process (0.5ms). So conduction is slow. Speed depends on Diameter of fibre. Conduction In both directions but refractory period prevents change of direction. Unmyelinated fibres is only about 1 m/s. Clearly this is too slow for large animals (2 sec from brain to foot!). Speed can be increased by making the fibres larger. Some invertebrate giant fibres conduct at > 10 m/s. Squid giant fibre reaches 20 m/s but it is almost 1mm in diameter. Can’t have many of these as nerves become very large. 6. Myelinated fibres Currents flow around the depolarised area. But current cannot flow through the myelin sheath. Therefore current flows to the next Node of Ranvier (the inter‐node section acting as an electric cable). So only a few I 6 depolarisations take place ‐ so much faster conduction. Speed depends on: Diameter of fibre and Internode distance. Thus myelinated fibres conduct over 100 times faster than the same size unmyelinated fibre. A fibre only 10µm in diameter can reach a speed of almost 100 m/s. The advantages of myelination are: faster conduction, reduced energy expenditure (less ion movement across membrane so less ions have to be pumped back). I 7 LESSON 3: NEUROMUSCULAR JUNCTION Neurons communicate across cell‐cell junctions known as synapses. The junction between a nerve and a muscle fibre is known as a Neuromuscular junction (NMJ), and is a special kind of synapse. 1. Muscle Innervation 1. Motor unit Each motor nerve fibre divides into many terminal branches (unmyelinated), each of which forms one NMJ with ONE muscle fibre. Each muscle fibre only has one NMJ. The group of muscle fibres innervated by one motor nerve fibre, together with that fibre, is called a MOTOR UNIT. Large motor units may have 1000 muscle fibres. Small fine control muscles (eg orbital muscles) may have as few as 10 fibres per motor unit. 2. Mechanism of action The end of a motor nerve fibre forms a swelling (Terminal Knob) which is imbedded in the surface of the muscle fibre. The muscle membrane is folded to increase its surface area (Motor end plate). The neuron and muscle membranes are separated by a narrow cleft. The action potential in the neuron cannot cross this gap. The neuronal terminal knob is well supplied with mitochondria and with small vesicles that contain neurotransmitters. The depolarisation of the neuron terminal knob causes the opening of voltage gated Ca2+ channels. Ca2+ enters the neuron (since it is in low conc inside & high outside). The Ca2+ ions trigger reactions which cause the vesicles containing neurotransmitters to migrate towards the junction membrane. Some vesicles fuse with the neuronal surface membrane and burst releasing their neurotransmitter content into the cleft. The neurotransmitter in this case is Acetylcholine (ACh). The ACh diffuses rapidly across the narrow cleft and attaches to receptors on the muscle membrane. These receptor operated channels (ROCs) are chemical‐gated and thus differ from voltage gated channels (VOCs). These receptors are connected to cation channels for example Na+ and K+ which allow these ions to pass across the muscle membrane (esp. Na+, which is driven by both concentration and electrical gradients) thus depolarising the muscle membrane. This depolarisation is known as the end plate potential (EPP). If the NMJ is partially blocked with curare no action potential is seen but these small local EPP are recorded. The EPP is always supra‐threshold so always initiates an AP in the muscle. The AP is initiated in the region on either side of the end plate. The AP passes along the muscle fibre (in both directions) as in unmyelinated nerve fibre. Importantly, the ACh only stays bound to its receptor molecules for a very short time, then it is released to either bind to another receptor or finds its way to the bottom of the folds and bind to the enzyme acetylcholineesterase which breaks it down. 3. Quantal Release of Ach Even in the absence of stimulation, odd vesicles of ACh are released giving tiny depolarisations of the muscle membrane (≈400µV) known as miniature end plate potentials (MEPPs). If the NMJ is almost completely I 8 blocked, so that each AP only releases a very few vesicles, EPPs can be seen to occur in steps of 400µV, indicating the quantal release of Ach from each vesicle . 2. Toxins and Disease The neuromuscular junction is very susceptible to toxins, drugs & diseases. Curare binds to the ACh receptor preventing ACh attaching. Botulinum toxin (food poisoning) blocks release of Ach. In both these cases the muscles stay relaxed. Black widow venom causes excess release of ACh leading to prolonged depolarisation and contraction. Organophosphates (in pesticides and nerve gas) irreversibly bind to the ACh esterase thus preventing breakdown of Ach In these cases the muscles stay contracted. These toxins are mainly dangerous because of their effect on the respiratory muscle which must contract & relax rhymically to maintain oxygen supply & sustain life. Myasthenia gravis is an autoimmune condition which depletes ACh receptors leading to muscle weakness. Neostigmine is an anticholinesterase drug that counteracts above condition. I 9 LESSON 4: SYNAPTIC TRANSMISSION 1. Mechanim of transmission The mechanism of neurotransmission at nerve‐nerve synapses (also known as a general synapse) are similar to NMJ. The following events occur: 1. Pre‐synaptic AP depolarises the terminal knob 2. Opens voltage‐gated Ca2+ channels 3. Vesicles of transmitter migrate to pre‐synaptic membrane and burst. 4. Transmitter crosses cleft and attaches to receptors on post‐synaptic membrane 5. These receptors are bound to chemical‐gated channels 6. Ions cross post‐synaptic membrane and change its trans‐membrane potential There are some key differences between nerve‐nerve synapses and NMJ: 1. Nerve‐nerve synapses are much smaller, so potential change/AP is far less 2. No folding of neuronal post‐synaptic membrane ‐ smaller surface area 3. Synapses introduce a delay (≈ 0.5 ‐1.0 mS) which is significant in complex circuits 4. In Nerve‐nerve synapses transmitter substance is not always ACh. There are many known transmitters 5. One AP hardly ever raises trans‐membrane potential to threshold. It usually takes many presynaptic APs to reach threshold and produce a post‐ synaptic AP. Potential is therefore lower, usually sub‐threshold in nerve‐ nerve synapses and requires summation 2. Summation The cell bodies of the postsynaptic neuron receive many synaptic endings from many per‐synaptic fibres (up to 100,000!). If 1 presynaptic AP is not sufficient to cause a post‐synaptic AP there must be summation to raise the Membrane potential to threshold. Each depolarisation of a terminal knob produces a small local (graded) excitatory postsynaptic potential (EPSP). Since the resistance of the soma is very low the EPSP spreads throughout the soma. 1. Spatial summation Most presynaptic neurones have more than one ending, so contribute more to the membrane potential (EPSP). Usually many presynaptic fibres have to fire together to raise the membrane potential to threshold. 2. Temporal Summation Alternatively one (or a few) fibres may fire repeatedly at high frequency. The individual EPSPs will summate and may raise the membrane potential (EPSP) to threshold. 3. Inhibition Not all presynaptic fibres are the same. Some have a different transmitter, which opens either K+ or Cl‐ channels, and hyper‐polerises the membrane giving rise to an inhibitory postsynaptic potential (IPSP). IPSPs summate by spatial & temporal summation but have opposite effect (cancelling out EPSPs). I 10 3. Neurotransmitters There are a number of known neurotransmitters and many more suspected ones. Some are always excitatory, some always inhibitory. Some may be excitatory or inhibitory depending on the synapse. However each ending only has 1 neurotransmitter and is either excitatory or inhibitory. Neurotransmitters do not survive for long in the synaptic cleft. They are either broken down by enzymes or taken back into the presynaptic terminal to be recycled. Many neuropeptides are also released by presynaptic neurones. They are produced in the soma, passed down to the terminal and released along with the transmitter. Their effect is slow and long‐term, usually enhancing or supressing the effects of tramsmitters by altering availability of transmitter or receptors. 4. Receptors It is necessary to get information INTO the system (from the environment or body) and OUT OF the system (as muscular work). The body is well supplied with sensory receptors of many types: − Chemo receptors Taste/smell, O2/CO2/pH − Photo receptors Vision − Mechano receptors touch, pressure, balance, motion, blood pressure, hear − Thermo receptors skin − Osmo receptors blood osmolarity − Nociceptors Non‐specific nerve endings ‐ pain The general mechanism is − The stimulus brings about a change in permeability of membrane to small ions. Na+ in particular moves into the cell (as its concentration and electrical gradients are strongest) − Na+ movement brings about a depolarisation − These potentials are graded potentials akin to EPSPs. If stimulus is strong enough they will exceed threshold and initiate an action potential. The Frequency of Action potentials depends on the size of the potential. Hence frequency signals stimulus strength (Frequency code). Stimulus strength may also be reflected in Number of endings which reach threshold, for example a light touch fires one ending and a heavy prod excites several (Population code). The modality of the stimulus is signaled by the anatomical arrangement of the nerves (where they come from & where they go to) (Place Code) eg. blow to the eye gives ‘stars’. Receptor desensitisation/adaption: Many sensory receptors do not keep firing at the same rate if stimulus is maintained. They adapt. Tonic receptors show little or no adaptation (eg joint position receptors or temperature receptors. Phasic receptors adapt rapidly to a constant stimulus (eg skin touch receptors). 5. Summary The EPSP of the post‐synaptic soma does not remain at resting membrane potential for long. It fluctuates up and down depending on the nett input from all the presynaptic fibres acting together by spatial & temporal summation of EPSPs & IPSPs. In this way synaptic connections are far more than just junctions between nerve fibres. Furthermore, the amount of transmitter released by one ending may be altered by presynaptic inhibition/facilitation from a second neuron. These processes give the CNS its powers of calculation and computation. I 11 LESSO ON 5: EXCITABLE X E MUSCL LE TISSUES 1. Obje ectives To be ab ble to: a) de escribe the e structure e, innervation and fu unction of the differe ent ty ypes of mus scle (smootth, cardiac and skeleta al) b) co omment on n their appe earance in section s c) de escribe the fibre-types s of skeleta al muscle d) de efine a mottor endplate and motor u unit e) exxplain fibre‐ttype groupin ng 2. Intro oduction Muscle is adapted for contracction. There are three types t of muscle: smoo oth, cardiac a and skeletal.. They can be considered d as a series of increasing g specialisattion for conttraction. Exccitable Tissu ues respond to approprriate stimuli by generatting electricaal responses. In the casee of muscle, the electricaal response is translated d to contracttion. There are three types of muscle m whicch can be seen s as beeing increasin ngly specialised for contraction. Figu ure Diagrram of Muscle Types. a) Skkeletal Muscle; b b) Cardiac Muscle; c) Smooth m muscle. The represent right‐‐hand pictures r transverse sections o of cells/ffibres. [Passmore & Robsoon (1968). Fig.155.1] 3. Smo ooth Muscle Smooth muscle con nsists of small spindle‐sshaped slen nder cells with w a relativvely undevelo oped contraactile apparaatus. The ceells are not striated. The cells tend d to aggregaate into bund dles or sheeets (e.g. gut)). Each cell has h a single nucleus in tthe slightly w wider centree. Note that because thee cells are much longer tthan the nucclei, long, maany profiles o of transversee sections will not contaiin a nucleus. ‐ In nnervation Du ual innervattion with bo oth excitatory and inhib bitory inputss. The balan nce beetween them m gives rise to a partial state of con ntraction or t tone. In sing gle‐ un nit muscle (e.g. gut, uteerus) not all cells are directly innerrvated; insteead th hey are coup pled more or less indirecttly, so that e excitation sp preads from t the directly‐innervvated cells to t those thaat do not recceive innervaation. In mu ulti‐ nit muscle (ee.g. airways, large arteriees) each cell is innervated d. un I 12 ‐ Function Relatively weak slow, spreading contraction (obviously related to the above properties. The contraction of each cell is graded (cf. ‘all‐or none’ of cardiac /skeletal muscle). 4. Cardiac Muscle Cardiac muscle consists of large cells or fibres which are closely attached to one another end‐to‐end at intercalated discs (modified Z discs). The fibres are striated indicating an elaborate well developed contractile apparatus. Fibres may branch and have electrical continuity; may be binucleate. The nuclei are centrally placed in the large cells, so that nuclei are not visible in all profiles. The electrical junctions between cells are indicated by modified Z discs; the intercalated discs. Contraction is faster than in smooth muscle and all‐or‐none. Not all cells are directly innervated and electrical excitation spreads through the tissue via electrical continuity and also by specialised cardiac muscle cells, the Purkinje fibres. The latter are pale because they are rich in glycogen which is unstained in conventional (H&E) preparations. ‐ Innervation Within the heart pacemaker cells control the rate of contraction and excitation spreads via the left and right branch bundles of Purkinje fibres: modified cardiac muscle cells containing glycogen. ‐ Function These muscles are non‐fatiguing, powerful, co‐ordinated contraction; rich in mitochondria; rich blood supply. 5. Skeletal Muscle Skeletal muscle consists of large fibres, each of which is formed by the fusion of primitive cells; thus multinucleate with relatively prominent cross‐striations. The contractile apparatus almost completely fills the fibres and is very regularly arranged to give the characteristic striations. The many nuclei are at the very edges of the fibres (i.e. peripheral). ‐ Innervation Each muscle fibre receives a specialised nerve terminal i.e. a single axon of an alpha motor neurone forming a motor endplate. The area of the fibre immediately below the endplate is modified; a specialised form of neuromuscular junction, characterised by much folding of the post‐synaptic (muscle) surface to give a large surface area for transmission. One spinal motor neurone (motoneuron) can innervate hundreds or even thousands of muscle fibres in a given muscle. These fibres comprise the motor unit. All the fibres of a motor unit contract at the same time and are all of the same type (below). Contractions are powerful due to the density of the contractile apparatus and are all‐or‐none. There are many motor units in a muscle; force is increased by increasing ‘recruitment’. ‐ Function These muscles are fast, powerful, ‘all or nothing’ (‘twitch’) contraction. I 13 6. Muscle‐Fibre Types Skeletal muscle has essentially two types of fibre according to enzyme profile and physiological characteristics, such as velocity of contraction and resistance to fatigue. The principal enzymes used are succinic dehydrogenase (SDH), a mitochondrial oxidative enzyme, and myofibrillar ATPase (mATPase). The Table below summarises the enzyme and physiological differences between Type I and Type II fibres. Further subdivisions of these two types are recognised. The fibre‐ type is determined largely by the neuron innervating it. The constituent fibres of a motor unit are scattered throughout a muscle (‘mosaic’ not ‘random’). The properties of an anatomical muscle depend upon the proportions of I and II fibres. Enzyme activity mATPase activity (anaerobic) Oxidative capacity (SDH, aerobic) Speed of contraction Type I low high slow Type II high low fast Table Properties of Types I and II Muscle Fibres a) Type I fibres have a relatively high activity of mitochondrial oxidative enzymes (e.g. succinic dehydrogenase, SDH) and lower activity of phosphorylytic enzymes (especially myofibrillar ATPases). The velocity of contraction is relatively slow (for skeletal muscle); the resistance to fatigue is high. b) Type II fibres in contrast have a relatively high activity of ATPases and lower activities of oxidative enzymes. The velocity of contraction is very high and resistance to fatigue is relatively poor. 7. Muscle‐Fibre Type Grouping Nerve damage may lead to loss of innervation to individual muscle fibres which will be re‐innervated by adjacent nerve fibres (‘collateral re‐innervation’). This will lead to an increase in motor‐unit size resulting in loss of fine control. Conversion of the histochemical fibre‐types will lead to fibre‐type grouping. 8. Self Test Questions 1. 2. 3. Explain the following histological observation made in wax‐embedded, H&E‐stained sections. a. In smooth muscle cut in transverse section, not all profiles contain nuclei. b. White adipose tissue shows no sign of fat in a wax‐embedded section What do the blue granules in conventional sections of neurones represent? Define a motor unit. I 14