Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
History of RNA biology wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
Ridge (biology) wikipedia , lookup
Chloroplast DNA wikipedia , lookup
DNA vaccination wikipedia , lookup
Genomic library wikipedia , lookup
Transposable element wikipedia , lookup
Epigenetics of neurodegenerative diseases wikipedia , lookup
RNA interference wikipedia , lookup
RNA silencing wikipedia , lookup
Genomic imprinting wikipedia , lookup
Extrachromosomal DNA wikipedia , lookup
Cancer epigenetics wikipedia , lookup
Human genome wikipedia , lookup
Point mutation wikipedia , lookup
Genome (book) wikipedia , lookup
Short interspersed nuclear elements (SINEs) wikipedia , lookup
Epigenetics of diabetes Type 2 wikipedia , lookup
Genetic engineering wikipedia , lookup
Designer baby wikipedia , lookup
Non-coding RNA wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
No-SCAR (Scarless Cas9 Assisted Recombineering) Genome Editing wikipedia , lookup
Gene expression programming wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Long non-coding RNA wikipedia , lookup
Minimal genome wikipedia , lookup
Non-coding DNA wikipedia , lookup
Microevolution wikipedia , lookup
Helitron (biology) wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Genome editing wikipedia , lookup
Primary transcript wikipedia , lookup
Gene expression profiling wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Genome evolution wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Mir-92 microRNA precursor family wikipedia , lookup
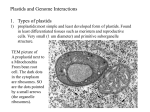
157 Protein expression in plastids Peter B Heifetz* and Ann Marie Tuttle† Current Opinion in Plant Biology 2001, 4:157–161 including roots, contain leucoplasts. The different plastids are further distinguished by their varying capacities for the expression of organelle-encoded genes. Illuminated chloroplasts, in particular, possess extraordinarily high rates of transcription and translation. These facilitate the accumulation of large amounts of soluble ribulose bisphosphate carboxylase (thus insuring that enzyme levels do not limit photosynthetic carbon fixation) and allow the rapid turnover of redox-sensitive electron transfer components, such as the D1 protein of photosystem II [4]. This feature makes leaf chloroplasts suitable for the production of large amounts of recombinant proteins. In this review, we summarize recent progress in understanding plastid protein expression and in transformation technology. 1369-5266/01/$ — see front matter © 2001 Elsevier Science Ltd. All rights reserved. Plastid transformation The genome of the plastid has generated much interest as a target for plant transformation. The characteristics of plastid transgenes both reflect the prokaryotic origin of plastid organelles and provide a unique set of features that are currently lacking in genes introduced into the plant nucleus. Recent progress has been made in understanding plastid expression of recombinant proteins. Addresses *Torrey Mesa Research Institute, 3115 Merryfield Row, San Diego, California 92121, USA; e-mail: [email protected] † Syngenta Agribusiness Biotechnology Research Inc, 3054 Cornwallis Road, Research Triangle Park, North Carolina 27709, USA Abbreviations aadA adenosyl-3′-adenyltransferase EPSPS 5-enolpyruvylshikimate-3-phosphate synthase GFP green fluorescent protein NEP nuclear-encoded RNA polymerase PEG polyethylene glycol PEP plastid-encoded RNA polymerase Introduction One of the unique and defining characteristics of plants is the presence of plastid organelles. These endosymbiotic remnants of a once free-living cyanobacterial progenitor [1] have, over evolutionary time, given up the vast majority of their genes and cellular functions to become the energy transduction and metabolic centers of the plant cell. Among the primitive features of the cyanobacterial progenitor that are retained in plastids is a circular and largely prokaryotic chromosome. This genome of circa 50–290 kilobases is remarkably similar across the algal and plant lineages with regard to the complement of genes encoded within it, their relative order and their sequence [2]. The plastid genome is also distinguished by its copy number, which even in the simplest of plants far exceeds the ploidy of the nuclear chromosomes. In a mature leaf cell, in which there may be as many as 100 plastids each containing upwards of 10–100 genomes, it is possible to find 10,000 or more identical copies of each plastid gene [3]. Plastids are distributed ubiquitously throughout the differentiated cells of diverse plant organs and tissues. Although each plant cell type contains identical copies of the plastid genome because of their organelles’ shared biogenesis from undifferentiated proplastids, the organelles themselves can vary tremendously in morphology and function. Thus, leaves and green tissues contain photosynthetic chloroplasts; mature fruits and flowers contain pigmented chromoplasts; tubers and other storage organs contain amyloplasts or elaioplasts; and other non-green tissues, Boynton and colleagues [5,6] demonstrated that all copies of the plastid genome in the single large chloroplast of the green alga Chlamydomonas reinhardtii could, following delivery of cloned DNA by particle bombardment, integrate such DNA in a targeted manner via homologous recombination. These pioneering studies made possible subsequent efforts to transform the plastids of higher plants [7]. They also laid out the unique ‘ground rules’ for plastid transformation as compared to nuclear transformation: foreign DNA must be delivered across the double membrane of the plastid envelope and across the cell membrane (and across the cell wall, if present); homologous (legitimate) recombination is the normal mode of DNA integration into the plastid genome. When suitable regions of homology between the introduced DNA and the target are provided, appropriate selectable markers for plastid transformation must facilitate the segregation of transformed genome copies at the expense of nontransformed ones; complete segregation of all the genome copies within a plastid, and within all of the plastids of a cell or tissue, must occur during clonal propagation in order to confer a stably transformed phenotype; and finally, bona fide plastid transformation events must be readily distinguishable at the molecular level from spontaneous mutations or ectopic nuclear integrations of the transforming DNA. In addition to particle bombardment, polyethylene glycol (PEG) treatment of protoplasts [8,9] has been found to be suitable for the delivery of DNA to plastids for stable transformation. This method appears more prone to creating unexpected nuclear mutations than is particle bombardment, however, as demonstrated by the relatively high level of selection escapes [10] and physiological artifacts [11] observed in PEG-transformed tobacco. Recently, an alternative means of directly delivering DNA to plastids was developed by Knoblauch et al. [12]. These workers used a sub-micron-diameter syringe 158 Plant biotechnology driven by the controlled thermal expansion of a liquid metal alloy to microinject DNA into chloroplasts. Although this method has not yet been shown to yield regenerated and fertile transformed plants, transient expression of the green fluorescent protein (GFP) transgene was achieved in tobacco without apparent harm to the injected cells. The ease with which plastid DNA can undergo homologous recombination between even short regions of sequence identity [13] has significant implications for the engineering of gene expression within plastids. First, targeted integration is the norm rather than the exception. This means that the location of transgenes can be predicted with precision so as to minimize interference with endogenous plastid genes and to prevent regulatory sequences near the integration site from influencing transgene expression. An added benefit is that comparatively few transgenic events need to be characterized as, in the absence of illegitimate recombination, only single-locus insertion events occur. (An exception to this norm results when the transforming DNA has been targeted to the inverted-repeat region of the plastid genome. Dual integration then occurs due to homologous copy correction between the two repeat regions.) Second, homologous recombination occurring spontaneously between directly repeated sequences flanking an integrated selectable marker gene can result in the excision of the marker gene from the genome when selective conditions are relaxed [14]. This approach has recently been shown to be feasible in plastids of higher plants [15••], raising the possibilities of sequential transformation by marker recycling and of elimination of bacterial antibiotic resistance genes from the final transgenic plants. A recent advance in selectable marker technology for plastids was the discovery that GFP can be stably transformed and expressed in the leaf chloroplasts of tobacco [16,17••] and potato [18••]. In potato, GFP was also expressed in tuber amyloplasts, albeit at a 100-fold lower level than in chloroplasts. Transient expression of GFP has been achieved in carrot, marigold and pepper [12], demonstrating that DNA can be delivered to plastids in these plant species. An even more versatile use of GFP was developed by Khan and Maliga [17••] who fused its coding sequence to that of the aadA gene, which encodes adenosyl-3′adenyltransferase. The resulting fusion molecule was functional both as an antibiotic-resistance gene and as a fluorescent protein in multiple types of plastids in tobacco. Moreover, following its introduction by bombardment and selection using streptomycin, activity of this fusion molecule was detectable at low levels in the plastids of rice suspension cells. These rice cells proved to be highly heteroplasmic, however, with only a small fraction of the plastid genomes transformed and no plants regenerated. Nonetheless, this remains the only successful demonstration of the stable delivery of foreign DNA to the plastids of a cereal species. Controlling plastid gene expression The plastid rpoA, rpoB, rpoC1 and rpoC2 genes encode the catalytic subunits of a eubacterial-type RNA polymerase that recognizes upstream sequences that have high homology to the consensus –10 and –35 regions typically found in eubacterial promoters. Promoter recognition by this plastid-encoded RNA polymerase (PEP) is mediated by sigma-like factors that are encoded in the plant nucleus. These factors are expressed in a regulated manner in response to environmental or developmental cues and are imported post-translationally into plastids via transit peptides [19]. Interestingly, plastids contain a second (and in spinach, likely a third) complete transcriptional apparatus that is entirely nuclear encoded. This apparatus contains a single-subunit that is phylogenetically related to the RNA polymerases of fungal and plant mitochondria, and of bacteriophages such as T7 and T3 [20–22]. The promoter elements recognized by the nuclear-encoded RNA polymerase (NEP) bear little similarity to eubacterial or PEP promoters [23] and require one or more specificity factors for their correct interaction with the polymerase [24]. Plastid genes can have only PEP promoters, only NEP promoters, or hybrid promoter regions that contain both PEP and NEP elements. The significance for message accumulation of these multiple transcription initiation sites is unclear. A recent study suggests, however, that the NEP may recognize DNA promiscuously and, thus, could be capable of transcribing any plastid gene [25]. Nonetheless, PEP and NEP elements are each capable of directing the expression of foreign genes in plastids [26,27]. Trans-activation expression systems using completely heterologous RNA polymerases, such as those from T7 phage, have also been developed for plastids [28,29•]. This approach allows the efficient and controlled transcription of target plastid transgenes as long as appropriate promoters, which are recognized by the T7 polymerase but not by NEP or PEP, are located upstream. The primary advantage of trans-activation is that it allows the imposition of developmental, tissue-specific, or chemically-inducible regulation upon the expression of plastid transgenes through control of the polymerase by nuclear promoters of the desired specificity. Using the tobacco PR-1a promoter [30] to direct the expression of a plastid-targeted T7 polymerase, β-glucuronidase (GUS) and a cellulose-degrading enzyme were expressed at high levels in tobacco chloroplasts in response to foliar application of an inexpensive fieldregistered compound [29•]. The production of primary transcripts is not typically the limiting step for the expression of plastid genes, although this step does limit plant nuclear expression and, to some degree, eubacterial expression. Instead, the rate of protein synthesis in plastids is much more dependent on posttranscriptional processes, which include transcript processing and stability, the conversion (by post-transcription RNA-editing mechanisms) of cytidines to uridines in certain Protein expression in plastids Heifetz and Tuttle editing-dependent plastid messages, and translational initiation or elongation on polyribosomes. These events are mediated at the RNA level by the binding of nuclearencoded factors to the 5′ untranslated leader and 3′ untranslated trailer regions of the plastid messages [31] and, in the case of RNA editing, by the site-specific interaction of an editing complex with RNA segments flanking the targeted nucleotides [32,33]. The efficient expression of transgenes in plastids thus requires not only the use of appropriate promoters but also the presence of the correct sequences in the 5′ and 3′ untranslated regions. The occurrence of RNA editing also necessitates care in the choice of the cloned sequences flanking a plastid expression cassette that are designed to direct homologous integration into the plastid genome. Editing sites, but not editing itself, may be conserved among related plant species [34]. Hence, it is important to prevent the inadvertent introduction of editing-dependent sequences into a genetic background that lacks the capacity for correct processing when flanking sequences from one plant species are used to direct integration into the plastids of a second plant species [34]. Uniparental inheritance of plastid genes Another unusual feature of plastid genes is their nonMendelian mode of inheritance. In gymnosperm plants, plastid DNA is maintained in sperm cells but not egg cells and is therefore transmitted uniparentally by the male. In the majority of angiosperms (including most of the important crops with the exception of alfalfa and to a limited extent potato), however, it is the pollen that loses the plastid DNA after mitosis I. Consequently, pollen cannot transmit the contents of the plastid genome to the zygote [35]. This could, in theory, prevent the spread of plastid transgenes through pollen to neighboring crops or related wild species [36•]. Recombinant protein expression in plastids The plastid genome is an attractive location for the engineering of pest-resistance and herbicide-tolerance traits, which typically benefit from high and stable levels of gene expression. Insecticidal Bt proteins derived from Bacillus thuringienesis cry genes are particularly amenable to plastid expression in tobacco. McBride et al. [37] and, more recently, Kota et al. [38•] demonstrated that plastid transformation with native bacterial cry sequences, produces extraordinarily high levels of protein accumulation as compared to nuclear transformation with codon-optimized transgenes. The resulting leaf tissue is extremely toxic to target insect larvae. In a recent follow-up study [39••], it was found that the introduction of the complete Bt cry2Aa2 operon into tobacco plastids caused even higher levels of expression than did the single genes. This was accompanied by the formation of cuboidal Bt protein crystals. This work is the first to demonstrate the expression of a natural bacterial operon in plastids and follows on from the initial report by Staub and Maliga [40] of chimeric operon expression from the tobacco plastid genome. Daniell et al. 159 [41] demonstrated that expression of a wild-type 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene and its complete transit-peptide sequence from petunia in the plastids of tobacco resulted in a ten-fold-greater level of tolerance to the herbicide glyphosate. The expression of additional herbicide-tolerance genes in plastids, including mutant EPSPS, protoporphyrinogen IX oxidase, and several bacterial genes, has been described recently in the patent literature [29•]. One of the most exciting potential applications of plastid protein expression is in the production of recombinant proteins for industrial, pharmaceutical or other valueadded uses. The biological containment afforded by plastid localization is attractive from the standpoint of reducing the risks of the pollen transmission of genes that may encode metabolically active proteins. Moreover, the high levels of expression that have been reported for plastid-encoded proteins could effect substantial cost savings in cases in which large amounts of tissue must be processed in order to yield purified molecules. Plastids are also capable of disulfide-bond formation and hydrolysis via their endogenous thioredoxin and protein disulfide isomerase enzyme systems. Mammalian proteins, such as the growth hormone somatotropin, normally require passage through the endoplasmic reticulum in order to attain their mature disulfide-bonded conformation. Recently, Staub et al. [42••] demonstrated that metabolically active human somatotropin could be expressed and correctly processed in the plastids of tobacco where it accumulated to 7% of total soluble protein. In contrast to this successful expression of a mammalian protein, attempts by Guda et al. [43] to express a protein biopolymer derived from mammalian elastin were disappointing. These authors found that although the synthetic biopolymer gene consisting of multiple repeating units of the amino-acid sequence GVGVP was transcribed effectively in tobacco plastids, the accumulated message was either not readily translatable by the plastid ribosomes or was rapidly turned over. Consequently, the levels of polymer product detected in plastids were low and thus highly inefficient compared to expression of the same recombinant protein in the plant nucleus. Conclusions and future prospects Plastid biotechnology has now progressed to the point at which a range of proteins from prokaryotic and eukaryotic sources have been expressed effectively in leaf chloroplasts and other plastids of tobacco. Progress is still needed in the improvement of transformation technology (to expand the range of plant species that can be efficiently transformed) and in the control of heterologous gene expression in plastids at the post-transcriptional level. Overcoming these challenges will result in a means of producing high levels of recombinant proteins or pathway-derived metabolites in a stable and predictable manner with the potential for robust physical and genetic containment. 160 Plant biotechnology Acknowledgements The authors thank Eric Boudreau for a critical reading of the manuscript. References and recommended reading Papers of particular interest, published within the annual period of review, have been highlighted as: • of special interest •• of outstanding interest resistant to high levels of this antibiotic because of naturally occurring mutations in their 16S ribosomal RNA. 18. Sidorov VA, Kasten D, Pang SZ, Hajdukiewicz PT, Staub JM, •• Nehra NS: Stable chloroplast transformation in potato: use of green fluorescent protein as a plastid marker. Plant J 1999, 19:209-216. The authors describe both the first published use of GFP as a plastid marker and stable plastid transformation in a food crop. Their results show a greater than 100-fold difference in the expression of protein in chloroplasts and amyloplasts. 19. Allison LA: The role of sigma factors in plastid transcription. Biochimie 2000, 82:537-548. 1. Moreira D, Le Guyader H, Philippe H: The origin of red algae and the evolution of chloroplasts. Nature 2000, 405:69-72. 2. Gray MW: Evolution of organellar genomes. Curr Opin Genet Dev 1999, 9:678-687. 3. Bendich AJ: Why do chloroplasts and mitochondria contain so many copies of their genome? BioEssays 1987, 6:279-282. 4. Heifetz PB, Lers A, Turpin DH, Gillham NW, Boynton JE, Osmond CB: dr and spr/sr mutants of Chlamydomonas reinhardtii affecting D1 protein function and synthesis define two independent steps leading to chronic photoinhibition and confer differential fitness. Plant Cell Environ 1997, 20:1145-1157. 23. Hajdukiewicz PTJ, Allison LA, Maliga P: The two RNA polymerases encoded by the nuclear and the plastid compartments transcribe distinct groups of genes in tobacco plastids. EMBO J 1997, 16:4041-4048. 5. Boynton JE, Gillham NW, Harris EH, Hosler JP, Johnson AM, Jones AR, Randolph-Anderson BL, Robertson D, Klein TM, Shark KB et al.: Chloroplast transformation in Chlamydomonas with high velocity microprojectiles. Science 1988, 240:1534-1538. 24. Bligny M, Courtois F, Thaminy S, Chang C-C, Lagrange T, Baruah-Wolff J, Stern D, Lerbs-Mache S: Regulation of plastid rDNA transcription by interaction of CDF2 with two different RNA polymerases. EMBO J 2000, 19:1851-1860. 6. Newman SM, Boynton JE, Gillham NW, Randolph-Anderson BL, Johnson AM, Harris EH: Transformation of chloroplast ribosomal RNA genes in Chlamydomonas: molecular and genetic characterization of integration events. Genetics 1990, 126:875-888. 7. Svab Z, Hajdukiewicz P, Maliga P: Stable transformation of plastids in higher plants. Proc Natl Acad Sci USA 1990, 87:8526-8530. 25. Krause K, Maier RM, Kofer W, Krupkinska K, Herrmann RG: Disruption of plastid-encoded RNA polymerase genes in tobacco: expression of only a distinct set of genes is not based on selective transcription of the plastid chromosome. Mol Gen Genet 2000, 263:1022-1030. 8. Golds T, Maliga P, Koop H-U: Stable plastid transformation in PEG-treated protoplasts of Nicotiana tabacum. Biotechnology 1993, 11:95-97. 9. O’Neill C, Horvath GV, Horvath E, Dix PJ, Medgyesy P: Chloroplast transformation in plants: polyethylene glycol (PEG) treatment of protoplasts is an alternative to biolistic delivery systems. Plant J 1993, 3:729-738. 20. Gray MW, Lang BF: Transcription in chloroplasts and mitochondria: a tale of two polymerases. Trends Microbiol 1998, 6:1-3. 21. Hess WR, Borner T: Organellar RNA polymerases of higher plants. Int Rev Cytol 1999, 190:1-59. 22. Lerbs-Mache S: Regulation of rDNA transcription in plastids of higher plants. Biochimie 2000, 82:525-535. 26. Sriraman P, Silhavy D, Maliga P: The phage-type PclpP-53 plastid promoter comprises sequences downstream of the transcription initiation site. Nucleic Acids Res 1998, 26:4874-4879. 27. Eibl C, Zou Z, Beck A, Kim M, Mullet J, Koop HU: In vivo analysis of plastid psbA, rbcL and rpl32 UTR elements by chloroplast transformation: tobacco plastid gene expression is controlled by modulation of transcript levels and translation efficiency. Plant J 1999, 19:333-345. 10. Koop HU, Steinmuller K, Wagner H, Rossler C, Eibl C, Sacher L: Integration of foreign sequences into the tobacco plastome via polyethylene glycol-mediated protoplast transformation. Planta 1996, 199:193-201. 28. McBride KE, Schaaf DJ, Daley M, Stalker DM: Controlled expression of plastid transgenes in plants based on a nuclear DNA-encoded and plastid-targeted T7 RNA polymerase. Proc Natl Acad Sci USA 1994, 91:7301-7305. … chlororespiration: only half a story. 11. Nixon PJ, Maliga P: Reply… Trends Plant Sci 1999, 4:51. 29. Heifetz PB: Genetic engineering of the chloroplast. Biochimie • 2000, 82:655-666. This paper includes a description of a chemically inducible transactivation system for the expression of genes from the plastid genome. Such an approach is important when heterologous gene products are deleterious to the plant because of either their high expression levels or their biological activity. The use of induction in mature plants allows the expression of otherwise toxic molecules as transformation is performed in the absence of expression, which is only enabled once the nuclear transactivator and plastid target are brought together in a sexual cross. 12. Knoblauch M, Hibberd JM, Gray JC, van Bel AJ: A galinstan expansion femtosyringe for microinjection of eukaryotic organelles and prokaryotes. Nat Biotechnol 1999, 17:906-909. 13. Kavanagh TA, Thanh ND, Lao NT, McGrath N, Peter SO, Horvath EM, Dix PJ, Medgyesy P: Homeologous plastid DNA transformation in tobacco is mediated by multiple recombination events. Genetics 1999, 152:1111-1122. 14. Fischer N, Stampacchia O, Redding K, Rochaix JD: Selectable marker recycling in the chloroplast. Mol Gen Genet 1996, 251:373-380. 15. Lamtham S, Day A: Removal of antibiotic resistance genes from •• transgenic tobacco plastids. Nat Biotechnol 2000, 18:1172-1176. The work described in this paper and [14] demonstrates the feasibility of excising selectable marker genes or other sequences from the plastid genomes of stably transformed plants. 16. Gray JC, Hibberd JM, Linley PJ, Uijtewaal B: GFP movement between chloroplasts. Nat Biotechnol 1999, 17:1146. 17. •• Khan MS, Maliga P: Fluorescent antibiotic resistance marker for tracking plastid transformation in higher plants. Nat Biotechnol 1999, 17:910-915. A technical advance resulting in the first stable integration of foreign DNA into the plastid genome of a monocot plant. The work uses an approach in which the initial selection of cultured cells on streptomycin is followed by screening for green fluorescence. The aadA gene product catalyzes the covalent modification of both streptomycin and spectinomycin. Although the latter is a more effective plastid-selectable marker, most cereals are naturally 30. Uknes S, Dincher S, Friedrich L, Negrotto D, Williams S, Thompson-Taylor H, Potter S, Ward E, Ryals J: Regulation of pathogenesis-related protein-1a gene expression in tobacco. Plant Cell 1993, 5:159-169. 31. Barkan A, Goldschmidt-Clermont M: Participation of nuclear genes in chloroplast gene expression. Biochimie 2000, 82:559-572. 32. Chaudhuri S, Carrer H, Maliga P: Site-specific factor involved in the editing of the psbL mRNA in tobacco plastids. EMBO J 1995, 14:2951-2957. 33. Bock R, Koop U: Extraplastidic site-specific factors mediate RNA editing in chloroplasts. EMBO J 1997, 16:3282-3288. 34. Corneille S, Lutz K, Maliga P: Conservation of RNA editing between rice and maize plastids: are most editing events dispensable? Mol Gen Genet 2000, 264:419-424. 35. Nagata N, Saito C, Sakai A, Kuroiwa H, Kuroiwa T: The selective increase or decrease of organellar DNA in generative cells just after pollen mitosis one controls cytoplasmic inheritance. Planta 1999, 209:53-65. Protein expression in plastids Heifetz and Tuttle 36. Scott SE, Wilkinson MJ: Low probability of chloroplast movement • from oilseed rape (Brassica napus) into wild Brassica rapa. Nat Biotechnol 1999, 17:390-392. A theoretical study comparing the effect of nuclear and plastid transformation on transgene flow in a readily out-crossed plant species. Plastid expression blocks pollen transmission but still allows the production of weedy hybrids if the female parent is pollinated by a related species. 37. McBride KE, Svab Z, Schaaf DJ, Hogan PS, Stalker DM, Maliga P: Amplification of a chimeric Bacillus gene in chloroplasts leads to an extraordinary level of an insecticidal protein in tobacco. Biotechnology 1995, 13:362-365. 38. Kota M, Daniell H, Varma S, Garczynski SF, Gould F, Moar WJ: • Overexpression of the Bacillus thuringiensis (Bt) Cry2Aa2 protein in chloroplasts confers resistance to plants against susceptible and Bt-resistant insects. Proc Natl Acad Sci USA 1999, 96:1840-1845. In a similar study to the pioneering work described in [37], a single bacterial gene encoding a Bt toxin was expressed in tobacco plastids and found to express at high levels, causing considerable mortality to target insect species. The use of highly insect-resistant plants as part of a ‘high dose resistance management strategy’ is discussed. 39. De Cosa B, Moar W, Lee S-B, Miller M, Daniell H: Overexpression of •• the Bt cry2Aa2 operon in chloroplasts leads to formation of insecticidal crystals. Nat Biotechnol 2001, 19:71-74. Insecticidal proteins from Bacillus thuringiensis are normally expressed in the insect as protoxins from operons, which can include a putative chaperonin. 161 These protoxins form crystalline inclusion bodies in the natural host, which increase their stability. The authors achieved the expression of one such complete operon, which was encoded by three genes, in tobacco plastids. This expression resulted in the production of cuboidal crystals within leaf chloroplasts. The resulting proteins were immensely toxic even to recalcitrant pests such as beet armyworm and cotton bollworm. 40. Staub JM, Maliga P: Expression of a chimeric uidA gene indicates that polycistronic mRNAs are efficiently translated in tobacco plastids. Plant J 1995, 7:845-848. 41. Daniell H, Datta R, Varma S, Gray S, Lee SB: Containment of herbicide resistance through genetic engineering of the chloroplast genome. Nat Biotechnol 1998, 16:345-348. 42. Staub JM, Garcia B, Graves J, Hajdukiewicz PTJ, Hunter P, Nehra N, •• Paradkar V, Schlittler M, Carroll JA, Spatola L et al.: High-yield production of a human therapeutic protein in tobacco chloroplasts. Nat Biotechnol 2000, 18:333-338. Proteins from eukaryotic sources, which require complex folding or disulfide bond formation, are generally not easily expressed in prokaryotic systems. The authors demonstrate that the tobacco chloroplast is capable of correctly processing fusions of human somatotropin and ubiquitin expressed from the plastid genome. Moreover, these transformed chloroplasts accumulate metabolically active protein to high levels. 43. Guda C, Lee SB, Daniell H: Stable expression of a biodegradable protein-based polymer in tobacco chloroplasts. Plant Cell Rep 2000, 19:257-262.