Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Eyeblink conditioning wikipedia , lookup
Limbic system wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Neuroregeneration wikipedia , lookup
Microneurography wikipedia , lookup
Apical dendrite wikipedia , lookup
Synaptogenesis wikipedia , lookup
Development of the nervous system wikipedia , lookup
Axon guidance wikipedia , lookup
Anatomy of the cerebellum wikipedia , lookup
Hypothalamus wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Circumventricular organs wikipedia , lookup
Subventricular zone wikipedia , lookup
Neuroanatomy wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Sensory cue wikipedia , lookup
Optogenetics wikipedia , lookup
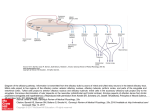
The Hunan Nervous System: An Ahatonical Viewoint, Mlnay L. Bar and John A. Kieman. J.B, Lippincott Company, Philadelphia, O 1993. Seventeen Sixth Edition, e The olfactory receptor cells are unique in being neurons located in an epithelium and in being regularly replaced from a population of precursor cells. The unmyelinated axons of the olfactory neurosensory cells constitute about 20 olfactory nerves on each side. These nerves pass through the cribriform plate o[the ethmoid bone and end in the overlying olfactory bulb. A fracture of the cribriform plate is likely to be followed by cerebrospinal fluid rhinorrhea. The principal neurons of the olfactory bulb have axons that form the olfactory tract. This follows the ventral surface of the frontal lobe and ends in the olfactory trigone, anterior (rostral) to the anterior perforated substance. Most of the axons of the olfactory tract follow the lateral olfactory stria and end in the lateral olfactory area, which comprises the corte>< of the uncus, the limen insulae, the entorhinal area, and the corticomedial nuclei of the amygdaloid body. Smaller numbers of olfactory tract fibers end in the anterior olfactory nucleus and in various nuclei in the region of the anterior perforated substance. Some of these cell groups give rise to fibers that pass centrifugally in the olfactory tracts and terminate in the olfactory bulbs of both sides, providing a mechanism for modulation of the input from the olfactory apparatus. The regions in which fibers of the olfactory tract terminate are connected, directly and indirectly, with the limbic system, hypothalamus, and reticular formation of the brain stem. These connections provide for visceral and behavioral responses to different odors. The olfactory system consists of the olfactory epithelium, olfactory nerves, olfactory bulbs, and olfactory tracts, together with functionally associated cerebral cortex and subcortical structures. Lower vertebrates and many mammals relyheavily onthe sense of smell. They are said to be macrosmatic; in the mammalian class, the dog is a familiar example. Humans are microsmatic, with smell being much less important than the other senses, especially sight and hearing. The study of comparative anatomy contributes much to an undetstanding of those parts of the brain involved in olfaction, which constitute the rhinencephalon, Thus, in macrosmatic animals, the rhinencephalic 269 i _. . l,:. i.. 27(l Regional Anatomy of the Central Nervous System in humans, they are small by comparison with the remainder of the brain. Even in humans, however, olfaction is a significant sense that structures are large and prominent, whereas conjures up memories and arouses emotions, Smell also contributes to alimentary pleasures. Those who have lost their sense of smell complain of impairment of taste, stating that everything is bland and tastes alike, and they may be unaware of their inability to smell. Much of our enjoyment of taste is in fact an appreciation of aromas through the olfactory system. Some chemical stimuli, notably those from foods with "hot" flavors, excite general sensory fibers of the trigeminal nerve in the nose and mouth. The olfactory, gustatory, and general sensory responses to chemical stimuli in the nose may be integrated in the insula, where the primary cortical areas for the three systems are in proximity. Olfactory Epithelium and Olfactory Nerves The olfactory epithelium is derived from an ectodermal thickening, the olfactory placode, at the rostral end of the embryonic head, The cells of this placode give rise to the cells of the epithelium, the glial cells of the olfactory nerves, and probably some of the glial cells of the most superficial layer of the olfactory bulb. In the adult, the olfactory ep]thelium (Fig. 17- I ) covers an area of 2.5 cnit" in the roof of each nasal cavity and extends for a short dis- tance on the lateral wall of the cavity and the nasal septum. The olfactory sensory cells are contained in a pseudostratified columnar epithelium, which is thicker than that lining the respiratory passages elsewhere, Olfactory glands (glands of Bowman) beneath the epithelium bathe the surface with a layer of Olfactory vesicles with cilia Supporting CCIIS Oltactory neu rosensory cell (position of nucleus) lntermediate stage between basal cell and olfactory neu rosetlsory cel Basal Olf actory IBowman's) ceils grano 1t metnbrane Figure 17-1. Olfactory epithelium. Filum of olfactory nerve in lamina propfla I Chapter 17: Olfactory System mucous fluid, inwhich odoriferous substances are dissolved. The olfactory neurosensory cells (also known as primary olfactory neu_ rons or simply as olfactory cells) are bipolar neurons that serve as sensory receptors as well as conductors of impulses. The major mod_ ification consists of specialization of the dendrite; this process extends to the surface of the epithelium, where it ends as an exposed bulbous enlargement known as an olfictory vesicle, bearing cilia that are exceptional in that they may be up to 100 pm long. R minority of human.neurosensory cells have apical tufts of microvilli and resemble the sensory neurons of the vomeronasal organ, a chemical sense organ of lower mammals and submam_ malian vertebrates (see end pf this chapter) . Unmyelinated axons of the olfactory cells are gathered into about 20 bundles on each side, which are the olfactory nerves. These enter the cranial cavity by passing through the foramina of the cribriformplate of the ethmoid bone and then enter the olfactory bulb. The axons form a superficial fibrous layer in the olfactory bulb, then continue more deeply, and termirLate in specialized synaptic conligurations, the glomeruli. The olfactory axon terminals release an excitatory neurotransmitter that has not yet been identified. (The dipeptide carnosine has been suspected.) The few neurosensory cells shown in Figure l7-1 represent some 25 million such cells in each half of the olfactory epithelium. The olfactory cells are continuously being produced by uritosis and differentiation of some of the basal cells of the olfactory epithelium, and lost by desquamation. Observations in animals indicate that although some cells die without reaching maturity, olfactory neurons probably are lost by wear and tear rather than because of an innately short life span, In the hurrran nose, each receptor neuron survives probably for about 2 months, Consequently there are always new axons growing along the olfactory nerves and into the olfactory bulbs. The olfactory system is exquisitely sensitive to minute amounts of excitants in the air. Direct stimulation of the receptors, convergence of many neurosensory cells on the principal neurons of the olfactory bulb, and facilitation by neuronal circuits in the bulb are among the factors responsible for the low threshold, Smell is a chemical sense, as is taste. For a substance to be smelled, it must enter the nasal cavity as a gas or as an aerosol and then dissolve in the fluid that covers the olfactory epithelium. The secretory product of Bowman,s glands con_ tains glycoproteins that can bind odoriferous substances that are not otherwise soluble in water, for presentation to receptor molecules on the surfaces of the sensory cilia. That a large range of odors and aromas can be appreciated may be due in part to the existence of neu.o_ sensory cells with different chemical speci_ ficities. The olfactory system adapts rather quickly to a contrnuous stimulus, so that the odor becomes unnoticed. A physiological mechanism that allows the receptors to recover is a cyclic alternation of mucosal blood flow in the left and right sides of the nose. At any instant, the side with the higher flow of blood presents greater resistance to the flow of air because of swelling of the mucosa. The nasal cavity with lower air flow consequently receives smaller amounts of the ambient odoriferous sub_ stances. Most older people have a reduced acu_ ity of smell, probably caused by a progressive reduction in the population of neurosensory cells in the olfactory epithelium. Ollactory Bulb, Ttact, and Striae The olfactory bulb is ventral to the orbital sur_ face of the frontal lobe. It is connected by the olfactory tract to a central point of attachment in front of the anterior perforated substance (see Fig. 17-3). The olfactory bulb has a characteristic cvto_ architecture in animals that rely heavily on the sense of smell. There are five layers (Fig. Z-2): 1. Nerve fiber layer (olfactr:ry axons) on the surface 2. Layer of glomeruli 3. External plexiform layer 271 272 Regional Anatoffiy of the Central Nerttous Systeu Fibers'of Cranule cell (has no axon) Periglomerular cell Layer of mitral cells External plexiform layer Tufted cell Layer of glomeruli Cribriform plate cel I Figure l7-2. Neuronal circuitry of the olfactory bulb' 4. Layer of mitral cells 5. Granule cell layer, which in its deeper parts also contains the myelinated axons ihat comprise the medullary center of the olfactory bulb The center contains nests of ependymal cells, which are vestiges of the extension of the lateralventricle into the bulb in embryonic life' e '1 :l t .; . Chapter 17: Olfactory neurons with which they synapse, and the transmitter probably is glutamate. The olfactory bulb contains interneurons of two types. Periglomerular cells have dendrites that receive ctory neuroepithelial fibers of the sy cells olfactory tract drodendritic synapses with the mitral cells. The axons of the periglomerular cells enter the external plexiform layer to contact the dendrites of mitral cells associate e><cite or i recePtors mine, 1-a peptides. The most numerous interneurons are the GABAergic granule cells, which have no axons and are located in the deepest layer ofthe olfactory bulb. Their dendrites receive axo- dendritic contacts from mitral 'cells and from the centrifugal fibers. Other dendrites form dendrodgndritic synapses with mitral cell dendrites. Sorne of these synaptic arrangements are shown irr Figure 77 -2.The complex circuitry of the olfactory bulb recalls that of the retina and indicates thiat, as is the case with visual images, sensory data are partially analyzed and edited before reaching the cerebral olfactory areas. Three srnall groups of nerve cells make up the anterior olfactory nucleus. One is situated ar the transition between the olfactory bulb and olfactory tract; the others are deepr to the lateral and medial olfactory striae described in the next paragraph. Collateral branches of axons of mitral and tufted cells terminate in this nucleus. Fibers that originate in the anterior olfactory nucleus pass through the anterior commissure to the contralateral olfactory bulb. This is only one of the populations of centrifugal fibers that project to the olfactory bulb. Centrifugal fibers synapse principally with the dendrites of the interneurons. Impulses from the olfactory bulb are conveyed to olfactory areas for subjective appreciation of odors and aromas. These areas also establish connections with other parts of the brain for emotional and visceral responses to olfactory stimuli. The olfactory tract expands into the ollactory trigone at the rostral margin of the anterior perforated substance. Most of the axons of the tract pass into the lateral olfactory stria (Fig. 17-3), which goes to rhe System lateral olfactory area. Other axons of the olfactory tract, traditionally named the intermediate olfactory stria, leave the olfactory tdgone to enter the anterior perforated sub_ stance, which is part of the intermediate olfactory area. The name "medial olfactory stria,,is applied to a ridge that was once thought to carry olfactory fibers to the septal area. It is now known that no such connection exists. Olfactory Areas of the Cerebral Hemisphere R.HINENCEP LON The "nose brain" was once thought to include a much larger proportion of the forebrain than that now known to be devoted to the sense of smell. The term is now restricted to those re- gions that receive afferent fibers from the ol_ factory bulbs. The lateral olfactory area receives afferents from the olfactory bulb through the lateral olfacrory stria (Fig. 1z-+; see also Fig. 17-3). The area consists of the paleocortex of the uncus, cortex of the entorhinal area (the anterior part of the parahip- pocampal gyrus) in the temporal lobe, and cortex in the region of the limen insulae (see Fig. 17-3). The uncus, entorhinal area, and limen insulae are collectively known as the pyriforrn cortex (or lobe) because the homologous area has a pear-shaped outline in macrosmatic animals. part of the amygdaloid body (amygdala) also is included in the lateral olfactory area; the uncus is its landmark on the medial sudace of the temporal lobe. The dorsomedial part of the amygdala consists of the corticomedial group of nuclei, It receives olfactory fibers, whereas the larger ventro- lateral portion, a component of the limbic system, is considered in Chapter lg. The lateral olfactory area is the principal region for awareness of olfactory stimuli and is, therefore, the prirnary olfactory area. The anterior perforated substance, situated between the olfactory trigone and the optic tract (see Fig. 17 -3) , derives its name from the penetration of many small blood vessels into the brain in this region. It contains several 27t 274 Regional Anatorny of the Central Nenous System Olfactory bulb Olf actorY trigone Lateral "Medial olf actory olfactory stria" stfla Anterioi perforated substance Limen insulae Metal retractor Figure l7-3. Some components of the olfactory system seen on the , ventral surface of the brain. The right temporal Pole has been cut away to give a clear view of the olfactory trigone, anterior perforated substance, and limen insulae. (x 1) groups of neurons that receive fibers from the olfactory trigone and together constitute the intermediate olfactory area. The diago' nal band of Broca, immediately in front of the optic tract and beneath the gray matter, connects the ventrolateral portion of the amygdala with the septal area and is, therefore, a fiber bundle of the limbic system, The adjacent nucleus of the diagonal band, however, is a major source of centrifugal fibers to the olfactory bulb, the other source being the contralateral anterior olfactory nucleus. The septal area, on the medial surface of the frontal lobe ventral to the rostrum of the corpus callosum, was formerly known as the "medial olfactory area" , but it does not receive any fibers of the olfactory tract. The septal area is a component of the limbic system of the brain and can no longer be assigned a role in olfaction as well.l Olfactory stimuli induce visceral responses by modulating the activities of the autonomic nervous system. Examples are salivationwhen there are pleasing aromas from the prepara- t The nomenclature of the olfactory areas of the human brain is decidedly unsatisfactory. Even though a "medial" olfactory area is no longer recognized in the mammalian brain, we have retained the old "intermediate olfactory area" for want of a better term. A detailed account of its components is beyond the scope of the present text. i, Chapter 17: Oflactory System Sources of centrifugal fibers of olfactory tract: anterior olfactory nucleus and nucleus of diagonal band Intermediate Olfactory bulb olfactory area Olfactory tract Lateral olfactory areA Entorhinal area Olfactory epithelium Figure l7-4. Components of the olfactory tract. tion of food, and nausea or even vomiting evoked by an ofllensive stench, The olfactory system shares the entorhinal cortex with the limbic system, and the limbic system has extensive connections with the septal area and the hypothalamus. Most of the fibers that connect the septal area'and hypothalamus with Clinical Coraideraiions Fractures of the floor of the anterior fossa of the autonomic nuclei are in the medial forebrain bundle. This bundle, which contains fibers projecting rosftally as well as caudally, traverses the lateral part of the hypothalamus. Descending fibers from the hypothalamus proceed to autonomic nuclei in the brain stem and spinal cord. Other descending fibers of the medial forebrain bundle end in raphe reticular nuclei and in the solitary nucleus.' 2 The autonomic nuclei are associated with cranial nerves III, VII. IX, and X (see Ch. 8) and with spinal nerves TI-L2 and S2-S4 (see Ch. 5). The organization and functions ofthe autonomic nervous system are discussed in Chapter 24. cation with the external environment is dangerous because it provides a route whereby bacteria.may enter and attack the meninges and the brain. tory loss is likely to be unilateral. An irritating lesion that affects the lateral olfactory area may cauEe acterized by an imaginary involuntary movements o 275 276 Regional Anatomy of the Central Neruous System and often by other features ol'disturbed function of the temporal lobe (seer Ch. 1B). rminal and meronasal Nerves - TWo small cranial nerves associated with the olfactory system were discov,:red after the l2 main cranial nerves were given their numbers. The terminal nerve (nervus terminalis) is present, although of microscopic size, in the adult human brain. Sometimes it is called cranial nerve zero because it is medial (and therefore perhaps rostral) to the olfactory nerves. The vomeronasal system appears on-ly transiently in human embryonic developrnent, but in most other terrestrial vertebrates, it has important functions in adult life. Terminal Nerue The fibers of the tiny terminal nerve lie along the medial side of the olfactory irulb and olfactory tract. Bipolar neuronal cell bodies are present in small ganglia along the coutrse of the nerve. Their distal processes pass through the cribriform plate and are distributed to the nasal septum. In animals, the proximal procesSes have been traced experimentally to the septal and preoptic areas. Vomeronasal Sgstem In most mammals other than humans and in many submammalian vertebrates, there is a vomeronasal organ, which is a blind-ended tube lined by sensory epithelium in the ventral part of each side of the nasal septum. The re- teptor neurons are similar to those in the olfactory epithelium, but they have microvilli rather than cilia at their apical poles. Vascular connective tissue with sympathetic vasomotor innervation separates the vomeronasal epithelium from the rigid cartilaginous wall of the tube' The neurosensory cells of the epithelium give rise to axons that constitute the vomeronasal nerve. This passes through the cribriform plate alongside the olfactory neryes an,l ends in a part of the forebrain called the accessory olfactory bulb. In lower mammals arrd reptiles, the entrance to the vomeronasal organ is immediately dorsal to the nasopalatine foramen, through which the oral and nasal cavities communicate behind the incisor teeth. Activity of the sympathetic nervous system causes constriction of the blood vessels of the vomeronasal organ, making the connective tissue layer thinner because it contains less blood. The resulting enlargement of the lumen sucks in tiny drops of liquid that have been either sniffed into the nostrils or deposited by the tongue into the nasopalatine foramen. The vomeronasal system is for the detection of chemical messages from other members of the same species. The compounds (pheromones) used as sexual attractants and for marking territory may be secreted by specialized sweat or sebaceous glands, or they may be in the urine. ln animals with welldweloped vomeronasal systems, such as snakes and rodents, transection of the vomeronasal nerves impairs reproductive behavior but does not interfere with feedino. excreted SUGGESTED READING Doucette JR, Kiernan JA, Flumerfelt BA: The re- of olfactory glomeruli following transection of primary olfactory axons in the central or peripheral nervous system. J Anat innervation l)7:l-19, 1983 Doucette R: PNS-CNS Tlansitional zone of the first cranial nerve , J Comp Neurol 312 451-466, l99r Eccles R, Jawad MSM, Morris S: Olfactory and trigeminal thresholds and nasal resistance to airflow. Acta Otolaryngol (Stockh) 108:268- 273,1989 Graziadei PPC, I(arlan MS, Monti Graziadei GA, Bemstein JJ: Neurogenesis of sensory neurons in the primate olfactory system after section of the fila olfactoria. Brain Res 186:289-)00, r980 Halpern M: The organization and Iunction of the vomeronasal system. Annu Rev Neurosci l0: 325-362, 1987 Hinds JW, Hinds PL, McNelly NA: An autoradiographic study of the mouse olfactory epithelium: Evidence for long-lived receptors, Anat Rec 2lO:375-)83, 1984 Mackay-Sim A, ICttel W: On the life span of olfactory receptor neurons. Eur J Neurosci 3:209- 2t5, l99l Morrison EE, Costanzo RM: Morphology of olfac- Chapter 17: OlfactorY SYstem tory epithelium in humans and other verte2)t49-61' Ur"tes, Microsc Res and Technique 1992 The differential projections of S.ufu --- n, Witt"ns SS: mammals' J Comp Neurol *r. ottu.tory bulb in 16r'.3r-56, r975 bulb' In ShepStrepherd GM, Greer CA: Olfactory e t:D il !S t- ng he 1at irst 66, rnd to t8GA, 'ons nof ]00, 'the l0: ,uto- :tory ltors. olfac- 20eolfac- herd GM (ed): The S1'naptic organization of the Brain, 3rd ed, pp I33-169. New York' Oxford UniversitY Press, 1990 Wysocki CJ. Meredith M: The vomeronasal system' In Finger TE, Silver WL (eds): Neurobiology of Taste and Smell, pp I25-I50' NewYork' WileyInterscience, 1987 277