Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Epigenetics of diabetes Type 2 wikipedia , lookup
Bisulfite sequencing wikipedia , lookup
Zinc finger nuclease wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Genealogical DNA test wikipedia , lookup
Gel electrophoresis of nucleic acids wikipedia , lookup
DNA polymerase wikipedia , lookup
United Kingdom National DNA Database wikipedia , lookup
Microevolution wikipedia , lookup
DNA damage theory of aging wikipedia , lookup
Designer baby wikipedia , lookup
Transcription factor wikipedia , lookup
Epigenetics of neurodegenerative diseases wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Nutriepigenomics wikipedia , lookup
DNA vaccination wikipedia , lookup
Molecular cloning wikipedia , lookup
Cell-free fetal DNA wikipedia , lookup
Point mutation wikipedia , lookup
Cancer epigenetics wikipedia , lookup
Epigenetics wikipedia , lookup
Non-coding DNA wikipedia , lookup
Epigenetics in stem-cell differentiation wikipedia , lookup
Extrachromosomal DNA wikipedia , lookup
DNA supercoil wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Nucleic acid double helix wikipedia , lookup
Deoxyribozyme wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
History of genetic engineering wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Helitron (biology) wikipedia , lookup
Primary transcript wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Epigenetics in learning and memory wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Histone acetyltransferase wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
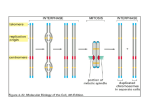
Chromatin Remodeling Levels of chromatin organization 300 nm fiber nucleosome arrays Structure of the nucleosome: problems of accessibility for DNAbinding proteins. Histone cores are predominantly alpha helical. Luger et al. (1997) Nature 389, 251. Analysis of binding of transcription factor to naked DNA and nucleosomal DNA • Assembling the DNA into a nucleosome strongly inhibits the binding of a sequencespecific transcription factor. • Assembling the DNA into a nucleosome leads to cleavage by DNase I at 10 nucleotide intervals. Transcription factor footprint Taylor et al (1991) Gene & Dev 5, 1285. DNase Footprinting DNase I binds the minor groove and cuts the phosphodiester backbone. When DNA rests against a surface, the minor groove is maximally accessible at ~10 base intervals. Analysis of chromatin changes by micrococcal nuclease transcription - + DNA Disappearance of ordered nucleosomes upon transcriptional induction Li & Reese(2001) JBC 276, 33788. “Chromatin remodeling complexes” and “Chromatin modifying complexes” are important for transcriptional activation Chromatin modifying complex Chromatin remodeling complex Examples of histone modification Berger (2002) Curr. Opin. Gene. Dev. 12, 142. The “histone code” hypothesis : the pattern of post-translational modifications occurring on the histone tails serves as binding sites for specific proteins. • Note that other chromatin modifying complexes include kinases, methylases and ubiquitin conjugating proteins. • Acetylation typically correlates with transcriptional activation while deacetylation correlates with repression. Marmorstein (2001) Nat. Rev. Mol. Cell. Biol. 2, 422. Histone Acetyl Transferases • Multiple families • Gene-specific or global activators of transcription • Distinct substrate specificities for different families • Could acetylate non-histone proteins (transcription factors) How acetylation might contribute to activation • Weakens interaction of basic tails with negatively charged phosphate backbone of DNA. • Weakens interactions that occur between nucleosomes, thus promoting decondensation of the chromatin fiber. • Provide a marker for recognition by other proteins. For example, a conserved “bromo” domain found in SWI/SNF and other transcription factors recognizes this marker. Non-enzymatic domains in Chromatin Modification proteins Marmorstein (2001) Nat. Rev. Mol. Cell. Biol. 2, 422. Bromodomain recognition of acetyl-lysine Dhalluin et al. (1999) Nature 399, 491. Multiplicity of non-enzymatic domains in histone modifying enzymes Marmorstein (2001) Nat. Rev. Mol. Cell. Biol. 2, 422. Chromatin remodeling complexes. (e.g. SWI/SNF, ISWI, etc.) • Couples ATP hydrolysis with altering the nucleosome structure so that DNA binding proteins can access the DNA. • DNase I footprinting analysis shows that the 10 base periodicity of cutting disappears. • Gel shift and DNase I footprinting assays like those shown previously show that the chromatin remodeling complexes decrease the binding constant of proteins for nucleosomal DNA. Assays for Chromatin Remodeling Altered positioning Changes in restriction enzyme access. Narlikar et al. (2002) Cell 108, 475 A chromatin remodeling complex increases the accessibility of DNA to restriction enzyme cleavage in an ATP-dependent fashion. Saha et al. (2002) Genes & Dev. 16, 2120. Structures of representative remodeling complexes ISWI family SWI/SNF family • Generally multi-component. • The large catalytic subunits contains both ATPase and nonenzymatic domains. Narlikar et al. (2002) Cell 108, 475 Mechanisms of Remodeling • Sliding Vs. Eviction • Translational repositioning • Conformational change: induce twisting and/or bending of DNA. How is the activation process initiated by a DNA binding protein if the protein can’t bind the DNA in the first place? • Some DNA binding protein recognize their sites of the surface of a nucleosome - e.g. Glucocorticoid receptor. • Position the binding site in linker DNA. • In some cases, nucleosome associations are quite dynamic in the absence of activities that constrain their locations on the DNA so that DNA binding proteins are provide “windows of opportunity” to associate. Chromatin immunoprecipitation (ChIP): an assay for interaction of proteins with regulatory sequences in vivo. ChIP analysis of the ßinterferon gene. Agalioti et al. (2000) Cell 103, 667. Loading of complexes at the ß-interferon gene i. Activator ii. HAT complex SWI/SNF iii. SWI/SNF complex iv. GTFs and Pol II Narlikar et al. (2002) Cell 108, 475. Loading of complexes at the HO gene in yeast i. SWI/SNF ii. HAT complex iii. SWI/SNF complex iv. GTFs and Pol II Narlikar et al. (2002) Cell 108, 475. The time course of association of factors with the HO endonuclease gene in yeast. The chromatin remodeling complex binds to the promoter prior to the HAT, followed by Pol II and GTF’s. The remodeling process may depend on the initial status of the chromatin