Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
SNARE (protein) wikipedia , lookup
Cytoplasmic streaming wikipedia , lookup
Cell nucleus wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell culture wikipedia , lookup
Cell growth wikipedia , lookup
Cell encapsulation wikipedia , lookup
Extracellular matrix wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Signal transduction wikipedia , lookup
Cytokinesis wikipedia , lookup
Cell membrane wikipedia , lookup
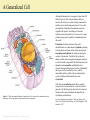
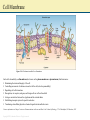
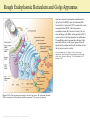
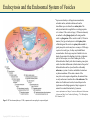
Chapter 2: Cytoplasm Color Textbook of Histology, 3rd ed. Gartner & Hiatt Copyright 2007 by Saunders/Elsevier. All rights reserved. Copyright 2007 by Saunders/Elsevier. All rights reserved. A Generalized Cell Although the human body is composed of more than 200 different types of cells, each performing a different function, all cells possess certain unifying characteristics and thus can be described in general terms. Every cell is surrounded by a bilipid plasma membrane, possesses organelles that permit it to discharge its functions, synthesizes macromolecules for its own use or for export, produces energy, and is capable of communicating with other cells. Protoplasm, the living substance of the cell, is subdivided into two compartments: cytoplasm, extending from the plasma membrane to the nuclear envelope and karyoplasm (nucleoplasm), the substance forming the contents of the nucleus. The bulk of the cytoplasm is water, in which various inorganic and organic chemicals are dissolved and/or suspended. This fluid suspension, the cytosol, contains organelles, metabolically active structures that perform distinctive functions. Additionally, the shapes of cells, their ability to move, and the intracellular pathways within cells are maintained by a system of tubules and filaments known as the cytoskeleton. Figure 2–5 Three-dimensional illustration of an idealized cell, as visualized by transmission electron microscopy. Various organelles and cytoskeletal elements are displayed. Copyright 2007 by Saunders/Elsevier. All rights reserved. Cells also contain inclusions, metabolic by-products, storage forms of various nutrients, or inert crystals and pigments. The following topics discuss the structure and functions of the major constituents of organelles, the cytoskeleton, and inclusions. For more information see Chapter 2 and 3 of Gartner and Hiatt: Color Textbook of Histology, 3rd ed. Philadelphia, W.B. Saunders, 2007 Cell Membrane Figure 2–8 A fluid mosaic model of the cell membrane. Each cell is bounded by a cell membrane (also known as the plasma membrane or plasmalemma) that functions in: Maintaining the structural integrity of the cell Controlling movements of substances in and out of the cell (selective permeability) Regulating cell–cell interactions Recognition, via receptors, antigens, and foreign cells as well as altered cells Acting as an interface between the cytoplasm and the external milieu Establishing transport systems for specific molecules Transducing extracellular physical or chemical signals into intracellular events. For more information see Chapter 2 section on Plasma membrane in Gartner and Hiatt: Color Textbook of Histology, 3 rd ed. Philadelphia, W.B. Saunders, 2007 Copyright 2007 by Saunders/Elsevier. All rights reserved. Rough Endoplasmic Reticulum and Golgi Apparatus Once the protein to be packaged is manufactured on the surface of the RER it enters the transitional ER from which it is released in COP II coated vesicles and transported to the ERGIC. Here the protein is examined to ensure that it is not a stowaway, that is it does not belong to the RER, and transported in COP I coated vesicles to the Golgi apparatus for modification. The modified protein is transported to the trans Golgi network for delivery to the proper region of the cell, namely into lysosomes, into the cell membrane, or into the cytosol as secretory vesicles. For more information see Chapter 2 section on the rough endoplasmic reticulum and the Golgi apparatus in Gartner and Hiatt: Color Textbook of Histology, 3rd ed. Philadelphia, W.B. Saunders, 2007. Figure 2–20 The Golgi apparatus and packaging in the trans Golgi network. ER, endoplasmic reticulum; ERGIC, endoplasmic reticulum/Golgi intermediate compartment; COP, coat protein (coatomer). Copyright 2007 by Saunders/Elsevier. All rights reserved. Protein Synthesis Proteins that will not be packaged are synthesized on ribosomes in the cytosol, whereas those that have to be packaged are synthesized on ribosomes on the rough endoplasmic reticulum (RER). As the mRNA enters the cytoplasm, it attaches to a small ribosomal subunit which has a binding site for mRNA, as well as three binding sites (P, A, and E) for tRNAs. Once the initiation process is completed, the start codon (AUG for the amino acid methionine) is recognized, and the initiator tRNA (bearing methionine) is attached to the P site (peptidyl-tRNA-binding site). Once this occurs, the large ribosomal subunit becomes bound, and protein synthesis is initiated. The next codon in the mRNA sequence is recognized by the proper amino acid bearing tRNA, which then binds to the A site (aminoacyl-tRNAbinding site). Methionine is uncoupled from the initiator tRNA (at the P site) and a peptide bond is formed between the two amino acids. The initiator tRNA is transferred to the E site (Exit site) and is released from the ribosome as the tRNA bearing the dipeptide transfers from the A site to the now empty P site. Figure 2–15 Protein synthesis in the cytosol. The next codon is recognized by the correct amino acid bearing tRNA and attaches to the A site. A peptide bond is formed between new amino acid and the dipeptide, forming a tripeptide as the dipeptide is uncoupled from the tRNA at the P site. The tRNA that lost its dipeptide is transferred to the E site and leaves the ribosome whereas the tRNA bearing the tripeptide moves from the A site to the P site. As this process is repeated the protein is formed by the constant addition of a new amino acid. For more information see Chapter 2 section on Protein Synthesis in Gartner and Hiatt: Color Textbook of Histology, 3rd ed. Philadelphia, W.B. Saunders, 2007 Copyright 2007 by Saunders/Elsevier. All rights reserved. Endocytosis and the Endosomal System of Vesicles The process whereby a cell ingests macromolecules, particulate matter, and other substances from the extracellular space is referred to as endocytosis. The endocytosed material is engulfed in a vesicle appropriate for its volume. If the vesicle is large (>250 nm in diameter), the method is called phagocytosis (cell eating) and the vesicle is a phagosome. If the vesicle is small (<150 nm in diameter), the type of endocytosis is called pinocytosis (cell drinking) and the vesicle is a pinocytotic vesicle. A typical pinocytotic vesicle may have as many as 1000 cargo receptors of several types, for they may bind different macromolecules. Each cargo receptor is linked to its own adaptin, the protein with a binding site for the cytoplasmic aspect of the receptor, as well as a binding site for the clathrin triskelion. Shortly after their formation, pinocytotic vesicles lose their clathrin coats (which return to the pool of clathrin triskelions in the cytosol) and fuse with early endosomes, a system of vesicles and tubules located near the plasma membrane. If the entire contents of the pinocytotic vesicle requires degradation, the material from the early endosome is transferred to a late endosome. This similar set of tubules and vesicles, located deeper in the cytoplasm near the Golgi apparatus, helps to prepare its contents for eventual destruction by lysosomes. For more information see Chapter 2 section on Endocytotic Mechanisms in Gartner and Hiatt: Color Textbook of Histology, 3rd ed. Philadelphia, W.B. Saunders, 2007. Figure 2–22 The endosomal pathways. CURL, compartment for uncoupling of receptor and ligand. Copyright 2007 by Saunders/Elsevier. All rights reserved. Mitochondrion Mitochondria are flexible, rod-shaped organelles, about 0.5 to 1 μm in girth and sometimes as much as 7 μm in length. Most animal cells possess a large number of mitochondria (as many as 2000 in each liver cell) because, via oxidative phosphorylation, they produce ATP, a stable storage form of energy that can be used by the cell for its various energyrequiring activities. Each mitochondrion possesses a smooth outer membrane and a folded inner membrane. The folds of the inner membrane, known as cristae, greatly increase the surface area of the membrane. The number of cristae possessed by a mitochondrion is related directly to the energy requirement of the cell; thus, a cardiac muscle cell mitochondrion has more cristae than an osteocyte mitochondrion has. The narrow space (10 to 20 nm in width) between the inner and outer membranes is called the intermembrane space, whereas the large space enclosed by the inner membrane is termed the matrix space (intercristal space). The contents of the two spaces differ somewhat. For more information see Chapter 2 section on Endocytotic Mechanisms in Gartner and Hiatt: Color Textbook of Histology, 3rd ed. Philadelphia, W.B. Saunders, 2007. Figure 2–28 The structure and function of mitochondria. A, Mitochondrion sectioned longitudinally to demonstrate its outer and folded inner membranes. B, Enlarged region of the mitochondrion, displaying the inner membrane subunits and ATP synthase. C, Two ATP synthase complexes and three of the five members of the electron transport chain that also function to pump hydrogen (H+) from the matrix into the intermembrane space. ADP, adenosine diphosphate; ATP, adenosine triphosphate; Pi, inorganic phosphate. Copyright 2007 by Saunders/Elsevier. All rights reserved. Cytoskeleton The cytoplasm of animal cells contains a cytoskeleton, an intricate three-dimensional meshwork of protein filaments that are responsible for the maintenance of cellular morphology. Additionally, the cytoskeleton is an active participant in cellular motion, whether of organelles or vesicles within the cytoplasm, regions of the cell, or the entire cell. The cytoskeleton has three components (A) mirotubules, (B) thin filaments, and (C) intermediate filaments. The only organelle that is composed of cytoskeletal elements is the centriole, a structure assembled from microtubules. For more information see Chapter 2 section on Cytoskeleton in Gartner and Hiatt: Color Textbook of Histology, 3rd ed. Philadelphia, W.B. Saunders, 2007 Figure 2–30 Elements of the cytoskeleton and centriole. A, Microtubule; B, thin filaments (actin); C, intermediate filaments; D, centriole. Copyright 2007 by Saunders/Elsevier. All rights reserved.