Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
SNARE (protein) wikipedia , lookup
Cell growth wikipedia , lookup
Cytoplasmic streaming wikipedia , lookup
Cell culture wikipedia , lookup
Cellular differentiation wikipedia , lookup
Tissue engineering wikipedia , lookup
Cell encapsulation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cell nucleus wikipedia , lookup
Cytokinesis wikipedia , lookup
Signal transduction wikipedia , lookup
Cell membrane wikipedia , lookup
Extracellular matrix wikipedia , lookup
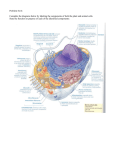
General Biology Chapter 2 Structure of the Cell Mohammed Al-Gayyar - 10 - Autumn 2012 General Biology Introduction to the cells The word cell comes from the Latin cellula, meaning "a small room". The cell is the basic structural and functional unit of all known living organisms. It is the smallest unit of life that is classified as a living thing, and is often called the building block of life. Some notes about cells should be kept in mind: § Nothing less than cell can be called living: The vital functions of an organism occur within cells. All cells come from preexisting. Like ourselves, the individual cells that form our bodies can grow, reproduce, process information, respond to stimuli and carry out an amazing array of chemical reactions. These abilities define life. Even simple unicellular organisms exhibit all the hallmark properties of life, indicating that the cell is the fundamental unit of life. § The Diversity: Organisms can be classified as unicellular (consisting of a single cell; including most bacteria) or multicellular (including plants and animals). Cells come in an amazing variety of sizes and shapes. Some move rapidly and have fast-changing structures. Others are largely stationary and structurally stable. Oxygen kills some cells but is an absolute requirement for others. § Similar basic chemistry: Despite the extraordinary diversity of plants and animals, all living things are fundamentally similar inside. Cells resemble one another to an astonishing degree in the details of their chemistry and sharing the same machinery for the most basic functions. All cells are composed of the same sorts of molecules that participate in the same types of chemical reactions. § Invention of the light microscope led to the discovery of cells: The descriptive term for the smallest living biological structure was coined by Robert Hooke in a book he in 1665 when he compared the cork cells he saw through his microscope to the small rooms. The name cell stuck even though the structures Hooke described were only the cell walls that remained after the living plant cells inside them had died. Types of cells: The biological universe consists of two types of cells: Mohammed Al-Gayyar - 11 - Autumn 2012 General Biology § Prokaryotic cells consist of a single closed compartment that is surrounded by the plasma membrane, lacks a defined nucleus, and has a relatively simple internal organization. Examples: Bacteria and Blue-green algae. § Eukaryotic cells: unlike prokaryotic cells, contain a defined membrane-bound nucleus and extensive internal membranes that enclose other compartments. The region of the cell lying between the plasma membrane and the nucleus is the cytoplasm, comprising the cytosol (aqueous phase) and the organelles. Eukaryotes comprise all members of the plant and animal kingdoms, including the fungi, which exist in both multicellular forms (molds) and unicellular forms (yeasts). Eukaryotes Complex in structure, with Prokaryotes nuclei and more complex in structure, with nuclei and membrane-bound organelles membrane-bound organelles Large (100 - 1000 µm) Small (1-10 µm) DNA in nucleus, bounded by membrane DNA circular, unbounded Genome consists of several chromosomes Genome consists of single chromosome Sexual reproduction common, by mitosis and Asexual reproduction common, not by mitosis meiosis or meiosis Mitochondria and other organelles present No general organelles Most forms are multicellular Most forms are singular Aerobic Anaerobic Mohammed Al-Gayyar - 12 - Autumn 2012 General Biology Biological Membranes Membranes Membranes are the outer boundary of individual cells and of certain organelles. Plasma membranes are the selectively permeable outermost structures of cells that separate the interior of the cell from the environment. Certain molecules are permitted to enter and exit the cell through transport across the plasma membrane. Components of biological membranes: All cell membranes are composed of the same materials: 1. Lipids Lipids are the most abundant type of macromolecule present. Plasma and organelle membranes contain between 40% and 80% lipid. There are three types of lipids are found: § Phospholipids: The most abundant of the membrane lipids are the phospholipids. They are polar, ionic compounds that are amphipathic (have both hydrophilic and hydrophobic components). The hydrophilic or polar portion is in the “head group”. Within the head group is the phosphate and an alcohol that is attached to it. The hydrophobic portion of the phospholipid is a long, hydrocarbon (structure of carbons and hydrogens) fatty acid tail. While the polar head groups of the outer leaflet extend outward toward the environment, the fatty acid tails extend inward. § Cholesterol: Another major component of cell membranes is cholesterol. An amphipathic molecule, cholesterol contains a polar hydroxyl group as well as a hydrophobic steroid ring and attached hydrocarbon. Cholesterol is dispersed throughout cell membranes, intercalating between phospholipids. Mohammed Al-Gayyar - 13 - Autumn 2012 General Biology Its polarhydroxyl group is near the polar head groups of the phospholipidswhile the steroid ring and hydrocarbon tails of cholesterol areoriented parallel to those of the phospholipids. Cholesterolfits into the spaces created by the kinks of the unsaturatedfatty acid tails, decreasing the ability of the fatty acids to undergomotion and therefore causing stiffening and strengthening of themembrane. § Glycolipids: Lipids with attached carbohydrate (sugars), glycolipidsare found in cell membranes in lower concentration than phospholipids and cholesterol. The carbohydrate portion is always oriented toward the outside of the cell, projecting into the environment. Glycolipids help to form the carbohydrate coat observed on cells and are involved in cellto-cell interactions. 2. Proteins While lipids form the main structure of the membrane, proteins arelargely responsible for many biological functions of the membrane. The types of proteins within a plasma membrane vary depending on the cell type. However, all membrane proteins are associated with membrane in one of three main ways: § Transmembrane proteins: They are embedded within the lipid bilayer of the membrane with structures that extend from the environment into the cytosol. All trans membrane proteins contain both hydrophilic and hydrophobic components. These proteins are oriented with their hydrophilic portions in contact with the aqueous exterior environment and with the cytosol and their hydrophobic portions in contact with the fatty acid tails of the phospholipids. § Lipid-anchored proteins: They are attached covalently to a portion of a lipid without entering the core portion of the bilayer of the membrane. Both trans membrane and lipidanchored proteins are integral membrane proteins since they can only be removed from a membrane by disrupting the entire membrane structure. § Peripheral membrane proteins: These proteins are located on the cytosolic side of the membrane and are only indirectly attached to the lipid of the membrane; they bind to other proteins that are attached to the lipids. Mohammed Al-Gayyar - 14 - Autumn 2012 General Biology Structure of biological membranes: The proteins and lipids of a cellular membrane are arranged in a certain way to form a stable outer structure of the cell. The membrane components, including lipids and proteins, are not fixed rigidly into a particular location. Both can exhibit several types of motions. Membrane proteins can also move laterally and can rotate. Despite its fluidity, the membrane structure is very stable and supportive for the cell. The arrangement of the phospholipids provides the basic structure which is then augmented by cholesterol, with functional roles played by proteins. The biological membranes have the following characters: § Bilayer arrangement: Membrane phospholipids are oriented with their hydrophobic fatty acid tails facing away from the polar, aqueous fluids of both the cytosol and the environment. The hydrophilic portions of the phospholipids are oriented toward the polar environment. Two layers of phospholipids are required to achieve this structure. § Asymmetry: The fatty acid tails of all the phospholipids are structurally very similar to each other. Some phospholipids are found on the outer leaflet while others are more commonly seen on the inner leaflet. In addition, glycolipids are differentially arranged as well and are always on the outer leaflet with their attached carbohydrate projecting away from the cell. § Fluid mosaic model: The membrane is described as a fluid, owing to the ability of lipids to diffuse laterally. The overall structure is likened to a flowing sea. Membrane proteins are dispersed throughout the membrane. Many of the membrane proteins retain the ability to undergo lateral motion and are likened to icebergs floating within the sea of lipids. Mohammed Al-Gayyar - 15 - Autumn 2012 General Biology Organelles Organelles are complex intracellular locations where processes necessary for eukaryotic cellular life occur. Most organelles are membrane-enclosed structures. Their membranes are composed of the same components asplasma membranes that form the outer boundaries of cells. Together with the cytosol (liquid portion of the cytoskeleton), the organelles help to form the cytoplasm, composed of all materials contained within the boundaries of the plasma membrane. Organelles do not float freely within the cytosol but are interconnected and joined by the framework established by proteins of the cytoskeleton. Each organelle carries out a specific function. Nucleus (plural = nuclei): All eukaryotic cells except mature erythrocytes (red blood cells) contain a nucleus where the cell’s genomic DNA resides. The outermost structure of the nucleus is the nuclear envelope. This is a double-layered phospholipid membrane with nuclear pores to permit transfer of materials between the nucleus and the cytosol. The interior of the nucleus contains the nucleoplasm (the fluid in which the DNAs are found). Within the nucleus there is a suborganelle called the nucleolus. The nucleolus is the site of ribosome production. Mohammed Al-Gayyar - 16 - Autumn 2012 General Biology Ribosomes: Ribosomes are the cellular machinery for protein synthesis. They are composed of proteins and ribosomal RNA (rRNA) with approximately 40% being protein and 60% rRNA. Ribosomes are found within the cytosol either free or bound to the endoplasmic reticulum. Endoplasmic reticulum (ER): ER is often observed to surround the nucleus. The outer layer of the nuclear envelope is actually contiguous with the ER. The ER forms a maze of membrane-enclosed, interconnected spaces that constitute the ER lumen Regions of ER where ribosomes are bound to the outer membrane are called rough endoplasmic reticulum (rER). Bound ribosomes and the associated ER are involved in the production and modification of proteins. Smooth endoplasmic reticulum (sER) refers to the regions of ER without attached ribosomes. Both rER and sER function in the glycosylation (addition of carbohydrate) of proteins and in the synthesis of lipids. Golgi complex: It appears as flat, stacked, membranous sacs. Three regions are described within the Golgi complex: the cis, which is closest to the ER; the medial; and the trans Golgi, which is near the plasma membrane. Each region is responsible for performing distinct modifications to the newly synthesized proteins, such as: § Glycosylations (addition of carbohydrate) § Phosphorylations (addition of phosphate) § Proteolysis (enzyme-mediated breakdown of protein) Mitochondrion (plural = mitochondria): Complex organelles, mitochondria have several important functions in eukaryotic cells. Their unique membranes are used to generate ATP (greatly increasing the energy yield from the Mohammed Al-Gayyar - 17 - Autumn 2012 General Biology breakdown of carbohydrates and lipids). The very survival of individual cells depends on the integrity of their mitochondria. One characteristic feature of mitochondria is the double phospholipidbilayer membranes that form the outer boundary of the organelle. The inner mitochondrial membrane forms folded structures called cristae that protrude into the mitochondrial lumen known as the mitochondrial matrix. Lysosomes: Lysosomes are membrane-enclosed organelles of various sizes that havean acidic internal pH (pH 5). Lysosomes containpotent enzymes known collectively as acid hydrolases. They function within the acidic environment of lysosomes to hydrolyze or break down macromolecules (proteins, nucleic acids, carbohydrates and lipids). Nonfunctional macromoleculesbuild up to toxic levels if they are not degraded within lysosomes andproperly recycled for reuse within the cell.In addition, lysosomal enzymes also degrade materials that have beentaken up by the cell through endocytosis or phagocytosis. Peroxisomes: Peroxisomes resemble lysosomes in size and in structure. They havesingle membranes enclosing them and contain hydrolytic enzymes. It helps in: § Break down of fatty acids and purines (AMP and GMP). § Detoxification of hydrogen peroxide (a toxic by-product of many metabolic reactions). § Synthesis of myelin (the substance that forms a protective sheath around many neurons). § In liver cells, peroxisomes participate in cholesterol and bile acid synthesis. Mohammed Al-Gayyar - 18 - Autumn 2012 General Biology Cytoskeleton The cytoskeleton is a complex network of protein filaments that establish a supportive scaffolding system within the cell. Cytoskeletal proteins are located throughout the interior of the cell, anchored to the plasma membrane and traversing the cytoplasm. Organelles reside within the framework established by the cytoskeleton. The cytoskeleton is not simply a passive internal skeleton but is a dynamic regulatory feature of the cell. These components of the cytoskeleton work together as an integrated network of support within the cytoplasm. Actin: Actin helps to establish a cytoplasmic protein framework known as microfilaments visualized radiating out from the nucleus to the lipid bilayer of the plasma membrane. Some forms of actin are found only in muscle cells, while other forms of actin are found within the cytoplasm of most cell types. Functions of actin in the cytoplasm of non muscle cells include: § Regulation of the physical state of the cytosol § Cell movement § Formation of contractile rings in cell division § Within the nucleus, actin is involved in the regulation of gene transcription. § Regulators of the gel/sol of the cytosol: One characteristic of a cell is the physical nature of its cytosol. It can be described either as gel, a more firm state, or sol, a more soluble state. The more structured the actin, the firmer (gel) the cytosol. The less structured (more fragmented) the actin, the more soluble (sol) the cytosol. Actin is Mohammed Al-Gayyar - 19 - Autumn 2012 General Biology continuously tread-milling in both the gel and sol states, contributing to the character of the cytoplasm. Intermediate filaments: They are larger than actin microfilaments and smaller than microtubules. Most intermediate filaments are located in the cytosol between the nuclear envelope and the plasma membrane. They provide structural stability to the cytoplasm, somewhat reminiscent of the way that steel rods can reinforce concrete. There are six categories of intermediate filaments, grouped by their location. Examples include keratins, vimentin and neuroflaments Microtubules: Microtubules are the last type of predominant structure observed in the cytoskeleton. The structure of a microtubule resembles a hollow cylindrical tube. They are involved in: § Chromosomal movements during nuclear divisions § Formation of cilia and flagella in certain cell types § Intracellular transport Mohammed Al-Gayyar - 20 - Autumn 2012 General Biology Extracellular Matrix A substantial part of tissue volume is an extracellular space largely filled by the intricate network of macromolecules of the extracellular matrix (ECM). The ECM is specialized to perform different functions in different tissues. For example, the ECM adds strength to tendons and is involved in filtration in the kidney and attachment in skin. The physical nature of the ECM also varies from tissue to tissue. Blood is fluid, while cartilage has a spongy characteristic owing to the nature of extracellular materials in those tissues. Three categories of extra cellular macromolecules make up the ECM: 1. Proteoglycans: Proteoglycans are aggregates of glycosaminoglycans (GAGs) and proteins. GAGs arealso known are composed of repeating disaccharide chains where one of the sugars is an amino sugar and the other is an acidic sugar. They are organized in long, unbranched chains. GAGs contain multiple negative charges and are extended in solution. The most prevalent GAG is chondroitin sulfate. Other GAGs include hyaluronic acid, keratin sulfate, dermatan sulfate, heparin and heparan sulfate. Because of their net negative surface charges, GAGs repel each other. In solution, GAGs tend to slide past each other, producing the slippery consistency we associate with mucous secretions. The bones of the joint are cushioned by the water balloon–like structure of the hydrated GAGs in the cartilage. When compressive forces are exerted on it, the water is forced out and the GAGs occupy a smaller volume. When the force of compression is released, water floods back in, rehydrating the GAGs, much like a dried sponge rapidly soaking up water. 2. Fibrous proteins: Fibrous proteins are extended molecules that serve structural functions in tissues. There are two types of fibrous proteins: § Collagen: The most abundant protein in the human body, collagen forms tough protein fibers that are resistant to shearing forces. Collagen is the main type of protein in bone, Mohammed Al-Gayyar - 21 - Autumn 2012 General Biology tendon and skin. In the ECM, collagen is dispersed as a gel-like substance and provides support and strength. Collagen is a family of proteins, with 28 distinct types. However, over 90% of collagen in the human body is in collagen types I, II, III, and IV. Together the collagens constitute 25% of total body protein mass. § Elastin: The other major fibrous protein in the ECM is elastin. Elastic fibers formed by elastin enable skin, arteries and lungs to stretch and recoil without tearing. The structure of elastin is that of an interconnected rubbery network that can impart stretchiness to the tissue that contains it. This structure resembles a collection of rubber bands that have been knotted together. 3. Adhesive proteins: The last category of ECM components consists of proteins that join together and organize the ECM and also link cells to the ECM. Fibronectin and laminin are adhesive glycoproteins secreted by cells into the extracellular space. Both are considered multifunctional proteins because they contain three different binding domains that link them to cell surfaces and to other components of the ECM, including proteoglycans and collagen. Through their interactions with fibronectin or laminin, proteoglycans and collagen are linked to each other and to a cell’s surface. Thus, adhesive proteins join ECM components to each other and link cells to the ECM. Mohammed Al-Gayyar - 22 - Autumn 2012 General Biology Cell Adhesion A growing tissue is able to form because the member cells remain attached and do not travel elsewhere. Cell junctions are important in maintaining the structure of a tissue as well as its integrity. They are composed of a collection of individual cell adhesion molecules. Cell adhesion molecules mediate selective cell-to-cell and cell-to-ECM adhesion. These are all transmembrane proteins that are embedded within the plasma membranes of cells. They extend from the cytoplasm through the plasma membrane to the extracellular space. In the extracellular space, they bind specifically to their ligands, which may be: § Cell adhesion molecules on other cells § Certain molecules on the surface of other cells § Components of the ECM Four families of adhesion molecules function in cell-cell adhesion: the cadherins, the selectins, the immunoglobulin super family, and the integrins. The integrins also function in cell-to-ECM adhesion. Cadherins: The cell adhesion molecules that are important in holding cells together to maintain the integrity of a tissue are called the cadherins. These transmembrane linker proteins contain extracellular domains that bind to a cadherin on another cell. Cadherins also have intracellular domains that bind to the actin cytoskeleton. Therefore, when two cells are linked together via cadherins, their actin cytoskeletons are indirectly linked as well. Calcium is required for cadherin binding to another cadherin. Adhesion mediated by cadherins is long-lasting and important in maintaining the tissue structure. Selectins: Selectins mediate more transient cell-to-cell adhesions. They are particularly important in the immune system in mediating white blood cell migration to sites of inflammation. Mohammed Al-Gayyar - 23 - Autumn 2012 General Biology Selectins are named for their “lectin” or carbohydrate-binding domain in the extracellular portionof their structure. A selectin on one cell interacts with a carbohydrate-containing ligand on another cell. Immunoglobulin superfamily: They are named because they share certain structural characteristics of immunoglobulins (antibodies). These members of the immunoglobulin superfamily of adhesion molecules regulate cell-to-cell adhesions. Some immunoglobulin superfamily members facilitate adhesion of leukocytes to endothelial cells lining the blood vessels during injury and stress. Integrins: Both cell-to-cell and cell-to-ECM adhesions are mediated by integrins. Members of this family of transmembrane proteins bind to their ligands with relatively low affinity; multiple weak adhesive interactions characterize integrin binding and function. Integrins consist of two transmembrane chains, α and β. When integrins mediate cell-to-cell adhesions, their ligands are members of the immunoglobulin superfamily. When integrins join a cell to the ECM, collagen and fibronectin commonly serve as their ligands. Therefore, Integrins mediate interactions between the cytoskeleton within the cell and the ECM surrounding the cell. Mohammed Al-Gayyar - 24 - Autumn 2012