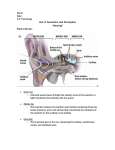
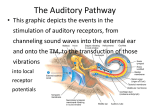
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Node of Ranvier wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Neural engineering wikipedia , lookup
Biological neuron model wikipedia , lookup
Sensory cue wikipedia , lookup
Sound localization wikipedia , lookup
Patch clamp wikipedia , lookup
Action potential wikipedia , lookup
Membrane potential wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Animal echolocation wikipedia , lookup
Circumventricular organs wikipedia , lookup
Neuroregeneration wikipedia , lookup
Single-unit recording wikipedia , lookup
Neurostimulation wikipedia , lookup
Resting potential wikipedia , lookup
End-plate potential wikipedia , lookup
Electrophysiology wikipedia , lookup
Microneurography wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Perception of infrasound wikipedia , lookup
PHYSIOLOGICAL THE AMERICAN PHYSIOLOGICAL JANUARY VOLaME 37 Biophysics the Central Institute SOCIETY, INC. NUMBRR I x957 and Physiology HALLOWELL From REVIEWS of the Inner Earl DAVIS for the Deaf, St. LOGS, Wssowi SCOPE HIS REVIEW of the biophysics and physiology of the cochlea is arbitrarily limited in scope, both in regard to the period covered and in regard to subject matter. Other review articles cover quite adequately the earlier years and many contiguous areas. The selection of topics, and particularly the relative emphasis devoted to them, reflect the personal interests and value judgments of the reviewer. The reviewer offers his apologies to those authors whose works should logically be mentioned here but which have not come to his attention. He has not attempted to search the literature and cite every relevant paper. This function is performed periodically by Annual Reviews of Physiology and Annual Reviews of Psychology. Also a special Bibliography on Hearing (223) includes the physiology of the cochlea within its larger scope. We shall not enter the area of transmission of sound to the cochlea, either by air or by bone conduction. Wever and Lawrence, in Physiological Acozcstics(249), reviewed this subject very competently in historical perspective and in a summary of experimental studies which includes their own important contributions. Part of the chapter by Bekesy and Rosenblith on “The Mechanical Properties of the Ear” in the Handbook of Experimental Psychology (222) deals with the middle ear and serves as the best companion review to Physiological Acozlsticson this subject. The remainder of Chapter 27 and also most of Chapter 28 on “The Psychophysiology of Hearing and Deafness” are the direct foundations for much of the present review. Fletcher’s Speechand Hear&g in Commwication (89) deals chiefly with human reactions and responses or with acoustic analyses but has chapters important for cochlear dynamics and the theory of hearing. We shall exclude the entire subject of psychoacoustics, i.e., what a person hears, what he can discriminate, how his hearing can be measured, and so forth, including the interactions of the two ears in providing cues for the localization of the source of sound. These topics are well reviewed in Licklider’s chapter in the Handbook of Experimental Psychology (222) and Measurementof Hearing by I. J. Hirsh (I 26). We shall practically confine ourselves to the cochlea itself and chiefly to the animal cochlea as studied in the laboratory. We shall not follow the neural pathways beyond the cochlear nucleus, as Galambos (92) has very recently reviewed the auditory path1Prepared under Contract N6onr-272 between Central Institute for the Deaf and the Office of Naval Research. Reproduction is permitted for any purpose of the United States Government. I HALLOWELL 2 DAVIS Votumc 37 ways of the mid-brain and the auditory cortex. For the biology and the comparative physiology of hearing we refer to the excellent articles by Pumphrey (175), by Prosser (174) and by Autrum (8). Topics that remain for present review, after this drastic fence-building are: I) The anatomy and biochemistry of the cochlea. 2) Injury to the ear, particularly by intense sound and by drugs. 3) The dynamics of the cochlea, including mathematical theories of its action. 4) The electric potentials of the cochlea. 5) Auditory nerve impulses. 6) The physiological background of cochlear activity. 7) Theories of cochlear function. ANATOMY AND BIOCHEMISTRY OF THE COCHLEA Anatomists and pathologists correctly stressthe importance of structure as the substrate of biophysics and physiology. The reviewer observes,however, after reading several recent anatomical articles, that the situation is reciprocal. Consideration of physical and physiological principles is helpful when speculating as to the probable functions of the structures that are observed under the microscope. Dimensions of Cochlea. Theories of the dynamics of the cochlea, and particularly the mathematical treatments which undertake to explain how the cochlea operates as an acoustic analyzer, depend on the gradation of size and physical characteristics of the structures of the cochlea. Wever’s (241) measurementsof 25 human cochleas give the cross-sectionareas of the scalae of the cochlea and the sensory structures, the width of basilar membrane, etc., as functions of distance along the basilar membrane, and Fernandez (84) gives similar data for the guinea-pig. There are two estimates of the distances along the basilar membrane that correspond to the turns of the guinea pig cochlea (84, IS), but there is a slight discrepancy between them (230). Tectorial Membrane. In preparations of the cochlea that have been fixed for staining and sectioning, the tectorial membrane is badly shrunken and distorted (164, 165). Recent microdissectionsof fresh specimens,both human and guinea pig, using conventional illumination with simple stains or phase-contrast microscopy (239, 136, 122, 21), show clearly that the tectorial membrane is a spiral ribbon, attached at one edge to the limbus and by one surface and its other edge to the organ of Corti. The outer edge attaches by a filmy meshwork border sheet to the region of Hensen’scells beyond the external hair cells. It also attaches at Hensen’sstripe just inside the inner hair cells, and the hairs of the hair cells are apparently imbedded in the tectorial membrane. All investigators agree that the membrane is composedof two kinds of extracellular material, a system of fibrils that run diagonally outward and an amorphous, poorly staining interfibrillar material. Differences of opinion (32, 122) as to whether the tectorial membraneis formed (secreted) by the cells of the limbus or by the hair cells have little bearing on the problem of function. The membrane is nearly transparent, but it hasappreciable tensile strength and considerablestiffness (IS, 16). The interfibrillar material seemsto be a colloidal gel, but its chemical composition can only be inferred from staining reactions, the action of enzymes, the uptake of radioactive tracers, etc. (260, 172, 28). The material seemsto be of protein nature with free carbonyl and bound disulphide groups. It seemsto be either a glycoprotein (260) or, less probably, a mucopolysaccharide (28). The earlier opinion that keratin is January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 3 present is not substantiated. Whatever their exact composition, the tectorial membrane, the otolithic membranesand the cupulaeof the semicircular canals have very similar histological and histochemical properties. Organ of Corti. Wolff (261) has reviewed the microscopic anatomy of the organ of Corti “as we seeit today, one hundred years after its discovery.” Phase-contrast microscopy has since confirmed many details that previously were uncertain (136, 122). Still finer details of structure have been revealed by electron microscopy (77-80, For example, the hairs of the hair cells (guinea pig) are as many as 70 to a cell. The intracellular rootlet of each hair has a diameter of about 0.1 p and extends through the cuticular layer into the subcuticular region. The hairs proper are about 4 p long and show no fibrillar structure (78). The internal hair cells differ in detail from the external hair cells sufficiently to suggestimportant differences in function (217). The reticular lamina is a dense granular structure. The outer (endolymphatic) surfacesof Hensen’scells, but not their other surfaces, are covered with fine fingerlike protoplasmic processeswhich suggest an absorptive function (80). The older descriptions of the supporting structures are confirmed and extended (79). We condenseEngstrijm and Wersall’s (79) description of the nerve endings on the hair cellsas follows: 217). The nerve fibers do not enter Deiters cells but continue their intercellular course and insinuate themselves between hair cell and Deiters cell. Here they terminate in club-like intercellular bodies, one for each fiber. A bundle of about 20-30 neurofibrils, each about 0.1 p in diameter, indents the membrane of the Deiters cell and splits up into separate neurofibrils, beneath the base of the hair cell but outside of the Deiters cell. Each terminates in a clubshaped nerve ending. The size of these endings varies within wide limits, down to less than I p in diameter. The larger nerve endings may collect in clusters covering an area of 4 by 5 p, resembling synaptic regions. They are intimately attached to the cellular membrane but the membrane is distinct and clearly not pierced. The club-shaped end organ also has a distinctly visible membranous envelope. The hair cell thus has a well-developed end-organ in the region between its own base and the Deiters cell. Even though the outer and inner hair cells present some very marked structural differences, their nerve endings show a remarkable similarity. The nerve fibers lose their myelin sheaths as they pass through the habenula perforata. Within the fluid spacesof the organ of Corti they are completely bare without even the usual sheath of Schwann (217). Innervation. Recent studies have confirmed the classical description (cf 224, 242) of the innervation of the hair cells. There are radial fibers running to both in& and outer hair cells and each fiber innervates only one or two hair cells. These radial fibers are the chief supply to the inner hair cells. The external hair cells,however, are more richly innervated by a secondclassof fiber, the spiral fibers. There are several bundles of them that run lengthwise of the organ of Corti, often for distances of several millimeters. It now appears (83) that these fibers run in both directions and not merely basalward. Each spiral fiber innervates many hair cells, but only every secondor third cell along its course. Multiple innervation of each hair cell is the rule rather than the exception. There is also a system of spiral fibers, the intraganglionic spiral bundle, that innervates the inner hair cells, although sparsely. The intraganglionic spiral bundle is a continuation of the tract of Rasmussen (182) by way of the bundle of Oort in the auditory nerve. The cell bodies of this A-,-L tract --are --A nor; locatea in tne spiral ganglion or Corti witnin tne moaiolus but are l- -- L-2 z- LX- --Z---l -_____ l?-_- -L n---L? --!A1.2-- Al - - l! 1 1 -1 --_- 4 HALLOWELL DAVIS Volume 37 located in the medulla in the superior olivary complex of the opposite side. Femandez (83) has traced fibers of this tract to the internal hair cells. Rasmussen (183) expresses some reservations as to whether the internal hair cells are the only cells so innervated. These fibers from the opposite side of the brain stem seem to be efferent fibers. The presence of the olivo-cochlear bundle is in keeping with the modern concept (92, 183) that there is an efferent ‘feedback’ system all the way from cortex to cochlea. A direct cochlea-cochlear pathway was postulated in a preliminary report (96) on the basis of an apparent peripheral interaction between nerve impulses stimulated by a click in one ear and those initiated by a separate later click in the other. Unfortunately the preliminary results were not confirmed in later experiments and the conclusions were withdrawn. Blood Supply and Formation of Cochlear Fluids. The arterioles and capillaries (guinea pig) form an intricate pattern, particularly in the stria vascularis, which is a specialized organ that is clearly differentiated and separated from the loose meshwork of spiral ligament behind it. Smooth muscle cells are described in the walls of arterioles and in capillaries of the cochlea (except in the stria vascularis) and non-myelinated nerve fibers accompany the cochlear artery in the modiolus (216). Perilymph. The perilymph, which fills Scala tympani, Scala vestibuli and the large perilymphatic spaces of the vestibule and semicircular canals, is practically identical with cerebrospinal fluid in its electrolyte composition. There has been disagreement in the past as to the protein content of perilymph and of endolymph also, but it now seems (45, 152, 219, 218) that endolymph contains no more protein than cerebrospinal fluid, and that perilymph may contain twice as much. The perilymphatic space communicates with the cerebrospinal space by the cochlear aqueduct and by perineural and perivascular spaces in the internal auditory meatus. In the guinea pig, cerebrospinal fluid certainly can enter by these channels when an artificial opening is made into the cochlea. Red blood cells and India ink (200), iron salts (4) and fluorescin (107) have all been shown to enter in this way, but it is not certain that such a flow of cerebrospinal fluid is the sole source in man under normal conditions. In any case, radioactive sodium (Na21 appears promptly in the perilymph following an intravenous injection (107), and the concept that perilymph is essentially an ultrafiltrate of serum seems well founded. The resorption of perilymph is generally believed to occur into the capillaries of the spiral ligament (200). There seems to be no significant barrier to diffusion of fluid or to electric current flow between Scala tympani and the interior of the spiral ligament. Endolymph. The endolymph fills the cochlear duct, the utricle and the remainder of the membranous labyrinth, including its extension through the ductus endolymphaticus to the saccus endolymphaticus. The saccus lies under the brain, between split layers of the dura mater, near the transverse venous sinus. The endolymphatic system is mechanically closed and fluid does not readily pass through the walls (154). Specialized areas in the walls of the membranous labyrinth include the walls of the saccus endolymphaticus (I I ra), the stria vascularis (and the analogous structures near the sense organs of saccule, utricle and ampullae), and the organ of Corti, the macula and other sensory structures. Since the classical work of Guild (I na) it has generally been believed that the endolymph is formed by the stria vascularis and resorbed in the saccus endolymphaticus. Recently Lindsay and his collaborators (I$, 2o6a) have cast doubt on the importance of the saccus endolymphaticus, at least in the cat. They destroyed the saccus endolyphaticus surgically but found no Janzcary rg57 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 5 histological or functional injury. Auditory acuity, tested by conditioned reflexes, was normal (three animals) after periods up to g months. Apparently the endolymphatic sac is not an essential structure in the cat. Adequate interchange of fluid and other substance can occur in and out of stria vascularis (200), and perhaps the external sulcus cells (203) or others may assist in resorption. Apparently some organic substances such as fluorescin can pass through the walls of the membranous labyrinth under certain experimental conditions, but it now seems clear that the walls are practically impermeable to ions. One piece of evidence is the high electrical resistance to direct current and to low-frequency alternating current (IS). Another is the slow appearance of Na24 in the endolymph (107). Still another is the unique chemical composition of the endolymph (218, 45). The chloride concentration of endolymph is only go per cent of that of perilymph and cerebrospinal fluid. Much more surprising is the very high potassium and the very low sodium content. The K concentration in endolymph is 30 times that of perilymph, the Na concentration is one-tenth that of perilymph. The K/Na ratio is practically that of intracellular fluid. This situation is unique among extra-cellular body fluids. Obviously endolymph is not an ultrafltrate of blood plasma. It must be produced by a process of active secretion analogous to that in the tubules of the kidney or the gastric mucosa. The stria vascularis is the most popular candidate for such a secretory organ. Incidentally, Smith, Lowry and Wu (218) do not make any mention of the high viscosity often attributed to the endolymph. They do write, “the stria vascularis was pierced by a small micropipette (40 p outside diameter). A small amount of fluid immediately (italics ours) ran up by capillary action.” It will be recalled that the protein content is the same as cerebrospinal fluid. On the other hand the endolymph in the shark is quite viscous and sets into a jelly after withdrawal (237). This consistency seems more like the tectorial membrane in mammals than like their endolymph* INJURY OF THE COCHLEA Three types of injury to the cochlea merit brief mention in this review because they give some insight into the normal mode of action of the cochlea. The agents in question are I) drugs, z) intense sound and 3) experimental surgery. Schuknecht also gives a useful review of lesions of the organ of Corti (204). Drugs. Some drugs have proved very useful in the treatment of tuberculosis, but unfortunately, when given in the necessary dosages, they show toxic side-effects on the vestibular and, to a less extent, on the auditory system. An extensive clinical literature has debated whether the primary effect of streptomycin is on the vestibular nucleus of the medulla oblongata or on the end organ. It is now generally accepted that whatever the direct neural effects may be, streptomycin does injure the sensory epithelium of the vestibular system and, less frequently, the hair cells of the cochlea as well (44, 202, 29, 120, 106, 206, 189). Similar effects can be produced in man, cat, guinea pig and mouse, although there are said to be significant differences among species. For example, dihydrostreptomycin is more injurious to the human cochlea than streptomycin but the reverse relation is claimed for the cat, and in cats neomycin has little effect on vestibular function but causes severe injury to the organ of COrti(I21, 134). The hair cells are the most vulnerable structures. The changes in stria vascularis, for example, require larger doses and may only be secondary results of the 6 HALLOWELL DAVIS Vt7Zzme37 degeneration of the organ of Corti (200). Particularly interesting is the special susceptibility to streptomycin of the internal hair cell of the cat (120). This is in contrast to the greater susceptibility of the external hair cells to all other toxic and traumatic agents,-such as quinine (202, 46), arsacetine (44), acoustic trauma (48, 56) and obstruction to blood flow (138). It is not surprising that in animals poisoned with quinine, or dihydrostreptomycin (120, 121), or affected by anoxia, acoustic trauma, etc., the action potentials (of the auditory nerve) should be depressedmore than the cochlear microphonic (ultimately related to the hair cells). It is important, however, that streptomycin reduces the cochlear microphonic with relatively little depression of the action potentials until the injury becomesvery severe. More specifically, it is the matimzcm voltage of the microphonic which is depressed-not, as with a conductive lesion, the ‘threshold’ which is raised (118). In severe intoxication the intensity neededto elicit a small standard microphonic responseis also elevated. In general, the cochlear microphonic is depressedrelatively more than the action potentials. Quinine, on the other hand, may selectively abolish the action potentials (60), perhaps by direct injury to the nerve endings (47), perhaps otherwise (67). Acoustic Trauma and Chronic Noise-Induced Hearing Loss. The subject of hearing lossesthat are induced by exposure to noise or pure tones is worthy of a monograph in itself but, in spite of current medical and legal interest in the problem, it will not be consideredhere except asit relates to known anatomical or physiological changes.This limitation will exclude practically all studies on man. Surveys of such topics as temporary changesin auditory threshold, in the loudnessof tones heard above threshold, in pitch, etc., induced by exposure to pure tones or to noise, will be found in the monographsby Riiedi and Furrer (201), by Kryter (146) and by Davis et al. (57). Suffice it to say that this temporary hearing lossin man is usually greatest for a frequency approximately one-half octave above the exposure tone, and that no physical, anatomical or physiological correlates have been establishedfor this upward shift. Even better known in man is the permanent hearing loss that results from exposuresrepeated over months and years to loud industrial noises,gunfire, etc. These hearing lossesare permanent and the pathology is generally believed to be atrophy of the hair cells, particularly the external hair cellsin the basal turn. The hearing loss usually appears first, and later is most severe, at the frequency of 4000 cycles per second.The amount of hearing lossincurred at a particular frequency has been only very roughly correlated with the spectrum of the noisewhich has presumably caused the hearing loss (7). The correlations are best with the intensity of the noise in the octave band one or even two octaves below the frequency at which the hearing lossis measured, but the report cited points out specifically that this does not imply that the hearing losswas causedby the energy at these lower frequencies. What has been establishedis only a tentative statistical correlation. Permanent hearing loss from chronic exposure to loud noise or tones has been very inadequately studied in animals. In one of the few direct comparisons (I&, guinea pigs were exposed to the noise of a boiler factory for g months. Histological study showed atrophy of hair cells, particularly the external hair cells in the basal turn. In another study (I-3), guinea pigs were exposedin a jet engine test cell with similar results. Attempts were made to assessthe functional hearing lossby meansof the electrical output of the cochlea (the cochlear microphonic). Unfortunately, this method of assessmentis wrong in principle unlessmultiple setsof intracochlear elec- January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 7 trodes are employed (cf. 56). What is needed in the experimental study of chronic noise-induced hearing loss is a set of animal exposures of this sort with accurate measurements of the intensity and the spectrum of the noise, with regular exposures for many months, with hearing assessed by behavioral methods (207), and with histological control. Also, the relation of noise-induced hearing loss to the loss of hearing expected with advancing age (presbycusis) should be examined in man and in animals. The losses appear very similar (205) and it is usually assumed that the two kinds of loss are independent but additive (7). This is merely an assumption, however. We have, then, three forms of hearing loss that are induced by noise: I) temporary threshold shift, which has been studied almost exclusively in man and is produced by exposure to tones or noises over IOO decibels (relative to 0.0002 microbar) in intensity; 2) permanent hearing loss that is produced in man by repeated exposure to loud sounds over months or years and is also called ‘occupational’ hearing loss; and 3) permanent hearing loss produced by very brief exposure to very loud noise. It has been proposed that the term ‘acoustic trauma’ be reserved for this third type. Experimental study of such acoustic trauma is confined to animals. These three forms of hearing loss are clearly different in many respects and it is very unfortunate that so little is known of the relations among them. We do not know, for example, whether susceptibility to temporary threshold shift is a valid index of susceptibility to either occupational hearing loss or acoustic trauma. ‘Tests’ based on temporary threshold shift have been suggested repeatedly, but without validation. Another test for ‘prediction of susceptibility to acoustic trauma’ is based on a measurement of the threshold for distortion, i.e., the production of aural harmonics (150). The argument is ingenious and the assumptions are plausible, but they are not acceptable substitutes for direct validation. In spite of the positive phrasing of the authors’ statements, this ‘test’ cannot be accepted as anything more than a working hypothesis. We shall not review the details of the anatomical injuries to the cochlea that can be produced by very intense sustained sound. Many descriptions are available (201, 160, 48, 220, 221, 56, 1-3, 142, 143). The injuries closely resemble those described many years ago as the result of pistol shots (III). The most severe injury is to the external hair cells, the internal hair cells and the supporting cells of the organ of Corti, in roughly that order but with much overlap. Complete destruction of the hair cells and even of the entire organ of Corti over considerable sections of the basilar membrane can be quite easily produced by tones of the order of 145-150 decibels. The intensity (sound pressure level) and the duration of exposure are important determinants of the severity of injury but, contrary to previous impressions (cf. 221), the severity is not strongly related to frequency (56). Specifically, in the guinea pig, if the acoustic pressure is measured at the ear drum, a given severity of injury (presumably permanent) could be produced by a one-minute exposure at an intensity of 144 decibels to any one of the four following frequencies: 185, 545, 2000 or 8000 cycles per second. ‘Severity’ is defined here as the degree of most serious injury to any part of the organ of Corti, and not in terms of extent of injury along the basilar membrane. It is dangerous to generalize too far from this single experiment, however. It would be even more dangerous to undertake to decide at the present time whether it is the amplitude of displacement, the particle velocity, the acceleration or the total energy that is correlated best with and is presumably the mechanism of the production of injury. 8 HALLOWELL DAVIS Volume 37 In general the basal turn of the cochlea is injured most by high-frequency sounds (4000 cps and higher) and the apical turns by low tones (500 cps and below). The localization is not precise, however, and the injury, if severe enough to be unmistakable under the microscope, is usually quite widespread, particularly when produced by low tones (2 20). Even after several weeks of recovery (48, 2), although details of the injuries are dBerent, the localization is still difYuse. These observations belong to ‘acoustic trauma’ rather than to ‘chronic noiseinduced hearing loss.’ They are broadly consistent with the place principle in auditory theory, but precise analysis is confused by the known scattering of acoustic energy by non-linear distortion into frequency bands above the exposure tone. Furthermore, energy may appear in subharmonic frequencies as well (61, 97, 75, 76). In any case, as we shall see below, the present concept of the place principle assumes a very broad zone of activity for low tones. Anatomical studies of acoustic trauma in which the cochlear microphonics were tested confirm the dependence of the cochlear microphonics on the hair cells (56, 142). This evidence is important for the question of the origin of the microphonics. An interesting side issue is the evidence (208), based on experiments on cats, that many of the permanent hearing losses that are produced by violent blows on the head, as in automobile accidents, are actually acoustic trauma from the intense boneconducted energy of the impact. The character and distribution of the anatomical changes in cats and of the hearing losses in man are practically identical with those resulting from blast or gunfire. Concern has frequently been expressed that hearing may be injured by the intense ultrasonic frequencies of the noise of jet engines and other sources. No positive evidence of any such injury has appeared (62). Such injury is very unlikely when the actual energies involved, the high acoustic impedance of the ear for ultrasonic frequencies, and the principle that injury occurs in general above the frequency of the injuring tone are all considered. The possible danger to humans from air-borne ultrasound must not be confused with the experimental, diagnostic or therapeutic use of focused, water-transmitted ultrasound directed toward the organ of Corti (40, 173). Intensities of about I watt per square centimeter at I megacycle frequency produce injuries that are comparable to those produced by electro-cautery applied to the cochlea (86, 225). Surgical Interference. In an effort to correlate hearing losses for particular tones with injury to particular regions of the cochlea, Neff, Schuknecht and their collaborators have carried out a brilliant series of studies on cats (167, 139, 207, 209, 225, 254, 21o), and Gross (xog) has performed similar experiments on guinea pigs. Local lesions were made in one ear. Sometimes the cochlea was entered surgically, sometimes it was cauterized, and in other experiments the auditory nerve was partially sectioned. The opposite ears were totally destroyed. The thresholds of hearing were measured before and after operation by behavioral tests, i.e., by the use of conditioned reflexes. These tests have been shown to be as reliable as or better than routine clinical audiometry on man. Ultimately the lesions and degenerations were studied under the microscope and their extent mapped by reconstruction from serial sections. Abrupt high-tone hearing loss was produced by lesions in the basal turn of the cochlea, and lesions extending to the middle of the cochlea caused considerable elevation of thresholds for tones of medium frequency. The relationships of high and low tones to base and apex of the cochlea were not symmetrical, however (254). Local lesions in the apex were more dficult technically to produce, and when produced Jawwy 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 9 they did not give abrupt or complete low-tone hearing losses. At one time it seemed that the low tones might have no corresponding location in the cochlea (167). It is now clear, however, that local surgical lesions of the apex of the cochlea do elevate the thresholds for low tones selectively (209). Partial section of the cochlear nerve may produce similar selective elevations of threshold for low as well as for high tones (210). The hearing losses from apical injury are never complete, however. The animals always respond to low tones if the tones are made strong enough. It seems that I> without the organ of Corti in the basal turn, high tones cannot be heard at all; 2) without the organ of Corti in the apical turn the sensitivity for low tones is reduced, but J) loud low tones can be heard somehow even if only the basal turn is intact. The lack of symmetry in these relationships was not predicted from the classical ‘resonance’ or ‘place’ theories of hearing (cf. 242) but it is in harmony with present interpretations of the dynamics of the cochlea. Experimental partial section of the cochlear nerve also has shown that over 75 per cent of the nerve fibers to a region of the cochlea must be lost before there is any elevation of threshold for corresponding tones (210). From clinical experience it appears, however, that although retention of a few nerve fibers may preserve a normal threshold many more fibers are needed to transmit effectively the complex neural signals of speech (210). The evidence from behavioral testing of cats relating frequency to place in the cochlea has been reinforced by the use of evoked cortical potentials as indicators. The tonotopic organization of the feline cortex is definite enough to make this a powerful method if the evoked potentials are ‘amplified’ by local applications of strychnine (234, 235, I 25). The place principle, including the unsymmetrical relationships of frequency to place as summarized above, is fully confirmed (139, 225). Against all of this clear evidence from animal experiments stands a very puzzling body of data collected by Dandy and recently summarized and discussed by Guild (112). For the relief of the symptoms of M&i&e’s syndrome, Dandy made partial sections of the eighth nerve in nearly a hundred patients. Audiograms were obtained on many of them following the operation, and Dandy had estimated how much and what part of the cochlear (auditory) nerve, in addition to the vestibular branch, he believed he had cut. When the nerve was totally severed hearing was lost completely, but for partial sections the hearing losses were always high-tone losses. The losses often included medium tones to an extent that correlated broadly with the amount of auditory nerve that was sacrificed. The point is that, short of total deafness but otherwise regardless of the extent of the cut, selective low-tone hearing losses simply did not appear. The general location in the nerve of the fibers to apex and to base is known, and it is incredible, if we read Dandy’s operative notes, that the fibers to the apex were &NZ~.S spared; yet low-tone hearing was not impaired. Guild states clearly the theoretical problem that is raised but offers no solution. The present reviewer regrets that no information is available as to the auditory discrimination of Dandy’s patients. Their ability to understand speech might have revealed an impairment other than elevation of the auditory threshold (2 10). One point that has confused the issue of the relation of frequency to place in the cochlea has, however, been clarified; namely, the failure of the electrical activity of the cochlea, as recorded from the round window, to give clear evidence of frequency localization. Frequency maps of the guinea pig cochlea had been successfully constructed on the basis of careful studies of small differences in output or in ‘threshold,’ (cf. 224), but in 1944 Lurie, Davis and Hawkins (160) concluded that the cochlear IO HALLOWELL DAVIS VoZwne 37 microphonic was a poor indicator of the extent and severity of acoustic trauma in the guinea pig and Smith (220) came to a similar conclusion. In their experiments on acoustic trauma, the electrical output was diminished by injury, but the relationships between the position and severity of the lesion and the elevation of ‘threshold’ for test tones of different frequency were very unsatisfactory. The confusion apparently grew from the assumptions I) that the place-frequency relationships within the cochlea were symmetrical and z) that the magnitude of the low-frequency cochlear microphonic recorded at the round window would be a valid index of the integrity of the organ of Corti near the apex. Actually, with electrodes on round window and neck, the cochlear microphonic generated at the apex is considerably attenuated and is weaker than the low-frequency response of the basal turn. A small segment of normal organ of Corti near the round window can give microphonics of all frequencies and at or near their ‘normal’ threshold. This situation has been explored thoroughly with pairs of intracochlear electrodes that record from only a small segment of the cochlea (56, 230) as will appear below. When the basal turn is depressed by surgical or acoustic trauma, the cochlea microphonic at the round window is depressed equally for all frequencies, because the responses are all coming from the same bit of tissue. Some of the best evidence for this generalization, although not interpreted in this way by the authors, is found in the papers of Wever and his collaborators (254, 259, who pointed out the independence of the frequency of the test tone in the effects of many agents that influence the cochlear microphonic. PROBLEMSOF AUDITORY PHYSIOLOGY The problems of the physiology of the inner ear can profitably be grouped around a series of questions which have been the core for many years of what has traditionally been called the ‘theory of hearing.’ We are not here concerned with the questions of how the acoustic energy is transmitted by the middle ear to the inner ear or of how the central nervous system analyzes and deals with the pattern of nerve impulses delivered to it by the auditory nerve. We start with the input of acoustic energy to the inner ear in the form of vibratory movements of the footplate of the stapes and stop with the transmission of impulses up the nerve. We will take it for granted that the movements of the stapes give rise to movements of the sensory structures of the organ of Corti and that the movements somehow set up the nerve impulses (180). We will ask, first, what is the gross pattern of movement of the sensory structures and how does it relate to the intensity and the frequency and time pattern of the sound waves. The sensory surface is obviously a long narrow ribbon, coiled in a spiral. Do its various segments move synchronously? Is there a maximum of amplitude of movement, and if so, does the position of movement shift lengthwise of this ribbon as the frequency is altered? Do the movements start and stop promptly with onset and cessation of the sound waves? And if answers to these questions are forthcoming, as they are, can we relate the answers to known physical principles and measured physical properties and make qualitative and quantitative predictions from them? To the extent that this can be done we can say that we ‘understand’ the dynamics of the cochlea. Then comes another major question: How does the mechanical movement of the sensory structures initiate nerve impulses? This leads to detailed questions concerning the minute anatomy of the sense organ and its innervation and also as to how the parts of the sense organ move relative to one another. Exactly what and where is the January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR II final mechanical movement that triggers the sequence of physicochemical processes that result in the excitation of a nerve impulse? Are one or more of the electrical potentials that can be measured in the stimulated cochlea involved in this trigger mechanism? How are these potentials generated? To what extent is mechanical (acoustic) energy transduced into the electrical or other energy that finally does the work of exciting the nerve fibers? This set of questions is of great interest to the neurophysiologist but traditionally has been almost entirely omitted from the ‘auditory theories’ of experimental psychologists for lack of any basis for answers. The physiologist asks his own favorite questions concerning adaptation, fatigue, dependence on circulation and oxygen supply, and possible regulatory influences, hormonal or nervous, that may modify the action of the sense organ. Finally comes the question of how the auditory information contained in the stimulus is coded in the space-time pattern of nerve impulses in the auditory nerve. To what extent does a mechanical analysis of sounds with respect to frequency in the cochlea encode the frequency information by exciting different nerve fibers? To what extent is frequency information transmitted unchanged as synchronized volleys of nerve impulses? To what extent must we look to neural interactions, either in the cochlea or in the central nervous system, to sharpen auditory discrimination with respect to intensity, to frequency and to simultaneity? These are the questions around which we shall build our descriptions and discussions in the following sections. DYNAMICS OF COCHLEARPARTITION Gross Movements of Cochlear Partition. Our knowledge of just how the basilar membrane, the organ of Corti and the tectorial membrane vibrate under the influence of sound is due almost entirely to the studies of one man, Georg von BekCsy. He has studied the acoustics and mechanics of the ear over a period of more than 25 years and has brought to bear his training and point of view as a physicist. An extremely useful summary of much of his work and ideas is to be found in Chapter 27 of the Handbook of Ex@rimental Psychology (27), and two others have appeared more recently (23, 25). For our description of the gross pattern movement of the cochlear partition, and also for many aspects of the fine movements relating to the sensory cells, we no longer need to make remote inferences and theoretical speculations (cf. 26). We can turn to Bekesy’s direct observations and measurements. They form a self-consistent body of information and the inferences from them have been confirmed at many points by the inferences of other investigators using somewhat different methods. Most of his observations were made on the cochlea of guinea pigs under the ordinary microscope or a slit lamp microscope (I: 2, 18, 21). In many cases stroboscopic illumination was employed and the structures under observation were made more visible by the use of intravital stain or by introducing tiny crystals of silver which settled on Reissner’s membrane, tectorial membrane, organ of Corti, and so on. Other observations involved very careful microdissection and still others probing with fine hairs and noting the patterns of distortion produced by given pressures. Mechanical frequency analysis: the place principle. Local static pressure, exerted by a hair applied near the helicotrema where the basilar membrane is broad, produces a circular deformation even when the pressure is not exerted at the center of the membrane. Because the deformation is circular, we infer that this most flexible part of the cochlear partition displays essentially the same elastic properties in the longi- 12 HALLOWELL DAVIS V&me 37 tudinal and in the transverse directions; although near the bony edge the partition displays considerable rigidity in the longitudinal direction. Near the stapes the deformed area is generally elliptical with its major axis parallel to the longitudinal axis of the cochlear canal. The deformations are broad and shallow, much as when a pencil point is pressed into the skin. From the flatness we can infer that the basilar membrane is not under tension. This is contrary to the theoretical assumption of many earlier writers who were undoubtedly influenced by the analogy with vibrating piano strings; but the conclusion is confirmed by making fine cuts in the basilar membrane, either transversely or longitudinally. The cut surfaces do not draw apart. Although there is no particular tension in the basilar membrane, there are marked dBerences in its stiffness from one position to another. These differences produce a continuous gradation in volume elasticity along the cochlear partition. The volume displacement, for a given hydrostatic pressure, at the widest part of the cochlear partition is two hundred times the volume displacement at a point three millimeters from the stapes. Furthermore, it is not Reissner’s membrane or the tectorial membrane but the basilar membrane that determines the movements of the cochlear partition. Bekesy set the stapes vibrating sinusoidally with constant amplitude and measured under the microscope as a function of frequency the displacements of various points along the cochlear partition (13, 14). He obtained, for human ears (cadavers) and for many species of animals, a series of curves relating relative amplitude to frequency. Each di8erent point on the membrane showed a maximum displacement for a particular frequency. In fact the behaviour of any point as a function of frequency resembled a resonance curve. In another experiment B6kesy cut into the cochleas of human cadavers at appropriate places and observed the vibration pattern of the entire partition for selected frequencies (13, 14). As the frequency of vibration was increased, the position of maximum response was displaced towards the stapes. No clear maximum was found for frequencies below 50 cycles per second and at IOO cycles per second it was still very close to the helicotrama, but at 1600 cycles per second the position was near the middle of the cochlea. Unfortunately, it proved impossible to measure directly the vibrations of the cochlear position for higher frequencies because the amplitudes were too small. When BCk&y plotted the distance from the stapes to the point of maximum displacement against frequency, the curve had the general form of the curves that had been inferred from psychophysiological data. There was little variability in the curves from different subjects. The structure of the cochlea showed great stability in this respect, in marked contrast to the elastic properties of certain parts of the middle ear, for instance. BCkesy has measured the positions of maximum amplitude as a function of frequency in many different kinds of animals from the elephant to the mouse (27). The curves look very much alike for all animals. He reckons the resolving power of a cochlea as the ratio of’ the shift of the point of maximum response along the partition for a given ratio of frequency change. In general the mechanical resolving power increases with the size of the animal. For higher frequencies and in most animals the resolving power is practically independent of frequency. The larger the animal the lower in frequency does mechanical resolving power extend. The differences in resolving power, BCkesy believes, are not due so much to differences in elastic constants as to differences in the effective mass that loads the cochlear partition. h?WWy 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 13 are heavily but not critically damped. decrement as I :3, and almost independent of frequency. In these experiments he introduced a tiny iron ball into the Scala vestibuli and lifted it abruptly by means of an external electromagnet. The movements of the cochlear partition were inferred from the cochlear microphonic, an electrical potential that is proportional to the displacement of the partition. This degree of damping is not very different from the damping that he measured long ago for the ear as a whole by recording optically the movements of drum or ossicles following a very abrupt acoustic stimulus. It is important theoretically that the damping is less than critical, because this allows the cochlea to operate as a mechanical analyzer by virtue of the principles of resonance. At the same time, the fairly heavy damping allows the cochlea to act as a rather good analyzer in the time dimension. There seems to be a good compromise here between frequency analysis and sharpness of discrimination in time. Such a compromise is a theoretical necessity because of the way in which frequency and time both involve the dimension of time (go). Tmvehg waves on the cochlear partition. Bekesy has measured phase differences, as well as amplitudes of displacements, among various structures and at various positions along the cochlear partition (I I), It was in these experiments that stroboscopic illumination and strewing tiny crystals of silver over the cochlear partition proved of particular value. He first showed that there were no significant phase differences between the basilar membrane, the organ of Corti and the tectorial membrane. Usually Reissner’s membrane also vibrated in phase with the other parts of the partition although at higher frequencies it sometimes vibrated in sections. He found, however, large and important phase differences between the movement of the stapes and the movements of various regions along the cochlear partition. Displacement is not simultaneous at all positions. Neither is there, as would be expected in a system of graded but independent resonators, a symmetrical lead in phase on one side and lag in phase on the other side of a point of maximum amplitude that moves in phase with the driving force. Instead, the movement of the cochlear partition lags more and more behind the movement of the stapes the farther from the oval window it is observed. This progressive lag amounts to at least r (‘opposite’ phase) at the point of maximum amplitude and to 2x, or one full cycle, a short distance beyond the maximum. As the waves shrink in amplitude, they lag more behind the movement of stapes, and they lag more and more rapidly as they move along the cochlea. Their wave length becomes shorter and shorter. Lags of as much as 3% have been seen by stroboscopic illumination (I I), and by electrical methods even larger differences have been demonstrated (230). The increasing phase lag at successive positions means that the position of maximum instantaneous displacement travels as a wave crest along the cochlear partition away from the stapes. The wave travels very rapidly at first and then more and more slowly. At first it increases gradually in amplitude as it travels. It goes through a rather broad maximum, and then rather abruptly it loses amplitude as well as velocity. This general pattern of a traveling wave, which is clearly visible under stroboscopic illumination, is the basic pattern of movement of the cochlear partition. The position of maximum amplitude of the envelope of the vibration pattern is a function of frequency, and the mean amplitude of displacement at any point is a function of intensity also. The approximate amount of phase shift at the point of maximum amplitude is independent of frequency and intensity. There is, or course, a lower frequency limit at which the point of maximum amplitude is at the apical end of the cochlear partition. Thus, below 50 cycles per second The vibrations of the cochlear partition B6k6sy (14) determined the value of the logarithmic J4 HALLOWELL DAVIS V&me 37 the movement of the entire cochlear partition is practically in phase, but above cycles per secondBekesy could always find the traveling wave. He writes (27): ISO In the vicinity of the region of maximum displacement amplitude, we observed traveling waves propagated toward the helicotrema. These waves are so damped that after one or two cycles they are no longer visible, but we can see them clearly enough to detect that the wavelength decreases markedly as they proceed toward the helicotrema. He also observed that the time required for the wave initiated by a sudden sound to reach the helicotrema is about 5 milliseconds. The pattern of traveling waves can be reproduced and studied in modelsof the cochlea. BekCsy made a model in which a very thin rubber membrane separated two fluid-filled channels (13). The membrane tapered systematically, like the cochlear partition, from one end to the other. The fluid in one channel was driven by a piston (stapes)and the other channel was provided with an elastic release(round window). When the piston is driven at an appropriate frequency, traveling waves can be seen (stroboscopically) on the membrane and two small eddies, one on each side of the membrane, appear just beyond the region of maximum amplitude of vibration. (If solid particles are suspendedin the fluid, the eddiesare clearly visible without stroboscopic illumination. Similar eddiesappear in the cochlea also.) The eddiesmove toward the narrower end of the membraneas the frequency is increasedand toward the broader end as the frequency is lowered. Parenthetically we note that theseeddieshave been invoked by somewriters to explain certain features of acoustic trauma (201) or of normal stimulation (178). In the model it was found that the formation and position of traveling waves and their eddies is, within wide limits, independent of the shape and dimensionsof the channelsand the viscosity of the fluid. More surprising, the formation and direction of travel of the waves are independent of the position of ‘stapes’and ‘round window.’ Both the driver and the elastic releasemay be at any position along their respective channels,but the waves always travel from the narrow, stiffer end of the membrane toward the broader, more flexible end. Thus when the ‘stapes’is placed at the broad end the waves travel toward the driver. B&k&y refers to this situation as “the paradoxical direction of wave travel along the cochlear partition” (24). He also notes that “this independence(of the location of the driving source) makes it clear why-an over-all vibration of the bony walls of the capsuleduring hearing by bone conduction producesthe samesensationas the transmissionof the vibrations through the stapes.” Paradoxical direction of travel of waves is not connned to models. If acoustic vibrations are introduced through a hole drilled in the apex, direction of the waves is from basetoward apex and the speedof travel is the sameas usual (230). Misunderstandingsof traveling waves.The term ‘traveling wave’ has led to considerable confusion and some controversy becauseit was not clear from BekCsy’s earliest statements that the term simply described the phase relationships of the cochlear pattern and did not imply anything as to the mechanism by which the pattern is created. Somefamiliar types of traveling waves, such as surface waves on water and shear waves in a beam or a rope, depend on the transfer of energy quite directly from one segmentof the water, beamor rope to the next, to which it is closely coupled. This is a different mechanismfrom the one assumedin all resonancetheories of hearing in which the force is transmitted through the surrounding medium and is applied directly to each vibrating element. (Here the coupling between elementshas traditionally been assumedto be small.) Another confusion was between the velocity January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR of sound (compression)waves in a fluid such asperilymph, of the order of a meter and a half per millisecond, and the velocity of the traveling waves, which varies from almost the velocity of the compressionwaves to a millimeter or lessper millisecond. For example: Wever and Lawrence (247) performed an experiment very similar to that of Tasaki, Davis and Legouix (230). Acoustic energy, at 5000 cycles per second,was applied simultaneously through the stapesand through a hole in the apex. The cochlear microphonic was measured,either at round window or at the apex, as the phase relation of the tone at the two positions was shifted. A cancellation (nul) could be obtained by adjusting both phaseand intensity, and the samesetting gave a nul for responsesat the apex as well as at the base. The authors concluded, like Bekesy, from these and other observations that stimulation of the cochlea by any channel, i.e., by oval window, by a hole in the apex, and by bone conduction, always produces the samepattern of vibration in the cochlea. But they did not consider the possibility of paradoxical direction of travel, which would be impossiblefor the type of mechanism which they believed was implied by the term ‘traveling wave.’ A similar argument wasmade again in a later paper (253). It is not necessaryto analyze the latter and point out in detail the errors of assumptionand interpretation because, fortunately, the semantic situation has now been clarified by a joint article by Wever, Lawrence and Bekesy (251) which states: Wever and Lawrence regarded their experiments, and other evidence as well, as out of harmony with a traveling-wave theory of cochlear action. By a ‘traveling wave’ is meant here a wave of transmission of vibratory energy along the basilar membrane. . . . Actually, of course, what is happening is that each element of the membrane is executing sinusoidal vibrations but the different elements are executing these vibrations in different phases. This action can be referred to as that of a traveling wave, provided that we mean simply to indicate that the amplitudes along the membrane are changing from moment to moment in the manner described, and nothing is implied about the underlying causes. It is in this sense that B&&y used the term ‘traveling wave’ in reference to his observation. It should be made clear that Wever and Lawrence, in their rejection of the traveling-wave theory, were not implying that there was anything at fault in B&&y’s observations of the cochlear action. Their position was that the observed traveling wave is to be taken simply as a temporal pattern of motion. . . . B&&y did not consider that his visual observations gave any decisive evidence on the paths of energy flow in the cochlea, and therefore he has not taken any position on this issue. The present reviewer is in full accord with this statement. With Tasaki and Legouix (230) he made explicitly the samesemantic point that the term ‘traveling wave’ wasused only in the senseof describing a sequenceof events in spaceand time. He believes that in what Wever, Lawrence and BCkesy call the ‘primary region,’ (see fig. I) up to and including the region of maximal amplitude, the transfer of energy is primarily through the fluid. In the ‘secondary zone,’ where the decrement of amplitude and increase of phase lag are both rapid, the transfer of energy along the basilar membrane seemsto become of greater relative importance and can no longer be neglected. But the small movements of the cochlear partition in this ‘secondary zone’ beyond the maximum are probably of little or no physiological significance (cf. also 24). But with this episodein mind the present reviewer, when lecturing about the pattern and the dynamics of cochlear movements, refrains from shaking a rope to illustrate a traveling wave and he avoids motion pictures that make an analogy with waves of the searunning up a sloping beach (232). 16 HALLOWELL Primary zone DAVIS Vohme 37 Secondary zone Distance along the cochlea Pattern of traveling waves on the cochlear partition. The stapes is at the left, the helicotrema at the right. The dotted curve, b, represents the displacement of the partition a moment later than the solid curve, tz. The wave length rapidly becomes shorter as the traveling wave enters the secondary zone beyond the position of maximum displacement (from ref. 251, by permission). FIG. I. Resonanceand traveling waves.BekCsy has continued to investigate, in a series of electrical and mechanical models, the necessary conditions for the production of traveling waves in a vibrating system and for the paradoxical direction of travel (24). His attention has been directed to systems that might show maxima of displacement as a function of frequency and thus serve as frequency analyzers. In particular, he shows how traveling waves are produced in a series of graded resonators when vibratory energy is applied to one or to all of them simultaneously. The variables that he manipulates are chiefly the stiffness of the resonators and the coupling between them. Both elastic coupling and friction-mass coupling are employed. The former is realized by rubber bands woven among the steel tongues of the vibrating ‘reeds,’ the latter by partially immersing the vibrators in water. BekCsy believes, incidentally, that the coupling along the cochlear partition is partly elastic and partly friction-mass. In the model various degrees of damping were also introduced. It becomes very clear in the course of Bekesy’s analysis that in a system with graded tuning of a series of elements that are coupled to a significant degree, or of equally tuned resonators with a graded degree of coupling, traveling waves are the rule, not the exception. Furthermore, the direction of travel is regularly from the stSfer toward the more flexible part of the system. “In both the model and the living cochlea, it seems that the stiffer part of the system drives the softer part.” An important condition for such ‘driving’ and the possibility of paradoxical direction of travel is that in such a system the wave length drops very abruptly to a small value. When this occurs, the section at one end of the system is completely under the control of the driving forces, whereas the other section is free in spite of the large driving forces because these cancel each other out (to the extent that they act on a full cycle or an integral number of cycles of traveling wave in a short distance). Another condition for paradoxical direction of travel is that the driving force be applied to all or to a considerable part of the length of the system, not merely to a single element. In a still later paper (25) BekCsy shows the continuity between four types of vibration patterns. These patterns correspond actually to four major classes of ‘theory of hearing.’ The patterns, for steady tones, are: 1) Simple resonance: movement in opposite phase on opposite sides of a maximum of amplitude. Phase varies continuously from r/2 to -r/2. (Helmholtz). A single maximum of amplitude. z) Movement of membrane as a whole without phase differences, like a telephone diaphragm (Rutherford). Not more than one maximum of amplitude. January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR I7 3) Traveling waves. Continuously increasing phase lag (BekCsy). One maximum of amplitude. 4) Standing waves. Successive regions of opposite phase. Abrupt discontinuities in phase (Ewald). Multiple maxima of amplitude. The transitions among these four classes or patterns are made by varying either a) the absolute stiffness of the membrane, b) the amount of coupling between adjacent parts, or c) both. A constant gradation of stiffness from end to end of the ‘membrane’ is assumed. A moderate stiffness, with fluid coupling between elements, gives the single classical stationary resonance pattern. The same slope of stiffness and the same driving frequency but with greater absolute stiffness gives vibration in phase throughout like a telephone membrane. Or a resonance type of pattern becomes a telephone type if the frequency is lowered below the resonant frequency of the lowest-tuned resonator. This is the only type of response possible at very low frequencies. Reduction in stiffness of the membrane, on the other hand, gives the traveling wave pattern with waves moving from the stiffer toward the more flexible region. Still further reduction in stiffness gives greater amplitudes, shorter waves that travel farther, until finally the waves are reflected from the end of the membrane and a standing wave pattern is formed. There seems to be no longer any doubt but that in the living cochlea the absolute values of stiffness and coupling and their gradations along the cochlear partition are such as to yield traveling waves over nearly the entire range of audible frequencies. This is not a ‘simple resonance theory’ of cochlear action, but it is rather a modified or extended resonance theory. So far all statements about patterns of vibration have been con6ned to steady states. Bekesy has not yet fully investigated the responses of his models to transients, but he does make the important observation that, with normal damping, the model that shows the stationary resonance pattern in the steady state shows a traveling wave in response to transient excitation. Otlzer tI&ories of co&ear dynamics. A puzzling set of observations, for which no clear explanation is now available, are those of Pumphrey and Gold (176, 103, 102) which led them to the conclusion that the cochlear analyzer is very sharply tuned. As a corollary, the damping is inferred to be very low. The observations turn on pitch discrimination and the recognition of transients near the upper end of the audible range. We can only offer the lame suggestion that some unrecognized clue or artefact assisted the subjects in making their discriminations. The authors’ theoretical conclusion that the cochlear partition is lightly damped is untenable in the light of Bekesy’s direct measurements (14) and the presence of traveling waves, even at IO,OOO cycles per second, in the guinea pig cochlea (230). Another theory of the mechanics of the organ of Corti, based on direct anatomical observations, has been advanced by Hilding (123, 124). His observations were confined to simple microdissection without stroboscopic illumination or electrical recording. His speculations as to the probable movements of the organ of Corti and the tectorial membrane are in part correct and in part erroneous. In Meyer’s writings, (162, 162a) experimental observations are so mixed with polemics and a very personal vocabulary that analysis of his ‘hydraulic’ theory in terms similar to BCkesy’s and Wever’s is difficult. This theory now has chiefly historical interest. Like Hilding’s, it seems to violate the principle (so) that, since stimulation of the ear near threshold occurs at energy levels near the minimum that 18 HALLOWELL DAVIS Volume 37 is physically possible(238, 50, 27), the transfer of this energy to the point of actual stimulation must be extremely efficient. More important, both theories are contrary at crucial points to B6kesy’s actual observations. Cochlea Models and Pitch Discrimination. The patterns of movement in cochlear models,and in actual ears, seemwoefully inadequate to explain the known facts of human and animal pitch discrimination (49). The place principle has been established, but the maxima of amplitude seemmuch too flat. The flatness is even more evident when the cochlear microphonic is taken as the measure of activity (230). The frequency principle is not disproved by the establishment of the place principle, however, and actually for medium and low tones the cochlear partition of the basal turn of the cochleamoves in phaselike a telephone diaphragm in a way ideally suited to set up synchronized volleys of impulses. BekCsy and Rosenblith (27) say: We are at present far from a definite solution of the problem of pitch discrimination. What was attempted here was to demonstrate the possibilities of a purely mechanical frequency analysis in the ear. It was never really possible to throw the entire burden of pitch discrimination on mechanical analysis alone because the mechanical excitation falls off much too slowly on both sides of the maximum (i.e., the resonance curve of pure tones is much too flat). It is still not clear by what process the nervous system increases the apparent sharpness. . . . The positions that have been stimulated less than maximally are apparently subject to some sort of nervous inhibition in the manner of the law of contrast. In any case the fact that mechanical time constants are very small compared with over-all auditory time constants as found in psychophysical experiments must eventually be fitted into the picture of a frequency-and-timeanalyzing system for which equivalent linear circuits with simple time constants and resonance curves cannot be readily established. One suggestionas to how the nervous system ‘sharpensthe pattern’ is that the place principle operates not on the maximum of amplitude of displacement but on someother feature of the spatial pattern. Shearing forces and longitudinal displacements are proportional to the first derivative of the traveling wave as well as to its vertical amplitude. The amplitude of the first derivative of the traveling wave has a sharper maximum than the wave itself and the maximum is displaced toward the helicotrema. The secondderivative is still sharper (132). But this fact is little help unlessneural mechanismsare invoked to determine the derivatives. Phasedifferences may also provide useful information. Huggins (130, 131) makes certain postulates, such as the stimulation of external hair cells by longitudinal movements and an inhibitory action in the cochlear nucleus of the impulsesso excited, and he develops the consequencesin a mathematically oriented theory. His theoretical treatment indicates that the kind of neural mechanismthat is postulated could do a pretty good job, but this doesnot prove that the assumptions,ingeniousas they are, are correct (cf. 249). A different approach has been to construct a cochlear model that is provided with a human nerve supply. Bekesy (25) used the skin of the forearm as the sensory surface. He applied the vibratory stimulus through one of three modelsthat provided the telephone type, the resonance type and the traveling wave type of wave patterns respectively. The resonating model consisted of a series of tuned steel reeds attached to a metal support that oscillated slightly around its longitudinal axis. The length of the support was equal to the length of the forearm. Thirty-six reeds, tuned in equal intervals over a range of two oc- January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 19 taves, were distributed along the whole length. A small pin on each reed, fastened close to the support, touched the surface of the skin of the arm. The pins had rather small points, so that the skin would not pick up too much energy from the vibrating reeds; otherwise it would not have been possible to obtain a sharp resonance of the reeds. When the arm is placed carefully along the pins, so that the points just touch the skin, the reed system transforms any change in frequency into an easily observable displacement of the stimulated area on the skin. The model for the telephone theory is a triangular metal frame made of tubes. The frame vibrates perpendicularly to the axis of the edge in contact with the forearm. The rigidity of the frame insured that all the stimulating parts touching the skin would vibrate in phase. The model for the traveling waves was a section of a model of the human cochlea, enlarged by dimensional analysis. The frequency range was two octaves. The model was a plastic tube cast around a brass tube with a slit. The tube was fUed with fluid. The forearm lies in contact with the vibrating membrane. A vibrating piston sets the fluid inside the tube in motion, and forces in the fluid produce waves that travel from the hand to the elbow, The traveling waves thus produced are similar to those observed in preparations of human cochlea. The maximum amplitude of vibration is rather broad, and when it is observed under stroboscopic illumination, it moves along the membrane as the frequency is changed. Although the maximum is quite flat as it moves along the arm, the sensation of vibration is concentrated on a relatively short length (about 2 to 4 centimeters); hence, any frequency change is easily recognized by a shift in the stimulated area. Seemingly the nerve network in the skin inhibits all the sensation to either side of the maximum of the vibration amplitude, thereby producing a sharpening of the stimulated area. If we compare the three models, we find that the difference limen for ‘pitch’ discrimination below 40 cycles per second is the same because the skin is able to discriminate the roughness of the vibrations as such. But for higher frequencies, displacements of the sensation along the arm, produced either by the resonating model or the traveling-wave model, permit much more accurate frequency discrimination than the telephone-theory model does. . . . Even when the stimulus was only two cycles, the pitch discrimination for both the resonance model and the traveling-wave model was just as good as it was for a continuous tone of longer duration. Closer examination showed that in both models the place of maximal amplitude was determined during the first two cycles of the onset of a tone. . . . The surprising fact is the inhibitory action of the nerve supply, which suppresses all sensation except on a small spot near the maximal amplitude of vibration. The reviewer finds it most encouraging that this kind of inhibitory sharpening of the locus of stimulation has been directly demonstrated in another, but closely related, sensory modality, although the mechanismof the inhibitory action is not yet disclosed.The ideas that were vaguely sought earlier by BbkCsy in his ‘law of contrast’ and by Gray in his ‘principle of maximum stimulation’ now begin to take reasonableform and tangible reality in spite of their disheartening complexity. The advance from Wever’s (242) unsympathetic discussion of Gray’s and Bekesy’s principles is a major one. At the sametime, as we shall see, the place principle and the frequency principle are now synthesized into a ‘duplex’ theory of hearing. Mathematical Theories of Co&ear Dynamics. The publication between 1940 and 1950by BekCsy of his numerousmeasurementson the middle and the inner ear stimulated a seriesof mathematical theories to account for and unify the observations (147, 184, 190, 171, 31, 87, 88, 179, 263, 264, 266). Zwislocki has written a very illuminating review (265) of these theories from which we condense the following comments. In the recent development of the theory of co&ear dynamics four trends can be distinguished. Two of them are not new. One follows more or less directly the resonance theory of 20 HALLOWELL DAVIS Volume 37 Helmholtz, and the second the standing-wave experiments of Ewald. The third trend puts the emphasis on the hydrodynamical phenomena within the co&ear fluid and ascribes a decisive role to the relation between the depth of the canal and the length of waves propagated along the cochlea. Finally, the fourth stresses the damping of the cochlear duct and ascribes the occurrence of a local maximum of vibration to the interaction between this damping and the elasticity of the cochlear duct, thus completely excluding any kind of resonance. From the mechanical point of view, the actual nature of the cochlear partition can most probably be neglected without committing too great an error and considered as a simple elastic wall. Another simplification which seems permissible is to straighten the canal, which actually has the form of a spiral. If no extensive simplifications were necessary for the mathematical treatment of cochlear dynamics, it would be sticient to find a solution to the equations of continuity and equilibrium of forces that would satisfy the boundary conditions, i.e., the conditions at the rigid walls, at the helicotrema, at the windows, and at the partition. Unfortunately, boundary problems are very dticult, and as yet no exact solution has been found for the dynamic behavior of the cochlea. If only an approximate solution can be found and simplifications cannot be avoided, numerical data are needed in order to see what is important and what can be neglected. The geometrical and physical constants which can be regarded now as fairly well established are the length of the canal and its cross-sectional areas, the dimensions and impedance of both windows, the dimensions of the helicotrema, and the density and viscosity of the perilymph. There is more uncertainty as to the components of the impedance of the cochlear partition, but the elasticity and mass per unit length may be considered to be known to a sufficient degree. The factor the least well established is the damping of the partition. Its order of magnitude can be deduced, however, from Bt?k&y’s observations of the decay of vibration. Zwislocki analyzes the several theories according to their mathematical approaches.A significant point to the present reviewer is that two of them, namely the theories of Kucharski (147) and of Reboul(184), can now be set asidedefinitely becausethey yield patterns of vibration that are clearly contrary to the observations of BCkCsy.The other theories all predict a region of maximal excursion whoseposition depends on the frequency and also a pattern of traveling waves. The theories of Peterson and Bogert (171) and of Bogert (31) are quite similar to the later theory of Fletcher (87, 88). All three of them are resonancetheories; they attribute the maximum of vibration to the resonanceof the cochlear partition. Zwislocki (265) notes that I‘. . . in his most recent theory Ranke (179) seems to abandon his earlier idea of a strong wave reflection at the place where the waves have ceasedto be long in comparisonwith the cross-sectionaldimensionsof the cochlea. . . . Ranke’s theory is a ‘short-wave’ theory and is subject to the same objection as are the ‘long-wave’ theories: it is only an approximation. . . . The question of which approach comes nearer to the actual situation-long waves or short ones-cannot yet be decided.” Zwislocki’s own theory (263, 264) was the first to be basedon Bekesy’s measurements of elasticity and damping of the cochlear partition. An important fact that emergedfrom it is the mathematical evidence of the possibility of frequency analysis without reliance on resonance.In his latest paper (266) he further shows that the pattern of responseof the cochlear partition must be constant regardless of the channel by which the acoustic energy reaches the cochlea. In other words, the paradoxical direction of travel of the waves is a theoretical necessity. Zwislocki (265) further states: January x957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 31 In one respect or another the theories disagree among themselves on numerical values and no theory is in complete accord quantitatively with the experimental results. For example, . . . because of different values taken for the impedance of the cochlear partition, Fletcher’s calculations seem to agree more closely with the observations of B&&y than do those of Peterson and Bogert. On the other hand, . . . nearly all recent theories state that, for frequencies above about IOO cps, waves shorter than the co&ear canal are propagated from the oval window towards the helicotrema. At a certain point, dependent upon the sound frequency, they reach a maximum, At least in the portion of the cochlea between the windows and the maximum of vibration, the wave length decreases steadily. This occurs independently of what assumption is made concerning the factors responsible for the maximum of vibration. Thus, whether resonance is postulated or not, approximately the same pattern of vibration results. And finally, The analysis of the individual theories indicates a certain convergence of opinions, and it is clear that definite progress has been achieved toward an understanding of the dynamic behavior of the cochlea. In spite of this trend, however, no one of the theories seems entirely satisfactory. Local Movements in the Cochlear Partition. The pattern of local movement within the cochlear partition has been described most fully by BCkCsy (21-23). He was seeking, of course, the details of mechanical movement which might indicate where the critical events of the excitatory processoccur and what particular mechanical force, displacementor strain might causeit. He employed the optical and acoustic methods already mentioned and also recorded local electrical potential changes (cochlear microphonics). Particularly ingenious and effective was his use of a tiny vibrating electrode that served simultaneously as stimulator and electrical probe. Summarizing BCkesy’sdescriptions, we can say first that the tectorial membrane doesnot show great stiffnesswhen probed by hairs and it may easily be moved slowly perpendicular to the basilar membrane “like the cover of a book hinged at the modi01~s.”The elastic forces of the hinge are small so that the return time after displacement is more than a second. The tectorial membrane is attached to the reticular lamina of the organ of Corti; but can be torn free. “It can be split easily along its fibers which form an angle of about 60 degreeswith its edge.” The membrane itself is soft, but it has great internal friction and seemsquite stiff and rigid when touched with a needlevibrating at 200 cycles per second.The inner consistency is something like that of “black pitch at a certain temperature.” The particular importance of this partial rigidity seemsto be that when the organ of Corti vibrates, it tends to move relative to the tectorial membrane. The reticular lamina, or ‘membrane’ asBekCsy calls it, is a stiff perforated plate in which the hair cells are fitted at one end. “The other end of the hair cells hangs down practically free.” Of course, the reticular lamina is rigidly attached to the basilar membraneby the two pillars of Corti. Unlike the tectorial membraneand the basilar membrane, it is easily perforated by needle tips smaller than 5 microns. It seemsto be mechanically like a stiff net lying on a soft cushion with considerable stignessagainst movements along the surface of the membrane. HALLOWELL DAVIS Vobne 37 F’IG. 2. Diagrammatic drawing of a cross section of the cochlear duct in the second turn of the guinea pig cochlea. Most of the structures mentioned in the text are labeled (from ref. 56, by permission). Bekesy points out that the high sensitivity of the ear requires that the energy transmissionbe very efficient. (cf. also so). This means that the mechanical impedancesmust be well matched. The eardrum and the ossiclesmatch the impedance of the air to the mechanical impedanceof the vibrating fluid columnsin the perilymph and the elasticity of the cochlear partition. In order to pick up the energy of these fluid movements and transfer it to the stiffer elastic tissuesa secondmatch of impedances is needed to transform the pressure acting on the basilar membrane into a larger force. One way of making such a match is to transform a pressureon an elastic membraneinto a shearingforce. The tectorial membrane is thin and unelastic. It lies on a thick elastic plate which has a relatively stiff upper surface, the reticular lamina. The cochlear partition bends both transversely and longitudinally (as described earlier), and this must set up shearing forces between the tectorial membrane and the reticular lamina and also between the reticular lamina and the basilar membrane. But Bekesy writes, “which is important is for the moment unknown to me.” A shearing action between tectorial membrane and reticular lamina must obviously bend the hairs of the hair cells becausethe hairs are attached to both of them. BCkesy (23) gives good reasonsfor discounting any physiological importance of the small short waves on the basilar membranebeyond the region of maximal amplitude. First he notes that the stiffness that determines the pattern of vibration is in the basilar membrane. The organ of Corti is a soft, thick, elastic sheet lying on this stiffer membrane. “If such a combined sheet is bent in long waves the surface of the elastic sheet will follow precisely the wave-form of the stiff membrane. But if the wave-length is short compared with the th icknessof the soft sheet, the surface of the soft sheet will not show any deformation. . . Therefore, if the stimulation . . . occurs Janzcary rg,p BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 23 helicatrema t FIG. 3. Distribution of radial and longitudinal vibration along the organ of Corti for stimulation with a tone, seen through Reissner’s membrane (from ref. 21, by permission). at a certain height above the basilar membrane, perhaps immediately under the tectorial membrane, the small (short) longitudinal waves of the basilar membrane have very limited physiological importance.” Bekesy saw in the organ of Corti both longitudinal and transverse vibrations relative to the basilar membrane. He writes “near the place of maximal transverse vibrations . . . on the side (toward) the stapes the ends of the hair cells under the tectorial membrane and the Hensen cells perform vibrations in a direction which is close to the direction of the fibers of the tectorial membrane . . . . Going toward the helicotrema, this type of vibration stops quite suddenly, and the inner and outer hair cells show only an up-and-down movement. Further in the direction of the helicotrema, the vertical vibrations pass(into) longitudinal vibrations, the amplitude goes through a peak, and then everything quiets down. . . . At the moment, I do not know which of the three sections stimulates or perhaps inhibits the auditory nerves.” ELECTRICAL PHENOMEBAOF TEE COCEILEA Four classesof electrical phenomenahave been identified in the cochlea. One is the action potentials associatedwith nerve impulses. A second class is resting DC polarizations, notably the positive endolymphatic potential, and also the familiar negative intracellular potential that is common to all excitable cells and to many others aswell. The other two are responsesto acoustic stimulation: one, an alternating current responseknown variously as the aural microphonic or (better) the cochlear microphonic; the other a change, positive or negative, in the magnitude of the endolymphatic potential. This fourth classhas been designated the summating potentials. For conveniencewe shall refer to them as AP (action potential), EP (endolymphatic potential), CM (cochlear microphonic), and SP (summating potential). The term ‘cochlear microphonic’ now seemspreferable to ‘aural microphonic’ advocated by Stevens and Davis (224) in order to distinguish explicitly the ‘microphonics’ of cochlear origin from those originating elsewherein the inner ear. Analogous ‘ampullar microphonics’ are generated in the nonauditory labyrinth. Special Recording Techniques. The CM, AP and SP responsescan all be demonstrated with extracochlear electrodes. The classicalleads, still employed almost exclusively by someinvestigators (250, 253) are from the round window, or sometimes the apex, and a remote referencepoint, usually on the neck. Sometimesthe reference point is selectedto minimize one type of responsewhile retaining another (194). Such extracochlear electrodes are simple and the cochlea is completely uninjured; and if 24 HALLOWELL DAVIS Voturne 37 the object is to use CM to study transmission properties of the middle ear (249, I&, IOI) or the overall action potential (Igb), they are ideal. They are in general quite inadequate, however, for analyzing intracochlear events because they record a mixture, in uncertain proportions, of AP, CM and SP and also a mixture of responses from various parts of the cochlea. This is a serious limitation becauselow tones activate all or nearly all of the cochlea while high tones activate only a small part. Furthermore, there are large phase differences between diff&ent segments of the cochlea ( ISI, I I, 230). We have already noted the difEculty of assessing acoustic trauma from changes in the cochlear microphonics at the round window. Successive refinements in electrodes have been, first, the introduction of single fine wires into one or more turns of the cochlea (61,6o, 188); then the placement of a pair of electrodes opposite one another in a single turn, one in Scala vestibuli, the other in Scala tympani; then two pairs of such ‘differential’ electrodes in two turns simultaneously (230) ; then the introduction of a pipette electrode into Scala media (19, 20, 22g), and finally the use of hyperfine pipette electrodes capable of recording intracellular potentials (159, 229). Further elaborations have been the vibrating electrode of microscopic dimensions for localized mechanical stimulation and recording (16) and the use of the pipette electrode in Scala media for injections of chemical reagents (65, $3). The ‘differential’ electrodes have the advantage of leaving the sensory structures in Scala media practically intact and giving a reasonable restriction of the length of cochlear partition from which potential differences across the organ of Corti are chiefly recorded. This distance is of the order of 2 or 3 millimeters for the cochlear microphonics because both Reissner’s membrane and the organ of Corti have considerable electrical resistance (IS). These barriers blur the fine pattern of potential distribution. Also if the local responses are much depressed these electrodes will show the weak electric fields produced by the response of more distant segments. Only with very fine pipettes introduced into the cochlear partition, preferably under optical control, can the tie pattern of electric response be adequately explored (20, 22, but cf. also 227a). With differential electrodes, however, it is easy, with a suitable input network, to record separately the gross CM across the partition without contamination by action potentials and, on another oscilloscope, the potential between this turn of the cochlea and the head as a whole. This latter combination rejects the CM and SP and records the gross action potential of the auditory nerve as volleys of impulses pass through the modiolus (230, 227). Single nerve impulses are unfortunately masked by the electrical ‘noise’ of tonic activity of neck muscles, etc. and can only be isolated by hyperfine pipette electrodes introduced into individual axons. Distinctive Features of AP, EP 9 CM and SP. AP. The characteristics of AP are those of axon ‘spikes’ of other medullated nerves. In individual fibers the responses are all-or-none and each is followed by the usual refractory period. In the over-all response of the auditory nerve most of the impulses in response to tones below 2000 cycles per second are approximately synchronized in clear volleys. Unfiltered clicks elicit particularly large, well synchronized volleys. Particularly characteristic is ‘masking,’ i.e., the reduction or obliteration of a response to one signal by simultaneous stimulation with another tone or noise. CM. The cochlear microphonic is an alternating electrical potential that is proportional, up to a limit, to the intensity of the acoustic stimulus. The response is continuously graded and there is no true threshold and no refractory period. The output reaches a maximum, rarely over about 5 millivolts (measured in Scala media), and Jamary 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 25 further increasein the stimulus may causea depressionof the CM. At low and moderare intensities CM apparently reflects accurately and without measureablephase difference the instantaneous amplitude of displacement of the cochlear partition (22), and more specifically the displacement of the tectorial membrane relative to the reticular lamina. It dependsupon the integrity of the hair cells. CM is closely dependent on an adequate oxygen supply, although a relatively small fraction seems to be anaerobic and persistsfor hours after death. (244, 252). It is generally agreed that this residue represents a second type of cochlear microphonic, which we will designate ‘CM; in contrast to the oxygen-dependent ‘CMI.’ Unlessotherwise noted ‘CM’ will refer to an unspecified mixture of CM1 and CM2, usually strongly don& nated by CMl. CM doesnot show adaptation or any evidence of fatigue, other than a hysteresiseffect following supramaximal stimulation (224). DC. The familiar intracellular potential of most if not all cellsis 20-80 millivolts negative relative to the surrounding tissue fluid (159). Such intracellular potentials have been demonstrated in the cochlea when hyperfine electrodeshave been inserted into nerve fibers, hair cells, and other cells of the organ of Corti, stria vascularis and even Reissner’smembrane (19, 20, 229, 227). EP. The endolymphatic potential2 is an extracellular polarization. The entire fluid-filled spacein Scalamedia, including the tectorial membrane, measuresabout 80 millivolts positive with respect to Scalatympani, Scala vestibuli or the tissuesof the head. The extent of EP is indicated in figure 4. EP, like CM, is directly and immedi~ ately dependent on an adequate oxygen supply. It can be increasedor decreasedby static displacementof the cochlear partition (19, 229, $3). SP. The fourth classof cochlear potentials, the summating potentials (SP), are changesin EP in responseto acoustic or mechanical stimulation (54, $3, 65). The changein EP is proportional to the intensity of acoustic stimulation, but unlike CM, it is a unidirectional changeand is proportional to RMS (root mean square) acoustic pressure (a running time integral) and not to the instantaneous displacement. SP appears as a shift in the baseline on which CM is superimposed.It is very easily demonstrated with an electrode on the round window by using brief high-frequency tone bursts with rise time of about I millisecond as stimuli (170). It behaves as if the alternating acoustic pressurehad been rectified and ‘detected,’ but it is not merely an unsymmetrical nonlinear distortion or ‘peak-clipping’ of CM. SP showsno latency and it hasno true threshold. For a given small voltage SP requires a stronger stimulus than CM but may increase to as much as IO millivolts, larger than CM. It does not reach a maximum below levels which causeanatomical injury. It can sometimes be seenin the range where the CM responseis still linear but is rarely prominent below the range of nonlinearity. SP is typically increasedby moderate anoxia (104) and by mild chemical injury (58, 65). Furthermore, SP may be either an increaseor a decreaseof EP. To explain this paradox we now recognize two summating potentials, SP+ and SP- (54, 58, 104). Anatomical Loci of Origin of Potentials. AP. AP is, of course, the sum of many all-or-none spikes generated in and by the nerve fibers. No all-or-none responseor ‘propagated disturbance’ has been describedfor the hair cells or any structure other than nerve. The modiolus is the effective locus of recording of AP with the usual * Since this was written Tasaki (227a) and also Smith, Deatherage, Gessert and Davis have found that the potential in the endolymph of the utricle is not more than +5 mv. It will be better in the future, therefore, to use the more restricted term ‘endocochlear potential’ instead of ‘endolymphatic potential.’ 26 HALLOWELL DAVIS Vohze 37 FIG. 4. The ‘endolymphatic space’ is shown in heavy outline. Only one external hair cell is drawn, the tectorial membrane is omitted entirely and the rods of Corti are barely indicated. Only within the endolymphatic space do we find the positive endolymphatic potential of 80 mv. Parts of Scala vestibuli may be as much as 5 mv positive relative to scala tympani. The negative intracellular potentials vary from 20 to 60 mv, depending probably on the degree of injury by the microelectrode (from ref. 229, by permission). electrode placements (227). (Both the spiral ganglion and the internal auditory meatushave been suggestedpreviously as the effective point of recording, but without direct evidence.) An early manifestation of AP, ascribedto the non-medullated nerve fibers in the organ of Corti, can be seenwith an electrode deep in Scala tympani or within the modiolus (64). This early registration of AP was at one time confusedwith sp (188, 59). El? The sourceof EP is not known, but it must lie somewherein the wall of the endolymphatic space.The stria vascularis has been suggested(so, 52) on the very general grounds of proximity to blood supply, but the observed electrical gradients in the spiral ligament and in the stria itself did not agree with the -predictions from this hypothesis (229). CM. The source of CM is clearly in the hair cells. This was adequately established many years ago (224). Recently Bekesy (19, 20) and Tasaki (229) have both found, with hyperfine electrodes, that CM appears strongly inside of some cells (presumably the hair cells) but not in all cells of the organ of Corti. There is an abrupt phase reversal of CM just as the electrode enters the positively charged endolymphatic space(2 29) . This locates the ‘source’ of CM1 at the level of the reticular farnina where the hairs are located. The locus of origin of CM1 and also its relation to movements of the tectorial membrane have been most fully described by BCkCsy (16-18, 20, 22, 23). He measured the cochlear microphonics produced by shearing vibrations near the inner and outer hair cells. The vibrations were produced by a laterally vibrating needleplaced on the tectorial membrane. He concludes that the microphonics are-produced by a January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 27 ‘generator’ located between tectorial membrane and the basilar membrane. It is the shearingforces, not the pull, that produce the microphonic. He summarizesas follows ( 22 ) : The size of these microphonics changes according to the direction of the lateral movements and the place on the tectorial membrane. On the outer edge of the tectorial membrane the microphonics went through a maximum when the direction of the vibrations lay in the direction of the fibers of the tectorial membrane. Perpendicular to this direction the microphonics went through a minimum. On the inner edge of the tectorial membrane the whole direction pattern is turned around counterclockwise somewhat less than go degrees. There are two sets of microphonic generators very close to the inner and outer hair cells. The inner set is more sensitive to lack of oxygen. These two sets may be able to discriminate between the radial and longitudinal vibrations of the organ of Corti, a change that occurs near the place of maximal stimulation in the cochlea exposed to a tone. No localization has been attempted for CMZ, the anaerobic post-mortem fraction. There may be several sourcesfor this classof CM, wherever a moving structure carries an electrical charge. No difference in phasebetween CM1 and CM2 has been described! SP. The source of SP is not known, but its close relation to mechanical movement clearly places it in the movable part of the cochlear partition rather than in stria vascularis or limbus. It seemsto be a changein EP, but it may have a different sourceand simply add algebraically to the resting endolymphatic potential. Perhaps SP+ and SP- are generated in slightly different areas, like the CM from internal and from external hair cells, and add algebraically with each other. Or there may be a singlesourceof SP the polarity of which may depend on the resultant of unsymmetrical but opposing mechanical forces which finally deform the generator in one direction or the other. When a microelectrode penetrates into the endolymphatic space, SP increases much more than CM. This does not prove that SP and CM have different sources or are daerently oriented, becausethe faster alternating current (CM) passesmore readily through the capacitative pathways acrossReissner’smembrane, stria vascularis, organ of Corti, etc. The rate of potential changein SP is always slowerthan that of the accompanying CM becauseit corresponds to the envelope of the acoustic stimulus while CM reproducesthe carrier wave. The simplest hypothesis is to ascribe SP, as well as CM, to the hair cells. The Problem of Maximum CM Voltage The familiar relation of CM to acoustic pressurehasbeen abundantly confirmed. The output voltage is directly proportional to the input pressureover a wide range but it finally becomesnonlinear, levels off and goesthrough a maximum. The maximum is rather flat, for medium and low tones. Two measuresdescribe this function adequately: one is a pseudo-threshold, i.e. the intensity at which some arbitrary small voltage within the linear range is reached; the other is the maximum voltage. These two parameters may vary together or quite independently. For example, in poisoning with dihydrostreptomycin the threshold is not much altered but the maximum voltage is greatly reduced (I 18, 120). The sameconcept has been useful in describing and analyzing the effects of exposure to very intense sound (56, 41, 233, 8 Another possible interpretation of the ‘anaerobic’ CM2 has been suggested to the reviewer by Dr. Rempton Wing. The post-mortem CM2 may be primarily or entirely a reduced CM1 that is sustained by oxygen dBusing in from the middle ear through the round window. 28 HALLOWELL DAVIS Vohme 37 246, 249). Injury sticient to cause clear intracellular injury to hair cells without disrupting them or other structures causes a rise in pseudo-threshold of CM of about 40 decibels but a reduction of only 20 decibels in the maximum CM voltage (56). The maximum voltage is greater for low frequencies than for high. Near the extreme upper limit (guinea pig) the maximum is only a few microvolts and it is reached only a few decibels above the pseudo-threshold (I 70), while for low frequencies apical electrodes show more than a millivolt of CM. At low frequencies the limit seems to be set by harmonic distortion in the middle ear. In the middle and high frequencies, however, the maximum (or a plateau) is reached long before harmonic distortion appears. At maximum the wave form is still very nearly sinusoidal. There is no ‘peak clipping’ such as we see when the limit is imposed by the middle ear. It is as though the efficiency of the intracochlear conducting mechanism were reduced, as in an automatic volume control circuit. A possible mechanism for this limitation on maximum output is suggested in a later section. Injury occurs at very high intensities, and the temporary depression or ‘hysteresis effect’ (224) that follows supramaximal stimulation suggests that some sort of stretch beyond elastic limits has occurred. Another unexplained effect, closely related to the nonlinear response at high intensities, is the ‘interference’ of two tones, first described by Wever, Bray and Lawrence (243,245). When one tone is above the limit of linear response, the response to a second simultaneous tone is also reduced. The effect is more marked the closer the two tones are in frequency. (Apart from such interference there is no masking of one CM by another. Unlike action potentials, which are followed by refractory pe riods, they simply superimpose.) Lerche (156) stresses (and in the opinion of the present reviewer overstresses) the similarity of interference to masking, and he adds some new experimental details. Interference is indeed one form, but only one form, of masking; and Lerche’s ‘explanations’ are chiefly a set of imperfect analogies to electron tube circuits. Wever and Lawrence (245) explicitly ascribe interference to some process in the hair cells. An alternative is suggested below in a later section which will relate the interference of one tone with another to the same process that determines the maximum voltage of the cochlear microphonic. This process is ascribed to mechanical events in the cochlear partition prior to the bending of the hairs. ACTION POTENTIALS OF THE AUDITORY NERVE The action potential of the auditory nerve as a whole can be recorded, reasonably free from contamination by CM and SP, by means of conventional electrodes. Recently the single fibers of the first order neurons have been studied by means of hyperfine microelectrodes (227, 105, 226). Two previous studies (91, g4), believed at the time to represent such first order activity, have now been shown to deal with second order neurons in the cochlear nucleus (95, 228). They therefore lie outside the scope of this review. Single Fiber Activity. The following description of single fiber activity is condensed from Tasaki’s extensive study (227). The single fiber responses are, of course, all-or-none. Their time course is similar to that of spikes recorded in other nerves. The rising phase is very sharp and the falling phase is about half a millisecond. Many of the eighth nerve fibers show spontaneous discharges, more or less regular in rhythm, from IO to IOO per second. Many of the most active fibers probably are afferents from the vestibule; others might be the efferent bundle of Rasmussen, and sometimes discharges are induced by injury, Jamary rg57 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 29 but most of the afferent fibers that show normal auditory responsesto sound also show occasional and sometimesmany spontaneous impulses. Tasaki further states explicitly that he never observed a casein which the spontaneousdischargesof impulseswere inhibited by a sound stimulus. A singlefiber usually respondsto a single soo-cycle ‘tone pip’ (63) with a seriesof one to three nerve impulses.The impulsesare synchronized with the individual sound waves. The responseof the whole nerve to such a signal is extremely stable but in the individual fibers the latency of the spikes varies greatly and so does the threshold. The fiber also respondsto an 8ooo-cpstone pip with double or triple spikes.They are not synchronized with the sound waves but are separated by intervals of about I millisecond, corresponding to the refractory period. This multiple responseseemsto be the chief basis for the second peak in the whole nerve responseknown as ‘Ni, although it is possiblethat Ns also receives somecontributions from the secondary neurons in the cochlear nucleus (137). With strong &oo-cps tone pips the latency of the earliest single-fiber action potentials varied from 1.1 to 1.3 millisecond, under conditions for which the latency for the whole-nerve responsewas very stable, close to 1.0 millisecond. The position of the microelectrode was halfway between the spiral ganglion and the internal auditory meatus. Evidently the action potential of the whole nerve, as recorded with wire electrodes in the basal turn, is generated by the nerve impulsestraveling in the modiolus. In responseto pure tones the nerve impulsestend to appear at approximately the samepoint in the cycle of the stimulating sound even though the intervals between spikes are much longer than one cycle and additional spontaneous impulses may appear in other phase relationships. Study of the phase relation in which the spikes appear confirms the conclusion derived from studies of the whole nerve that excitation occurs during the phase of the microphonic in which Scalamedia is becoming more negative relative to Scalatympani. With strong stimuli the excitation seemsto occur just after the positive peak of CM. Near threshold the latency is longer and excitation occursjust before the end of the falling phase.Occasionally, if the frequency of the soundis 500cycles per secondor less,the nerve impulsesin a given fiber showa different phase relation to the microphonic. These occasional fibers are presumably fibers from the more apical portion of the cochlea where the traveling waves arrive only after a time delay. The majority of fibers encountered by Tasaki could give impulsesin responseto tones or pips up to at least 7000 or 8ooo cycles per second, and were presumably fibers from the basal turn. Only 2 out of 30 fibers had limiting frequencies between IOO and IOOO cycles per second.The limiting frequency of a nerve fiber never changed during the courseof an experiment and dependedonly slightly on the intensity of the tone. In general the nerve fibers arising in the basal turn of the cochlea respond to tones of any audible frequency but the fibers arising in the upper part of the cochlea respond only to low frequency tones. As it was difficult to keep a microelectrode in one nerve fiber for a period longer than about IO seconds,the relation of threshold to frequency was studied by stimulating with successivetone bursts of different frequency that followed one another at intervals of about 50 milliseconds.The intensity of the tone bursts was also varied systematically and thus the ‘responsearea’ was plotted. The boundary of this area is always extremely steep on the high frequency side at a ‘limiting frequency’. This steep side is comparable to the corresponding part of the Galambos and Davis (94) curves for the secondary neuron. The rise in threshold on the low frequency side of the area is extremely gradual, however. The difference between such a gradual slope 30 HALLOWELL DAVIS Volzdme 37 and the slopes found in the cochlear nucleus by Galambos and Davis is attributed by Tasaki to some sort of inhibitory action (91) taking place in the cochlear nucleus. A given fiber seemed to be quite stable with respect to the number of impulses, whether one or several, that it would yield in response to a given stimulus and there was some tendency of fibers to group according to their thresholds, some high, others low. With stimulation by steady tones the frequency of discharge in individual fibers gradually diminishes. Following a relatively strong tone pip impulses may continue in an after-discharge for as long as 30 milliseconds after the end of the pip. Action Potentials of Whole Auditory Nerve. Studies of the action potential response to clicks by Rosenblith and Rosenzwieg (191, 193, 194, 198, rgg, 192) have led to conclusions in close conformity with those derived by Tasaki from the single nerve fiber. In particular the initiation of nerve impulses early in the phase of positive-to-negative in Scala vestibuli was established rather clearly. Fluctuations of threshold were also found in this type of experiment. The amplitude of N1, studied as a function of intensity, was found to be nonlinear. It rises slowly at first but then, some 40 decibels above threshold, the increase suddenly becomes much more rapid and Nz appears clearly. The discontinuity indicates that there are two populations of excitable elements with different distributions of excitability among them. The recovery curve for N1 is a monotonic function. The amount of depression of the second response depends on the intensity of the first click as well as the interval between clicks and it considerably outlasts (about IO msec.) the recovery period of nerve fibers. The latency of the action potential relative to CM in response to high-frequency tone bursts is a function of intensity, of the rate of rise of the intensity of the tone burst, and perhaps also of the frequency (I 70). The minimum latency is 1.05 milliseconds (or perhaps 0.55 ; ref. 59) ; the maximum about 2.3 milliseconds. The minimum figure is considerably less than that given by Tasaki (227), but the conditions of Tasaki’s experiment did not allow the use of very intense stimuli (cf. also 39). Masking. Masking of action potentials is very prominent. Another noise delivered simultaneously, particularly if it overlaps the frequency spectrum of the stimulus for the AP and is of comparable intensity, greatly reduces the amplitude of N1 and N2 or even completely abolishes them. The relationships are not quite the same, however, as for ‘interference’ of CM noted above. Not much has been added in recent years to the physiological observations on masking. It is still interpreted as being due to a pre-empting of the activity of the nerve fibers by asynchronous impulses generated by the masking sound. Upper Frequency Limit of Response. In numerous studies the upper frequency limit for a visible cochlear microphonic response has been determined for various species (cf. 256), and sooner or later these data are quoted elsewhere as a measure of “the upper limit of ‘hearing’ as determined by electrical methods.” This, of course, means something very different from a limit determined by behavioral methods; and if any peripheral electrical method is to be used it logically should be the threshold of AP, not CM. But the presence of CM does give a presumption of possible auditory function, for it implies that hair cells can vibrate in the inner ear at that frequency. It is reassuring now to learn (170) that for guinea pigs the upper frequency limit for both AP and CM (and SP also) is the same, at about 50,ooo cycles per second, within the limits of the method of observation. Fatigue and Accommodation. The cochlear microphonic shows no measurable Janlcary 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 31 fatigue effects short of injuriously high intensities of sound. On the other hand adaptation, both fast and slow, in the overall AP discharge has long been familiar (69). Tasaki (227) reports a slowing of the discharge of action potentials in single units during constant stimulation, and following masking the depression of AP long outlasts the recovery period (I rg). Interest in the question of fatigue and adaptation of AP and also their possible relation to the early stages of acoustic trauma and to M&&e’s disease (i.e. sense-organ hearing loss) has been aroused by studies on human listeners of the shifts in threshold and also shifts in apparent loudness of sounds, both during and shortly after the end of stimulation (186, 43, 115-117, 127, 128). Very marked changes in human sensitivity have been demonstrated in this way and the recovery curve of threshold after moderate stimulation is not monotonic. At least two simultaneous and partly opposing processes must be assumed in order to account for the complex multiphasic recovery curves. In spite of these suggestions from psychoacoustics, however, only very little in the way of possible physiological correlates has yet been demonstrated by electrical methods in the ears of animals (193, 199). An Efferent Inhibitory Action. Stimulation, in the medulla, of the tract of Rasmussen produces an inhibitory effect on the AP response to clicks (93). N1 is clearly reduced, but CM is not affected. The effect is very specific with respect to the location of the stimulating electrodes, and the middle ear with its intra-aural muscles is definitely not involved. The reduction appears some 20-30 milliseconds after stimulation has begun and increases up to about 250 milliseconds. Rather rapid (3o-4o/sec.) stimulation is required. The optimal frequency is IOO per second. The long latency and the repetitive stimulation show quite clearly that this efferent inhibitory action is not related to temporal priority of nearly simultaneous bilateral signals but is apparently an expression of a rather general principle, namely, central regulation of the sensitivity of sense organs. The functional relationships of this reduction in sensitivity are completely unknown. Vulnerability of AP. The efferent inhibitory action, which leaves CM unaffected, is only one example of the greater vulnerability of AP to almost all adverse circumstances. AP is more sensitive to anoxia and recovers less readily. Quinine may abolish AP quite selectively (60). The latency of AP but not of CM is prolonged by cold (42). These relations are in accord with the theory that AP is the last link and CM an intermediate link in the causal chain of auditory events. Special interest therefore attaches to the relative resistance reported for AP to dihydrostreptomytin (II& 120). Here CM, notably the maximum voltage of CM, is reduced far more than AP. Relation to Frequency of Pure Tone. Pure tones up to 2000 cycles per second yield clear synchronous volleys of action potentials. Wever’s (242) volley theory is fully substantiated. Actually the synchronization is closer and the overall peak sharper in response to a IOOO- or soo-cps tone than for the usual unfiltered click or tone pip (rgg). But in addition to the sharp volley, which represents the nearly simultaneous discharge of the entire basal turn, there is a ‘tail’ of impulses coming from the more apical region of maximal amplitude where the phase differences and therefore the time differences become considerable. Therefore synchronization is less precise near threshold where the synchronous basal discharge is small. Furthermore, it is increasingly difficult to determine the real threshold for neural activity at low frequencies. This may be one reason for the very strong inverse relation of threshold of AP to frequency reported for the guinea pig (60). At frequencies above 3000 cycles per second the neural discharge becomes increasingly randomized and a steady tone of 4000 cycles per second or more is effec- 32 HALLOWELL DAVIS Vohme 37 tively a steady stimulus. However, there is a well-synchronized volley of action potentials a) at the onset of a steady tone, b) in responseto any rapid increase in intensity, or c) in responseto any other acoustic transient. The neural dischargeseems to be determined by the envelope of the acoustic stimulus when it can no longer follow the individual sound waves. General conclusions from the observations on synchronization and the variability of latency in individual fibers are: I> Time information, which is known from psychophysics to be both preciseand important, is conveyed statistically by groups of fibers, not by each individual fiber. 2) The basal turn, where vibration is in phaseat medium and low frequencies,is responsiblefor the volley principle and for any contribution of this principle to the discrimination of frequency (pitch). 3) The basal turn, where synchronized impulsesare evoked by changesin the intensity of high frequency sounds, is in general the time-sensitive portion of the cochlea. Effects of Physical Variables. Electrical polarization of the cochkar partition. Both the pseudo-threshold of the cochlear microphonic and its maximal amplitude can be altered by passinga direct current acrossthe cochlear partition (231). When the current flows from Scala vestibuli toward Scala tympani, which would tend to increase the positive polarization of the reticular lamina, CM may be increased in voltage by as much as IOO per cent. The action potentials are also increasedin magnitude for a given strength of stimulus. The opposite direction of polarization reduces or even suppressesboth CM and AP, but no reversal of phase of CM was seenand currents greater than 200 microamperesinjured the cochlear partition. Temperature changes.Lowering the body temperature of a hamster, down to 18”c, reducesthe amplitude of both CM and AP. AP declinesmore rapidly than CM and disappearsfirst. Its latency is greatly increased. The latency of CM, however, remains constant over the entire range (135). A curious effect following local cooling of the cochlea of the guinea pig (in the region of the secondto fourth turn) is first the complete but reversible disappearance of CM, including the anaerobic CM,. Then, while the cochlea is warming up again, CM showsstrong but varying amplitude modulation, unsymmetrical nonlinear distortion, and large phase shifts (42). It seemsprobable, but is not proved, that these effects are due to a temperature-dependent change in the mechanical properties of someof the intra-cochlear structures. Recovery from asphyxia is slower when the cochlea has been cooled (42). IncreasedendoZpz@tic pressure.In the courseof injections into the Scalamedia (58), the endolymphatic pressure has frequently been increased considerably. The pressureswere not measured but some of the injections were sufficient to rupture Reissner’smembrane. When the membrane did not burst EP was increased by as much as 10-q millivolts, but CM, AP and SP were all temporarily reduced. The reductions in the electrical responsesmay have been due in part to a stiffening of the cochlear partition and reduced acoustic transmission. Physiological and Pharmacological Effects. oxygen supply, carbon dioxide and blood sugar Zevel.The dependenceof a major part, i.e. CMl, of the cochlear microphonic on an adequate oxygen supply has been reaffirmed many times (252, 35-38, 17, 259, IOO, 85). The recovery of CM1 is reversible unlessthe anoxia has been pro- January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 33 longed and severe. Very severe or repeated bouts of anoxia cause degeneration of the hair cells (in cats), and even with less severe or prolonged anoxia changes in cells of Hensen and of Claudius and in the stria vascularis can also occur ( ISI). The action potential response is even more closely dependent on the oxygen supply than is CM1 (104) . Its failure is less readily and completely reversible (85, 33) The very prompt failure of AP and of CM argues for a high metabolic rate of the nerves and sensory cells (85) and the absence of any significant storage mechanism for oxygen (38). The endolymphatic potential is as sensitive as CM1 to oxygen - - lack (17) and recovers as promptly when oxygen is readmitted ($3). Very high concentrations (25 95) of carbon dioxide reduce CM1 (259). Extreme hypoglycemia produced by insulin injections can depress CM1 and AP (258). AP is more sensitive than CMI, just as for oxygen lack, Wing gives 40 milligrams per cent as a critical level for failure, but Fern&de2 (85) states-that both AP and CM1 can tolerate levels as low as 20 milligrams per cent. In fact, the ability to resist the expected effects of lowering the blood sugar seems to be the outstanding feature of these studies. In&a-cochkar injections. The histological effects on the cochlea of oral or parenteral administration of drugs such as ascaridole, streptomycin, quinine, etc., have been reviewed in a previous section and parallel changes in CM have been demonstrated (133, 134, II@. The intracochlear injection of drugs into either Scala tympani or Scala media has been announced as an experimental method but few details are yet published (65, $3). A definite and striking effect, however, is the prompt abolition of cochlear microphonics, endolymphatic potential and action potentials by small doses of sodium cyanide in any one of the three scalae. Apparently, the cyanide penetrates readily through structures such as Reissner’s membrane which are impermeable to potassium and sodium ions. This effect of cyanide, like the sensitivity to anoxia or asphyxia, argues clearly for an oxidative metabolism as an essential and immediate feature of the response mechanism of the cochlea. The perilymph may be completely replaced by Ringer’s solution without significant depression of the electrical responses of the cochlea (231). If Ringer’s solution is injected into Scala media, however, there is a rather rapid failure of all electrical responses, although not so rapid as following the injection of cyanide. The fluid that can be injected into Scala media without injury (65) is one which follows the usual Tyrode formula except that the potassium concentration is high and the sodium low according to the ratio determined (2 18, 219) by direct analysis of endolymph. This result is an indirect confirmation of the correctness of the chemical analysis. On the other hand, if this ‘endo tyrode’, or a solution made of one part of isotonic potassium chloride and three parts of Ringer’s solution (231)~ is injected into Scala tympani the action potentials and the cochlear microphonics fail rather rapidly and quite completely b.These solutions, high in potassium, when injected only into Scala vestibuli, however, have no such depressing effects. The conclusions (231, 229) are, nrst, that the basilar membrane is easily permeable to ions such as potassium and sodium while Reissner’s membrane is not; and second, that the fluid in the canals within the organ of Corti, such as the space of Nuel, is perilymph and not endolymph (see fig. 4). It would not be possible, incidentally, for the bare nerve fibers in the organ of Corti to conduct nerve impulses if they were immersed in a fluid of the composition of endolymph. Third, by implication, the nutriment of the hair cells comes not from the endolymph but from the 34 HALLOWELL DAVIS V olzcme 37 Scala tympani. Apparently we should think of the basilar membrane and its covering of mesothelial cells as a rather open although elastic net that is of considerable physical importance because of its stiffness but which does not serve as a barrier to diffusion. The reticular lamina and the cuticular surfaces of the hair cells both do serve as barriers to diffusion and also form a rather solid plate of mechanical importance. Reissner’s membrane and the cells of Claudius have little mechanical importance but do prevent diffusion. The problemof acetytcholine. A puzzling observation is an apparent prolongation of the latency of cochlear potentials following the parenteral injection of physostigmine or other substances that inhibit the action of cholinesterase. Gisselson (98) has described such an effect and has satisfactorily controlled the possibility that the observed changes might be due to tonic contractions of the intra-aural muscles (gg). The potential observed by Gisselson is apparently the cochlear microphonic. This is the only set of observations that suggests a latency between the mechanical movement of the cochlear partition and the appearance of the cochlear microphonic. Gisselson’s interpretation that acetycholine operates as a chemical mediator between the movement of the cochlear partition and the generation of the cochlear microphonic is completely at variance with all other relevant observations and interpretations, and remains unexplained. The action of a chemical mediator between hair cell and nerve fiber was suggested long ago (68) to explain the latency between CM and AP, but is no longer supported (64). Some otologists believe, however, that acetycholine is somehow involved in M&&e’s disease (5, 257). Acetylcholinesterase has been found in the intracochlear fluids (98) and more specifically in the organ of Corti, within or near nerve endings rather than in or on hair cells, and more plentifully near the internal rather than the external hair cells (44a). It has been suggested that it might be associated with the efferent fibers of the olivo-cochlear bundle (44a). Circ~Zution in ttte cochlea.Much attention has been given by otologists to the circulation in the cochlea (6). Capillaries in the region of the stria vascularis seem to behave much like capillaries elsewhere in the body in response to strong stimuli such as anaphylactic shock. There is some question as to the possibility and significance of blocking of capillary flow by ‘blood sludge,’ The possibility of sympathetic control of the capillaries of the cochlea has been both affirmed (211, IO) and denied (141, 145, 169, 177) with the weight of evidence against a significant amount of such control. In cats a large fall in systemic blood pressure (35, IOO) or a large increase in intracranial pressure (60 mm Hg) reduced the cochlear potentials on an average of 17 decibels (I&. These effects are probably due to interruption of adequate oxygen supply to the inner ear. Mt%zi&e’s disease. A major reason for clinical interest in the circulation in the inner ear is the possibility that disorders of this circulation, such as might be ascribed to local vascular spasm or to blockage by sludged blood, might cause the symptom complex known as Men&e’s disease. The presenting symptoms are periodic attacks of vertigo, a variable elevation of the auditory threshold, tinnitus and loss of auditory discrimination. Excellent reviews are available (113, 5, 257). The small amount of autopsy material that is available shows that Reissner’s membrane bulges far into the Scala vestibuli (71, I 14, s>. It is inferred that there has been a temporary elevation of the hydrostatic pressure in Scala media. The symptoms are usually attributed to this increase in pressure, and the increase in pressure is usually assumed to be secondary to local anoxia, based in turn on circulatory changes. It is not clear how or January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 35 why the circulatory changes could produce the increased pressure but it is perfectly plausible, according to the theory suggested in the next section, that the increased pressure might elevate the auditory threshold and also cause tinnitus. It has also been suggested, however, that changes in the chemical composition of the endolymph may be the cause of the symptoms (51,155,214). In animal experiments very minor surgical damage to Scala media, as by the insertion two or three times of a pipette of 15~ outside diameter, is often sufficient to cause a slow fall in EP, SP+, CM1 and AP. We believe that this depression is due to the leakage of small amounts of tissue fluid into Scala media. The suggestion that major changes in ionic composition of the endolymph are responsible for M&&e’s syndrome must remain speculative, however, until endolymph is collected and analyzed from a patient who is actually suffering from the symptoms at the time of operation. In addition to tinnitus and an elevated threshold, patients with M&i&e’s disease Suffer a loss of auditory discrimination and show ‘loudness recruitment’. Recruitment means that, in spite of the elevation of the auditory threshold, sounds at levels well above threshold sound as loud or nearly as loud in the affected ear as in the normal ear. This sign is now generally accepted as indicating a ‘sense-organ hearing loss’ as opposed to a conductive hearing loss on the one hand and to a nerve hearing loss, such as may be caused by pressure of a tumor on the fibers of the eighth nerve, on the other (72, 73). The mechanism of recruitment is not known, although some interesting speculations have been offered (204). In loudness balance tests the recruiting ear seems to ‘adapt’ very rapidly and profoundly with continued stimulation although it gives a fairly normal ‘on-effect’ (128, 129). This suggests that the sense organ is unable to sustain some necessary metabolic activity at its normal pace although it may accumulate some small reserve. In this connection it is sign&ant that pathological changes in the organ of Corti and also in the stria vascularis have been described (71, 113). Careful analysis of the signs and symptoms in Men&e’s disease and other abnormalities of the cochlea should provide useful clues for a better understanding of normal function. Formation and Chemistry of Labyrinthine Fluids. The subjects of the composition, formation, circulation and reabsorption of perilymph and endolymph have been adequately reviewed in the earlier section on anatomy. HYPOTHESES AND MODELS In this final section we will attempt to unify in a set of theories or models, some of the facts that have been summarized above. Some of the theories are old and some new. The models are tentative, but they should suggest further experiments and they emphasize certain gaps in our knowledge. Theories of Mechanical Action. The middle ear has not been included in this review, but Wever and Lawrence have given an excellent account of its action and have reviewed previous theoretical interpretations (249). Two leading ideas are that the drum and ossicles match the impedance of the air to that of the fluid of the inner ear and that the intra-aural reflex serves primarily a protective function, We have reviewed the dynamics of the cochlea above and have nothing to add to the account and interpretation given by Bekt%y of the traveling wave pattern of vibration of the cochlear partition and the relation of traveling waves to the principles of resonance, coupling and damping, or to Zwislocki’s discussion of the mathematical theories of cochlear action. 36 HALLOWELL DAVIS Vdzmze 37 Electrical Amplification in the Cochlea. The electrical phenomenaof the cochlea have been describedand the anatomical locus of the origin of each has been identified as closely as present evidence permits. We shall now assemblethese fat ,ts in a single model with the help of a few additional assumptions. Possibly some of the same relationships found in the cochlea may hold also for other senseorgans such as those of the labyrinth, the lateral line organ (in fish), and perhaps even for mechano-receptors in general. A special point of interest in the cochlear microphonics and the summating potentials is that they may represent the action of a biological amplifier ahead of the excitation of nerve impulses(so, 55, 17, 19). The very small amount of acoustic energy that is neededto reach the threshold of human (or animal) hearing under good conditions raisesgreat theoretical dficulties (238, 240) in respect to possiblemasking of the signal by thermal energy (Brownian movement) unlesssomesuch amplification takes place at the end of the chain of mechanical events described in the previous chapter. The average amplitude of movement of the basilar membrane at threshold is of subatomic magnitude (27). In other words, we can expect with someconfidence that the bending of the hairs of the hair cells should releaseor modulate the flow of energy from somebiological source. Simple transducing of the mechanical into electrical energy followed by electrical stimulation of nerve endingswould be too wasteful (25). This is one reason,although not the only one, why the present reviewer has now explicitly given up the simpler piezo-electric (t ransducer) theory of the genesisof cochlear microphonics (cf. 222, 224) in favor of some form of amplifier or trigger action (so, 52, 55). A Theory of Electra-Mechanical Relationships. Relations of CM and SP to EP. The endolymphatic potential (EP) is sensitive to mechanical displacement of the basilar membrane. Displacement toward Scala tympani increasesEP, toward Scala vestibuli diminishes it. The change in polarization is maintained if the displacement is maintained. The displacement may be caused by direct manipulation (17), by injection of fluid, or by spontaneous contractions of the intra-aural muscles (in guinea pig, ref. 58). Injections of fluid into Scalamedia or into Scalavestibuli both causean increasein polarization. Reissner’smembrane moves in opposite directions in the two situations and therefore it cannot be responsiblefor the changesin EP. If the movement of the partition is slow and periodic it is impossible to distinguish the corresponding changein EP from CM. B6kesy (17) implicitly assumedthat they are the same thing. He showed that the change in EP continued as long as displacementwas maintained and was related to displacement and not to acceleration and he then argued from these observations that the energy for CM was derived from a biological sourceand that CM was proportional to amplitude of displacement. This identification of CM1 with modulation of EP is compatible with our conclusionsas to locus of origin. Also both CM1 and EP are similarly sensitive to oxygen lack, and minor differences in the time course of their failure under oxygen lack are probably due to changes in the electrical resistance of Reissner’s membrane, etc. (x7, 19, 4* The summating potentials also seemto be closely linked to both EP and CM1. The chief divergence seemsto be the persistenceof negative SP for sometime after EP and CM1 have disappearedfrom lack of oxygen. Relation of CM and SP to mechanicalmovement.We will assumethat SP, like CMI, is produced by the bending of hairs in the proper direction. The hairs are bent by the shearingaction between tectorial membraneand reticular lamina. Symmetrical vibration produces CM : unsymmetrical displacement produces SP. January 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER FAR 37 We suggestthat the unsymmetrical bending that is responsiblefor SP is a consequenceof the one-way movement of the traveling waves on the basilar membrane. Perhaps the eddies described by Bekesy produce a longitudinal one-way movement of the tectorial membrane. In any case, traveling waves that can produce eddies should tend to move the tectorial membrane apically relative to the basilar membrane just like the fluid. We assumethat this longitudinal movement does not become significant until moderately strong stimuli are used. When the longitudinal movement does become significant, however, the tectorial membrane is subjected to an additional stressand we assumethat becauseof this stress it no longer moves transversely as readily as before. This restraint is, we suggest,the causeof the nonlinearity of the CM response at high levels, and may even account for the CM going through its maximum and becoming smaller at very high intensities. The negative summating potential becomesvisible a little below the intensity of stimulation where the CM responsefirst becomesnonlinear. This is a reasonable relation if an apical displacementof tectorial membranerelative to the organ of Corti is assumedto be the causeof the nonlinearity. The continuing longitudinal stress,causing a net one-way bending of the hairs, should cause a corresponding one-way potential, namely the negative summating potential. Bek6sy observed just such a negative change in the potential of the perilymph in Scala vestibuli when, with intact Reissner’smembrane, he displaced the tectorial membranelongitudinally toward the apex. In fact, when he applied a needle that vibrated from side to side to the middle of the tectorial membrane the strongest microphonics appeared when the direction of the vibration was longitudinal (22). Other Implications of the Theory. The phenomenon of interference becomes noticeable when nonlinearity appears (156). The interference we ascribe to an extension of the apical displacementof the tectorial membranefrom the area of the interfering tone to the area of the tone whose CM is depressed.The greater interference of tones closetogether in frequency is a natural consequenceof this mechanism.The nonlinearity of CM output at high intensities and the consequent maximum of CM voltage is the limiting caseof interference, in which the interfering tone and the target tone are one and the same. Indirect evidence that SP is due to an unsymmetrical mechanical action of some sort is found in the phenomenon of ‘remote masking.’ If a fairly strong amplitudemodulated high-frequency tone or a high-frequency band of noiseis delivered to the cochlea,electrodesin one of the upper turns of the cochleawhere the carrier frequency doesnot appear will show a CM corresponding to the modulation or envelope of the cafrier (66). The slow CM arrives at the apical turn with a delay corresponding to a traveling wave. The delay proves that the transmissiontoward the apex is mechanical, not electrical. Any AP excited in the apical region by a low-frequency tone may be maskedby the modulating signal. Of course in the basal turn, SP, varying with the root-mean-squareamplitude of the carrier, appearsin addition to the high-frequency CM. The CM pattern showsthat the demodulation doesnot occur in the middle ear. We infer that demodulation does occur mechanically (not electrically) in the basal turn and generates there the low-frequency mechanical waves that travel as usual toward the apex. It is not clear to what extent such demodulation of a high-frequency noise or carrier wave may contribute to normal auditory function. Demodulation, revealed by SP, occurs at fairly high sound levels, but within the physiological range. It fulfills the postulate (sg) that there must be somesort of ‘summation’ in auditory stimu- 38 HALLOWELL DAVIS Volume 37 lation at high frequencies. Whatever the mechanism, SP is the electrical sign of such a summation. The present interpretation locates the summation in an unsymmetrical mechanical process, not, as we formerly believed, in temporal or spatial summation of neural excitatory effects. Positive and negative summating potedials. The positive summating potential is produced, we suggest, by a one-way displacement of the tectorial membrane radially or perpendicularly, just as the negative summating potential is produced by a longitudinal displacement. The viscous tectorial membrane may not follow the movements of the basilar membrane in one direction as readily as in the other, and the resulting net displacement could produce the positive summating potential. The net displacement must be ‘downward’ toward the Scala tympani because an increase in static positive pressure in Scala media (without traveling waves) causes an increase in the positive potential of Scala media ($3). If there is, as B6k6sy’s observations suggest, a preferential activation of external hair cells by longitudinal vibration and of internal hair cells by perpendicular vibration, we should ascribe the negative SP to the external hairs and the positive SP to the internal. There are more external than internal hair cells. Therefore, with strong stimuli the negative summating potential should be larger than the positive, which is actually the case. Our best explanation of the apparent increase in negative summating potential under anoxia is that the anoxia reduces the opposing positive summating potential. This is reasonable, because BCk&y (22) tells us that the internal hair cells are more sensitive to anoxia than the external hair cells. Most theories of cochlear action ascribe a lower threshold to the external than to the internal hair cells, largely because the external hair cells are more vulnerable to injury. It now seems, however, ($3) that the threshold for positive SP, apparently associated with the internal hair cells, is lower than for negative SP. Perhaps the same relation holds for CM also (cf. 139). In order to explain the curious sensitivity of the summating potentials to changes in the chemical composition of the endolymph we assume that the physical state of the tectorial membrane is sensitive to such changes. It is a colloidal gel, a mucopolysaccharide or glycoprotein, and it may change its state of hydration and its viscosity and thus ‘creep’ more or less readily. Its marked shrinkage under the influence of fixing agents is well known, and it is one of the few structures that is fully exposed to the endolymph. The unique high potassium concentration here may be essential to preserving the normal colloidal state of the tectorial membrane and may not be related to the endolymphatic potential. According to our hypothesis the positive SP should have its maximum at the point of maximum amplitude of displacement of the basilar membrane and should be relatively strong on the basal side of the maximum. The maximum of the negative SP, on the other hand, should be farther toward the apex, for this is where the longitudinal eddies are observed. The negative SP should therefore be strongest on the apical side of the maximum. This prediction seems to be fulfilled. Goldstein (104) found, with dBerentia1 electrodes in the basal turn, a strong negative SP for 8ooo-cps pips, no SP with a 2000-cps pip, and a weak positive SP with soo-cps pips. Continuing our speculation, the negative summating potential should be an adequate stimulus for the auditory nerve fibers, or at least lower their threshold. It might cause a repetitive discharge of impulses just as a continuing depolarization of a dendrite causes repetitive firing in the axon. The proposition that the negative summating potential should be excitatory is in agreement with the observation that it is the phase of CM in which Scala media becomes negative that initiates the nerve JanmY 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER FAR 39 impulse. On the other hand, the positive summating potential should tend to inhibit the discharge of nerve impulses. Note, however, that the summating potentials appear only at moderate and high intensities. An inhibitory action in the auditory nerve centers that increases the discriminating power of the system has been suggested repeatedly. The summating potentials, however, act peripherally. Our present theory and that of Huggins (131) are independent and are not antagonistic. Furttter impZic&ons of the &my. Some salient features and rather obvious implications of the foregoing theory are: I) Cochlear microphonic and summating potential both represent a valve action. Biological energy, probably derived from the endolymphatic potential, is released by the bending of hairs of the hair cells. Cochlear microphonic and negative summating potential both stimulate the nerve fibers. 2) The summating potential is part of a mechanism of response that provides for gradation of response at high intensities. 3) The cochlear microphonic and the summating potential supplement one another in a way that greatly extends the dynamic range of the ear. 4) Different directions of mechanical movement associated with the traveling wave pattern affect different hair cells and may produce different electrical and neural effects. Some effects, both direct and indirect, are presumably inhibitory and may serve to improve auditory discrimination. 5) The mechanism of mechanical restraint in the inner ear, revealed by the nonlinear and limited response of the cochlear microphonic, should act as a protective mechanism for the very sensitive low-intensity mechanism. 6) Demodulation, revealed by SP, makes the ear directly sensitive to the time and to the frequency of changes in amplitude of high-frequency signals. Origin and Function of Cochlear Potentials. InhceWar polarization and AR The negative intracellular polarization and also the action potential response are familiar but are only partially understood. Basic concepts are, I) differences in ionic concentration, particularly of potassium; 2) selectively permeable membranes; 3) sudden and specific alterations in selective permeability; and, in the background, 4) a metabolically driven ‘sodium pump’ (213). Endolymphatic potential. It is tempting to try to relate the endolymphatic potential (+8o mv) to the very unusual, if not unique, high potassium concentration of the endolymph by analogy with the familiar intracellular polarization. Inside cells the potassium concentration is high and the sodium concentration is low, just as in endolymph. The absolute magnitudes of many intracellular potentials and of the endolymphatic potential are almost the same. The fatal objection to such an identtication of the two polarizations is that they are opposite in sign. The intracellular potential is negative, the endolymphatic potential is positive. If we were dealing with a potassium concentration potential, we should rind zero potential difference across the cuticular surface of the hair cell between the interior of this cell and the endolymph. Actually there is a potential difference of 160 millivolts. Evidently the source of the endolymphatic potential is not a sodium or potassium ion concentration difference. Even as the basis for the negative intracellular potential the concept of ionic concentration difference has recently been challenged most seriously (I IO, I Ioa). It is not the only possible model for bioelectric potentials (212). In short, EP may not be related in any way to the unusual chemical composition of the endolymph. EP, and perhaps the intracellular potential also, seems to depend directly on a metabolic 40 HALLOWELL DAVIS Volume 37 activity that requires a ready supply of oxygen and is rapidly poisoned by cyanide. The recovery of EP after a single spontaneousgasp in Cheyne-Stokes respiration requires only a few seconds. We cannot even be sure that EP is the ‘store of available biological energy’ for our ‘biological amplifier.’ The negative intracellular potential of the hair cells also fulfills the requirements. It may be significant that no positive endolymphatic potential hasbeen reported in the utricle, although undoubtedly more than one investigator has sought for it; yet it is for utricular fluid that we have the most reliable analyses for high potassium and low sodium (cf. footnote 2). As far as the reviewer is aware, the positive endolymphatic potential in the cochlea is unique. Analogies among other potentials seemremote or else the potential is of the opposite sign, as, for example, frog’s skin (236), human skin (g), synovial membrane (I&), gastric mucosa(187), and the musclecell of the uterus whosepotential has been reversed by oxytocin (262). The closestanalog may be the cameo-retinal potential (163, 167a). Among other mechano-receptor organs no corresponding positive potential has been demonstrated; not in the Pacinian corpuscle, the muscle spindle, the stretch-sensitive neuron in arthopods, the much more closely related lateral line organ of fish, or even in the utricle. A specific search of these and other senseorgans might be rewarding. Can it be that the endolymphatic potential is merely a nonessential specialization of the cochlea? It might serve, for example, merely to give a hyperpolarization of the cuticular (hair-bearing) surface of the hair cells and thus increasethe sensitivity of response. Variable resistancetheory of thecochlearmicrophonic. As possiblesourcesof energy for our ‘biological amplifier’ we have, then, either the negative intracellular potential or the positive endolymphatic potential or both. The next problem is how the potential difference of 160 millivolts acrossthe cuticular layer of the hair cells is modified by bending the hairs to produce CM and SP. One specific theory for the derivation of the cochlear microphonic from the endolymphatic potential is that the bending of the hairs causesa changeof ohmic resistance at the cuticular surface of the hair cell (so, 52, 55). This is the point of greatest potential drop between endolymph and interior of hair cells, and some current is continually leaking through this barrier. The change of resistance alters the amount of current flow and thus produces CM by altering the IR drop in this part of the circuit. Excitation is caused, according to this theory, by the outflow across the nerve membrane of current that has passedfrom endolymphatic space through the hair cell, acrossthe nerve ending and into the fiber. At the submicroscopiclevel a movement of ions through submicroscopicpores that have electrically charged walls has been suggested(52; cf. also 213), but so far the electron microscope has not revealed anything corresponding to such a model. (Engstrom, ref. 77, suggeststhat the lateral walls of the hair cells, not their cuticular surfaces, do look like a plausible source of CM.) It is dif&ult to distinguish experimentally between a change in internal resistances(seriesand/or shunt) and a change in the EMF of the source.We shall, therefore, speak simply of ‘changesin the polarization of the cuticular membrane’ as a consequenceof bending the hairs and regard the change-of-resistancehypothesis as simply one possiblemodel of such a process. Art maZog of the co&ear micro#holtic and summating potential. The nearest analog to the postulated modulation of EP by mechanical stretch to give CM is the changein intracellular potential of a receptor neuron in the crayfish (81,82,148). The dendrites of this neuron are attached to special cells in a muscle and are stretched or relaxed according to the contraction of the muscle. The intracellular potential re- Ja-afy 1957 BIOPHYSICS AND PHYSIOLOGY OF INNER FAR 41 corded by a microelectrode decreases and increases as the dendritic processes are stretched or relaxed. The change in potential is maintained as long as the deformation is maintained. The change is in a ‘generator’ located peripherally in the dendrites. The change is conducted electrotonically to the body of the neuron and determines the rate at which the cell fires impulses. Furthermore, the dendritic potential may be reduced by inhibitory impulses arriving over efferent nerve fibers that end on the dendrites. This receptor neuron exhibits a cellular potential that is modulated by physical stretch, but it is a little puzzling just how to apply the analogy to the structures of the cochlea. Apparently the hair cell must be considered as a neuron which is totally dendritic in character, in the sense that it merely produces potentials which are conducted elsewhere electrotonically but carries no all-or-none impulses itself. Excitation of Nerve Impulses. An electrica theory. The third element of our theory of cochlear action is simply an electrical theory of excitation of auditory nerve impulses. It states that increase in the current flow, associated with the cochlear microphonic and the negative summating potential, stimulates the nerve. The nerve ending itself is assumed to be inert and merely to provide electrical contact. The whole terminal non-medullated section of the neuron is presumably excitable, and excitation may occur at varying distances from the hair cell according to the intensity of the microphonic and summating potential. This would explain the variation of latency with intensity of acoustic stimulus. Incidentally, the whole non-medullated section is anatomically unique in being completely bare and not surrounded even by a neurilemma1 sheath (2 I 7). It all presumably acts as the first node of Ranvier in the conducting pathway. In favor of the electrical theory, the latency of the earliest neural activity in the organ of Corti is too short for chemical mediation (64). Slow conduction in the nonmedullated terminals can explain the latency of AP in the modiolus. The excitatory phase of CM, i.e., increasing negativity in scala media, is associated with outward current flow across the membrane of the nerve fibers, as it should be. This relation is not immediately obvious, becausethe phaseof the cochlear microphonic (and consequently of the summating potential also) is opposite on the opposite sidesof the reticular lamina (229). Therefore a negative CM or SP in Scala media implies greater positivity within the hair cell. This should causea stronger flow of current from hair cell into the nerve endingsin contact with it and in turn a greater flow of current outward acrossthe cell membraneof the nerve fibers. This direction of current flow should tend to depolarize the membraneand thus excite nerve impulses. The old objection that the cochlear microphonic reachesits maximum well within the physiological range of acoustical intensities while the senseof loudness(and apparently also the size of action potential volleys) continues to increaseis now met by assumingthat the stimulating function is taken over by SP at high intensities. A theoretical objection that AC stimulation of nerve at audio frequencies is too inefficient has been largely offset by the studies of Rosenblueth, Luco and Garcia Ramos (195-197). Apparently under many conditions anodal and cathodal effects are not antagonistic but may summate to produce excitation. Among the observations that raise some dficulty for the electrical theory are those in which AP has been demonstrated in the presenceof very small CM. Streptomycin poisoning is such a case, and the vicissitudes of acute experiments provide others (60). Even the normal CM in man and monkey is small (168, 153). In spite of such difficulties, however, the electrical theory seemsto have gained widespreadtacit acceptance. HALLOWELL 42 DAVIS V&me 37 The chemicalmediutor theory. In contrast to the electrical theory, the chemical mediator theory states that a specific substance, such as acetylcholine, is required for the stimulation. It stands or falls with the resolution of the broader question of the role of acetylcholine and related substances in synaptic action and elsewhere (74, 166). At present an acceptable view is that, while acetylcholine or acetylcholine-like substances may alter the excitability of nerve endings, it is not essential for the excitation (215) and the time required for liberation and diffusion of acetylcholine is not the basis of the latent period of AP. Acetylcholine, etc. apparently can sensitize muscle spindles (ION), baroreceptors of the carotid sinus (70) and other sensory nerves such as cold, pain and taste (149, 215). It would be convenient to assume for the excitatory mechanism, whether electrical or chemical, that I) it is relatively insensitive to low frequencies (60), and also that z) it responds with repetitive discharge to negative SP as a constant stimulus. Unfortunately, these two assumptions are at least partly opposed to each other. In summary, we believe we can now give a fairly satisfactory account, with explanatory physical or mathematical models, of the dynamics of the cochlear partition, the formation on it of traveling waves, the relation of the position of the traveling wave pattern along the cochlea to frequency, and the smaller-scale movements in the sense organ that lead to a final critical event, the bending of the hairs. We also can give a fairly complete account of electrical responses in the cochlea and can relate them reasonably to the mechanical events. The origin of the electrical potentials is very obscure. The electrical theory of excitation of nerve impulses seems to be the most plausible. The behavior of action potentials is consistent with that of nerve impulses from other sense organs; but both anatomy and the complex patterns of cochlear activity have made the study of single auditory fibers both difficult and limited. Theories of Frequency Discrimination. Concerning patterns of auditory nerve impulses and their relation to the pattern of incoming sound waves on the one hand and to activity in the central nervous system on the other, we can make a few positive statements and also point to several gaps in our knowledge. We do know that the yolley principle, as proposed by Wever (242), is substantially correct. By rotation of activity a group of fibers reproduces in its total output of nerve impulses the frequency of the stimulating sound waves. This effect is clear up to 2000 cycles per second and detectable to 4000 cycles per second. Even at 500 cycles per second, however, we now know that not all of the fibers fire synchronously, due to the time lag of the traveling waves. We also know, with equal certainty, that individual nerve fibers have definite but different upper frequency limits. It is almost certain that the limit is associated with the position along the cochlear partition at which the fiber originates and thus in turn with the position of the traveling wave pattern as determined by frequency. Thus the place principle for the ‘coding’ of frequency information is established as well as the volley (frequency) principle. In psychoacoustics the duplex theory of hearing, which combines the two principles, is steadily gaining favor (140, 157, 63, 132, 30, 248, 161). The perennial problem of frequency discrimination is not solved, in spite of the very steep upper limits of the response areas of Tasaki’s individual nerve fibers. It seems to be agreed that some further ‘sharpening’ must take place by a neural inter- h?M&W~ I957 BIOPHYSICS AND PHYSIOLOGY OF INNER EAR 43 action that involves inhibition in the cochlear nucleus. The possibility of peripheral neural interaction has not been investigated sticiently. The significance of the multiple innervation and of the wide distribution of endings of nerve fibers among the external hair cells is unknown. Inhibitory efferent impulses seem to be a reality, but apparently they affect sensitivity rather than discrimination. Other Unanswered Questions. More attention should be given to the temporal aspects of excitatory and neural action. In the classical preoccupation with steady tones and the problem of frequency discrimination, transients and temporal relationships, including binaural localization, were largely overlooked. Hearing has a time dimension as well as a frequency dimension (53, go). In terms of the sense organ we may ask, for example, how the degree of damping and the natural period of the ear modify the synchronization of the neural output in response to various types of transients or other signals whose intensity is varying with time. Also, to what extent do fatigue and adaptation occur in the sense organ and how much must be attributed to the central nervous system? The problem of excitation requires much further study. It will not be fully solved until we can describe the intracellular mechanism, at the level of the electron microscope or even at the molecular level, by which the biological ‘preamplifier’ can operate with an input, at threshold, of subatomic amplitude. Also we need to know how oxidation maintains a bioelectric potential. There are physicochemical gaps also. What is the composition of the tectorial membrane? What are its physical properties and how do they depend on the oxygen supply and on the chemistry of the endolymph? The theory of the summating potential suggested above leans heavily on assumptions in this area. And can we perhaps separate into different pathologic, etiologic and therapeutic groups the patients who suffer from Meniere’s disease and ‘sense-organ hearing loss’? The physiologist can derive many helpful ideas from the clinic and from psychoacoustics as well as from anatomy, biophysics and biochemistry. REFERENCES In order to conserve space the editorial office of Physiological Rev&m has condensed certain titles by the elimination of unnecessary words. I. ALEXANDER, I. E. AND F. J. GITHLER. logical examination of cochlear structure exposure to jet engine noise. J. Cap. & PS@ZOt. 44: 513-524, 1951. 2. ALEXANDER, I. E. AND F. J. GITHLER. effects of jet engine noise on structure and of the cochlear apparatus. J. Cump. & PsychoZ. 45: 38r-391, 1952. 3. ALEXANDER, I. E. AND F. J. GITHLER. intense injury pure tone stimuli when magnitude is controlled. J. Expsr. Psychot. Histofollowing 8. AUTRUM, Physiol. Chronic function Physiol. Effects of of initial 45: 49-56, Nerven- Sinnesphysiologie. rgs2. g. BARNES, T. C., J. KARASIC AND M. D. AMOROSO. Further studies of rate of healing of human shin measured by electrical wound potential of experimental abrasions. Am. J. Swg. 82: 72~26, xgsx. IO. BEICKERT, P. Die vegetative Beeinflussbarkeit des Innenohres. Arch. Ohron- Nascn- ii. KchJRopfh. 163: 260-1769 wss. II. VON BEE&Y, G. Variation of phase along the basilar membrane with sinusoidal vibrations. J. Acoust, Sot. Am. rg: 49-460, 1953. 4. ALTMANN, H. F. AND J. G. WALTNER. fluids (rabbit). Ans. Circulation O#oL Rhk. und Fortschr. dcr ZOOtOgi6. g: 537-604, of & labyrinthine Larpg. 56: m-08, 1947. 5. American Academy of Ophthalmology and Otolaryngology. Symposium on M&&e’s Disease. 2’~. Am. Acad. Ophth. 60: 167-198, 1956. 6. American Otological Society. Symposium on blood circulation (in the inner ear). Anti. Otol. Rhin. & Laryng. 64: 1x64-1212, Igss. 7. American Standards Association. Relations of hearing loss to noise exposure. New York: Am. Standards Assoc., rgs4. 1947. 12. VON BEKESY, G. Elasticity of the cochlear partition. J. Acoust. sot. Am. 20: 227--241,1g48. 13. VON BEI&SY, G. Vibration of the cochlear partition in anatomical preparations and in models of the inner ear. J. Acoust. Sac. Am. 21: 23302451949. 14. VON B&&Y, G. Resonance curve and decay period at various points on the cochlear partition. J. Acowt. Sot. Am. 21: 245-254, Ig4g. IS. VON BEK%SY, G. Coarse pattern of electrical resistance in the cochlea of the guinea pig. J. Acoust. Sot. Am. 23 : x8-28, rgsr . 16. VON BEKESY, G. Microphonics produced by touch- 44 17. 18. IQ. 20. 11. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. 37. HALLOWELL ing the cochlear partition with a vibrating electrode. 1. Atowt. Sot. Am. 23: 2~-3S, 1951. VON BEKESY, G. DC potentials and energy balance of the cochlear partition. J. Acowst. Sot. Am. 23 : 576582,IQSI. VON BEK$SY, G. Direct observationof the vibrations of the co&ear partition under a microscope. Ada oto-laryng. 42: fQp201, 1952. VON BEICESY, G. DC resting potentials inside the cochlear partition. 1. Acowt. Sot. Am. 24: 72-76,1~52. VON BEKESY, G. Gross localization of place of origin of the co&ear microphonics. 3. Acoust. Sot. Am. 24: 3QQ-4%1952. VON BEKESY, G. Description of some mechanical properties of the organ of Corti. J. Acowt. SOC. Am. 25: 770385, WS3. VON BEKESY, G. Shearing microphonics produced by vibrations near the inner and outer hair cells. J. Acorut. Sot. Am. 25: 786700, 1953. VON BEKESY, G. Some &&o-mechanical properties of the organ of Corti. Ann. Otol. Rhin. & Laryng. 63:4&468,x954. VON BEKESY, G. Paradoxical direction of wave travel along the cochlear partition. J. Acowt. Sot. Am. 27: 137-16, IQ$$ VON BI%K&Y, G. Current status of theories of hearing. Scicroc6 123 : 779383, 1956. VON BEKBSY, G. AND W. A. ROSENBLITH. The early history of hearing-observations and theories. J. Acowt. Sot. Am. 20: 7277&, 1~48. VON BEK&Y, G. AM) W, A. ROSENBLITH. Mechanical properties of the ear. In: Handbook of Ex@rimeda Psychology, edited by S. S. Stevens, New York: Wiley, rgsr. BRLANGER, L. F. Autoradiographic detection of S*a in membranes of the inner ear of the rat. Scimx 118: 520-52X, 1953. BERG, K. The toxic effect of streptomycin on the vestibular and co&ear apparatus. Acta otu-Zarye SUPPI. 97:f-77,195f. DE BOER, E. & ths “1R6sidU6” iu Hcarifig. S-Graver&age: Excelsior, 1954. BOGERT, B. P. Determination of the &e&s of dissipation in the cochlear partition by means of a network representing the basilar membrane. J. Acoust. Sot. Am. 23: 151-154, 1951. BORGHESAN, E. Tectorial membrane and organ of Corti considered as a unique anatomic and functional entity. Ada oto-Zaryng. 42: 473-486, 1952. BORNSCHEIN, H. AND B. GERNANDT. Selective removal of the nerve discharge component from the cochlear potential during anoxia. Acta fihysiol. scandinav. 21: 82-89, 1950. BORNSCHEIN, H. AND F. KREJCI. Das Verhalten der Cochlear-potentiale bei Sauerstoffmangel. Mscht. Ohrenheilh. 83: I~I~~,IQ~Q. BORNSCHEIN, H, AND F. KREJCI. Untersuchungen Uber die Kreislaufabtigigkeit der Cochlearpotentiale bei Anoxie. Mscht. Okrenheitk. 83: 386-392, 1949a BORNSCHEIN, H. AND F. KREJCI. Uber die Frequenzabhtingigkeit reversibler Finderungen der Cochlearpotentiale bei temporllrer Anoxie. Experie&a 5: 359-369, 1949. BORNSCHEIN, H. AND F. KREJCI. Der Einfluss kunstlicher DurchstrCSmung auf den postmortalen Schwund der Cochlearpotentiale. &p6rientia 6: 67-68, 1950. DAVIS vohne 37 38. BORNSCHEIN, H. AND F. KREJCI. Beitrag zur Analyse des postmortalen Verhaltens der Cochlearpotentiale. Expcri6ntia 6: 271-273, 1950. 39. BORNSCHEIN, H. AND F. KREJCI. Eine Interferenzmethode zur Bestimmung der Latenzzeit van Aktionspotentialen des Innenohrs. Experientia 6: 354-357, IQSO40. BORNSCHEIN, H. AND F. KRE JCI. Zur Frage einer selektiven Ultraschallwirkung auf die Cochlearund Aktionspotentiale des Innenohres. Exfierientia 7: IOQ-112,IQ51. 41. BORNSCHEIN, H. AND F. KREJCI. Die Iatensitllts funktion der Cochlearpotentiale nach intravitalem und postmortalem akustischem Trauma. Experie&ia 9:6Q72,~95342. BORNSCHEIN, H. AND F. KRE JCI. Elektrophysiolog&he Untersuchungen tiber Temperatureffekte in der Schneclce. Acta oto-Zarpg. 4~: 467-478, 1955. 43. CARTERE’M’E, E. C. Perstimulatory auditory fatigue for continuous and interrupted noise. J. Acowt. Sot. Am. 27: 103-m, 1~5s. 4. CAUSSR, R., N. P. BW-HOI, I. GONDET AND N. HOAN. Modification des potentiels cochlCaires du cobaye sous l’influence de l’acide ac&ylaminophenylarsinique (arsachtfne). Cornpi. ~69~i. Sot. biol. 143: 68-70, IQ4Q. w. CHURCHILL, J. A,, H. F. SCHUKNECHT AND R, DORAN. Acetylcholinesterase activity in the cochlea. Laryngoscope 66: I-IS, 1956. 45. CITRON, L., D. EXLEY AND C. S. HALLPIKE. Labyrinthine fluids. &it. M, J, 12 : IOI-204, 1956. 46. COVELL, W. P. A cytologic study of the effects of drugs on the cochlea. A.M.A. Arch. Otolaryng. 23: 633-64~936* 47. COVELL, W. P. Peripheral endings of the cochlear nerve. Ann. Otol. RhiB. & Laryng. 47 : 63=67,x938. 48. COVELL, W. P. Histologic changes in the organ of Corti with intense sound. J. Conop. N~YoZ. gg: 43.~9, 1953. 49. CREMER, L. Uber die ungelosten Probleme in der Theorie der Tonempfindungen. Acwtica 2 : 83-~6, 1951. so. DAVIS, H. Energy into nerve impulses: hearing. (The Advancement of Science, March, 1953). Reprinted in bf6a. Butt. St. Louis Univ. 5: 4.3-481953. 51. DAVIS, H. Tinnitus: physiologic aspects. Z’r. Am. Ad. Ophth. 58: 527-528, 19~4. 52. DAVIS, H. Mechanism of hearing. In: Nerve Im$ds6 edited by D. Nachmansohn. (Tr. of Fourth Conf., Josiah Macy, Jr. Foundation). Caldwell, N. J.: PrOgrtSSS &SOCS., 1953. 53. DAVIS, H. Space and time in the central nervous system. Etectrocnc6phatog. & Clin. Neuro#hysioi. 8: 185-191, 1956. 54. DAVIS, H. The four electric potentials of the cochlea. Am. J. Physiol. In press. 5s. DAVIS, H. Initiation of nerve impulses in the cochlea and other mechano-receptors. In: Physiological Triggers, edited by T. H. Bullock. (Symp. Sot. Gen. Physiol.) Washington, D. C.: Am. Physiol. SOC., 1956. 56. DAVIS, H. AND ASSOCIATES. Acoustic trauma in the guinea pig. 1. Acowt. Sot. Am. 25: II~Q-I&, 1953. 57. DAVIS, H., C. T. MORGAN, J. E. HAWKINS, JR., R. GALAMBOS AND F. W. SMITH. Temporary deafness following exposure to loud tones and noise. Ada oto-tafyng. suppl. 88: 1-57, 1950. 58. DAVIS, H., B. H. DEATHERAGE, D. H. 3amwy BIOPHYSICS 1957 AND PHYSIOLOGX ELDREDGE m C. A. SMITE Summating potentials of cochlea. In preparation. H., C. FERNWDEZ AND D. R. 59s DAVIS, McAULIFFE. Excitatory process in cochlea. Proc. Nat. Ad. SC. 36: 58o-587, 1950. 60. DAVIS, H., B. E. GERNANDT AND J. S. RIESCOMAcCLURE. Threshold of action potentials in ear of guinea pig. J. Newophysiot, 13 : 73-87, 1~50. 61. DAVIS, H., B. E. GERNANDT, J. S. RIESCOMAcCLURE AND W. P. COVELL. Aural microphonics in cochlea of guinea pig. J. Acoust. SOC. Am. 21: sO2-510, IQQ9. 62. DAVIS, H., H. 0. PARRACK AND D. H. ELDREDGE. Hazards of intense sound and ultrasound. An*. otol. Rhin. & Laryng. 58: 732-738,1949. H., S. R. SILVERMAN’ AND D. R. 63* DAVIS, McAULIFFE. Some observations on pitch and frequency. J. Acoust. Sot. Am. 23 : m@, 1951. DAVIS, H., I. TASAKI AND R. GOLDSTEIN. Peripheral origin of activity, with reference to the ear. Cold Spring Harbor Symposia Quad. Biot. 17: 14.30 154, 1952. DAVIS, H., I. TASAKI, C. A. SMITE AND B. H. DEATHERAGE. Cochlear potentials after intraco&ear injections and anoxia. F6daration PYOC. 14: EAR 45 organ lations of Corti; supporting structures and their reto sensory cells and nerve endings. Acta OtO-tafyng. 43 : 323-334, 1953. 80. ENGSTRGM, H. AIUD J. WERSALL. Is there a special nutritive system around the hair cells in the organ of Corti? Ann. Oiot. Rhin. & Laryng. 62: 5070~12, x953* 81. EYZAGUIRRE, C. AND S. W. KUFFLER. Process of excitation in dendrites and soma of single isolated sensory nerve cells of lobster and crayfish. J. Gcn. Physiot. 39: 87-119, 1955. EYZAGUIRRE, C., AND S. W. KUFFLER. Further study of soma, dendrite, and axon excitation in single neurons. J, Gen. Physiot. 39: 12~153,r~~~. FERNhDEZ, C. Innervation of cochlea (guinea pig). hry#gOSC6#6 61: 11~2-1172, 1~51. FERNtiDE& C. Dimensions of cochlea (guinea pig). J. Acowt. SOG. Am. 24: 5x9-523, 1952. FERNbE& C. Effect of oxygen lack on cochlear potentials. An% Scot. Rhin. & &wy#g.bq:r1~3-1203, 1955. FERNtiDE& C., H. LUDWIG AND D. McAULIFFE. Electrocoagulation of vestibule in &lea PigS. hI~gOSCdp6 62: IISQ-1174, 1952. FLETCHER, H. Dynamics of the cochlea. J. Acowst. Sac. Am. 23: 637-66, 1951. FLETCHER, H. Acoustics, Physics Today 4: 12-18, 1129 1955. 66. DEATHERAGE, B. H., H. DAVIS AND D. H. ELDREDGE. Physiological evidence for the masking of low frequencies by high. In preparation for J. 19% FLETCHER, H. s#66ch and acaring in CmmunicaNew York: Nostrand, 1953. GABOR, D. Acoustical quanta and the theory of hearing. Natur6 159: 59x-#& 1~47. GALAMBOS, R. Inhibition of activity in single auditory nerve fibers by acoustic stimulation. J. Newo#kysiot. 7 : 287-303, IQ#. GALAMBOS, R. Neural mechanisms of audition. Physiot. Rev. 34: 497-528, 1954. GALAMBOS, R. Suppression of auditory nerve activity by stimulation of efferent fibers to the cochlea. J. Nsrrrophysiot. IQ: 4240437~ 1956. GALAMBOS, R. AND H. DAVIS. Response of single auditory-nerve fibers to acoustic stimulation. J. Acowt. Sot. Am. tbs. D. Egbct of Q&&w on Guinea-Pig Copenhagen: Centraltrykkeriet, I#$ A. J. AND H. DAVIS. Probable mech68. DERBYSHIRE, anism for stimulation of the auditory nerve by the organ of Corti. Ah. J. Physiot. 113: 35,193~. DERBYSHIRE, A. J. AND H. DAVIS. Action potentials of the auditory nerve. Am. J. Physiot. 113: 67a DEDERDING, Labyrinth. 47~504, OF INNER 193s. J. Observations on the excitation by 709 DIAMOND, acetylcholine and by pressure of sensory receptors in the cat’s carotid sinus. J. Physiot. 130: ~13-s32,r~S~. M. R. AND C. S. HALLPIKE. Pathology, 71. DIX, symptomatology and diagnosis of certain common disorders of the vestibular system. Ann. Otot. Rhin. dk Laryng. 61: 987-r016,19~2. AND J. D. HOOD. 72. DIX, M. R., C. S. HALLPIKE Observations upon the loudness recruitment phenomenon. Proc. Roy. Sot. Med. 41: 516526, rd. AND J. D. HOOD. 739 DIK, M. R., C. S. HALLPIKE “Nerve” deafness: its clinical criteria, old and new. Proc. Roy. Sot. Med. 42 : 527~536,rw. ECCLES, J. C. An electrical hypothesis of synaptic 74* and neuro-muscular transmission. Am. NGI~ York Amt. SC. 47 : 42Q-4SS, 1946. ELDREDGE, D. H,, H. 0. PARRACK AND H. DAVIS. Some responses of the ear to intense highfrequency sound. Pedwation PIOG. 9: 37, 1950. D. H., H. 0. PARRACK AND H. 76. ELDREDGE, DAVIS. Some responses of the ear to intense highfrequency sound. U. S. Air Force Tech. Rep. no. 6564, 1951, pp. I-II H. Morphological studies on the pos77* ENGSTRGM, sible origin of cochlear microphonics. Rev. taryng. 76: -16, r955. H. AND J. WERSilLL. StNCtUI'e of 78. ENGSTRGM, organ of Corti; outer hair cells, Acta oto-tafyng. 43: I-IO, 1953. -79. ENGSTRGM, H. AND J. WERSALL. Structure of Nctwofihysiot. 6: 39-S7,1943. GALAMBOS, R. AND H. DAVIS. Action potential from single auditory-nerve fibers? scicnc6 108: 513, w& GALAMBOS, R., W. A. ROSENBLITH AND M. R. ROSENZWEIG. Physiological evidence for a cochleocochlear pathway in the cat. Exparientia 6: &l-w, 1950. VON GIERKE, ears of humans Fcdwation l IOI. H. E. Subharmonics and animals at intense generated in sound levels. Proc. 9: 130, 1950. GISSELSSON, L. Experimental investigation into the problem of humoral transmission in the cochlea. Acia oto-taryng. suppl. 82 : 1-78, IQSO. GISSELSSON, L. Effect of acetylcholine-esterase inhibiting substances on muscles of middle ear and on latency of cochlear potentials. Acta oto-Zaryng. 42: 208-218, x9.52. GISSELSSON, L. Effect of oxygen lack and decreased blood pressure on microphonic response of cochlea. Acta oto-taryng. 44: 101-118, 1954. GISSELSSON, L. AND 0. RICHTER. Ein Beitrag sur Frage des H&vermogens bei Verschluss des runden Fensters. Arch. Ohree Nascu- Y. Kt?htko#jh. 166: 410-418, 1955. HALLOWELL 46 T. Physical basis of action of cochlea. PIOC. Roy. SOG. Med. 135(B): 490’498,1gg8. 103. GOLD, T. AND R. J. PUMPHREY. The cochlea as a frequency analyzer. Proc. Roy. Sot. Med. 135 (B) : b-m, ~4. 10q. GOLDSTEIN, R. Analysis of summating potential in cochlear responses of guinea pigs. Am. J. Physiot. membrane. 102. GOLD, 178: 331-337, 19% 10s. GOT'O, S., K. YAMAMOTO AND H. NISHIKAWA. phenomena of the auditory Electrical organ. Nogoyu J. M6d. SC. 15(o):gI-g$ 195% 106. VON GRAF, K. Histologische Ver%nderungen des Innenohres nach Behandlung der Miningitis Tuberculosa mit Streptomycin. Acta otu-kyug. 39: 121-131, 19% 107. VON GRAF, der Perilymphe. K. AND G. PORETTI. Die Entstehung Pratt. -oto-rhin-Zoryvq. 12 : 351-365, 19so. R., C. R. SKOGLUND AND S. THESLEFF. Activation of muscle spindles by aucciuyIchoIine and decamethonium. E&&s of curare. Acta physioi. scandkav. 28: 1:34-ISI, Igs. GROSS, N. B. Effects of cochlear lesions on auditory response of guinea pig. J. Camp. & Physiot. PsychoC. 45: 127-39, r952. GRUNDFEST, H., C. Y. KAO AND M. ALTAMIRANO. Bioelectric effects of ions microinjected into the giant axon of Loligo. J. Cm. PhysioC. 38: 245-282, x954. GRUNDFEST, H. In: Electvochemistry in Biotogy ad ktuiiGin6, edited by T. Shedlovsky. New York: Wiley, 195s. GUILD, S. R. War deafness and its prevention. J. Lab. & Ctin. Med. 4: I~-I&, Igrg. GUILD, S. R. Circulation of endolymph. Am. J. m&GRANIT, rag. IIO. 1108. III. rrra. Anat. 39: 112. GUILD, 57-81, 1927. S. R. Effects on hearing of partial sections of nerve in man. Acta oto-larp. 43: 199-207, cochlear 113. HALLPIKE, C. S. Mgnibre’s 31: 330-340, disease. Postgrad. 673, 1053. J. E., Jn. AND M. KNIAZUK. Recovery nerve action potentials after masking. Scit?nct 111: $57-#3, 1950. 120. HAWKINS, J. E., JR. AND M. H. LURIE. of streptomycin. 7&+9, An*. Otof. Rltin. Ototoxicity & Luryng. 61: 1952. 121. HAWKINS, J. E., JR. AND M. H. LURIE. of dihydrostreptomycin and neomycin Am. otoc. Sot. 41: 203-213, x953. HILDING, A. C. Origin and insertion Ototoxicity in cat. Tr. 122. of tectorial & Larmg. 61: 354-370, HILDING, A. C. A theory on the stimulation of the organ of Corti by sound vibration. Am. OtoZ. Rhin. & Larpg.61:371-383,x9$2. McGraw-Hill, 1952. 127. HIRSH, I. J. AND R. C. BILGER. Auditory-threshold recovery after exposure to pure tones. J. Acoust. SOG. Am. 27: 1186=119q, rgss. 118. HOOD, J. D. Studies in auditory fatigue and adaptation. Acto oto-taryng. suppl. g2 : r-57, rgso. 129. HOOD, J. D. Fatigue and adaptation of hearing. Brit.M.J. x2:125-1304956. 130. HUGGINS, W. H. A phase principle for complexfrequency analysis and its implications in auditory theory. J. Acoust. Sot. Am. 24: 582-589, 195% 131. HUGGINS, W. H. A theory of hearing. U. S. Air Force Tech. Rep. no. $3-14, 1953, pp. 1-124. 132. HUGGINS, W. H., AND J. C. R. LICKLIDER. Place mechanisms of auditory frequency analysis. 1. Awutt. Soc.Am.23: 2gw299,19~1. 133. JUUL, A. Electrobiological IS. and histological studies on localization of cochlear microphone effect. Actu oto-tarmg. suppl. 74: rop106, x948. JUUL, A. AND G. VRAA-JENSEN. Effect of ascaridole on inner ear. Actu phwmaud. et to&oC. 3: 5X-72, 1947. 135. KAHANA, L., W. A. ROSENBLITH AND R. GALAMBOS. Effect of temperature change on roundwindow responses in hamster. Am. J. Physiol. 163 : 213-233, 137. 138. 1950. KATSUKI, Y. AND W. P. COVELL. The organ of Corti by phase contrast microscopy. Laryngoscope 63: I-17, 1953. KATSUKI, Y. AND H. DAVIS. Electrophysiological studies of ear of kangaroo rat (Dipodomys). J. Newophysiot. 17: 308-316, 1954. KIMURA, R. AND PERLMAN, H. B. Extensive venous obstruction of the labyrinth. An*. Otol. Rhi~. & Larmg. 65 : 332-350, 1956. KIMURA, R. S., H. F. SCHUKNECHT AND S. SUTTON. Effects of cochlear lesions on threshold responses of auditory cortex in chronic experiments. J. Cum& & Physid. PsychoC. 49: gkoq, 1956. KOSTOLIJK, P. J. Theo&s of &wing. Leiden: Universi taire Pers Leiden, 1950. KRE JCX, F. Experimentelle Untersuchungen tiber den Einfluss des Sympathicus auf die cochleare Funktion. Arch. Ohrcn- Nasen- u. Kehlkopfh. 163: 279-280, 1953 1950. 119. HAWKINS, of auditory Rbin. 124. HILDING, A. C. The tectorial membrane in the theory of hearing. Ann. Otot. Rhin. & Lurpg. 62: 757-769, 1953. 115. HIND, J. E. An electrophysiological determination of tonotopic organization in auditory cortex of cat. J. Ntwrophysioi. x6:475-&, 1953. 126. HIRSH, I. J. Meusrrremwt of l&wing. New York: M. 1. 116. HARRIS, J. D. AND A. 1. RAWNSLEY. Patterns of cochlear adaptation at three frequency regions. J. Acoust. Sot. Am. 25: 760764, x953. 117. HARRIS, J. D. AND A. I. RAWNSLEY. Locus of short duration auditory fatigue of “adaptation”. 1. Exper. Psychot. 46: 457-461, 1953. 118. HAWKINS, J. E., Jn. Cochlear signs of streptomycin intoxication. J. Pharmacol. & Exfier. Thcrafl. IOO: 38-44, 123. 19SS. 114. HALLPIKE, C. S. AND H. CAIRNS. Observations on the pathology of MCni&re’s syndrome. J. L4rpg. 6. Otut. 53 : 625-655, 1~38. 115. HARRIS, J. D. Recovery curves and equinoxious exposures in reversible auditory fatigue following stimulation upto 14o db plus. &rryn~oscups 63: 660- AH~. Oiol. X9$2* 136. 1953a Vdume 37 DAVIS l KREJCI, F. AM) H. BORNSCHEIN. Knalltrauma und ca-Senke beim Meerschweinchen. Pratt. otorkin-laryng. IO: r-8, 1950. KREJCI, F. AND H. BORNSCHEIN. Der Einfluss chronischer uirms&tdigung auf die Cochlearpoten tiale von Meerschweinchen. Acta: oto-Zaryng. 39: -9, 19% KREJCI, F. AND H. BORNSCHEIN. Tierexperimentelle Untersuchungen uber die Cochlearfunktion bei endokraniel er Drucksteigerung. Pratt. oto-rhinlarpg. 13: 146&6,195x. Jmbiwy 1957 BIOPHYSICS AND PHYSIOLOGX 145. KRE JCI, F. AND H. BORNSCHEIN. Cochlear microphonic potentials during sympathetic stimulation. Acta oto-taqysg.qrc: x54-136,1954. 146. KRYTER, K. D. Effects of noise db Haaring Disordws, Monograph 1950. 147. KUCHARSKI, W. Schwingungen in einer pulsierenden Flussigkeit. 264-280, on man. J. S#ech suppl. I : x-gs, von Membranen Physih. 2. 31: IWO. 148. KUFFLER, S. W. AND C. EYZAGUIRRE. Synaptic inhibition in an isolated neme cell. J. Gmr. PhysioZ. 39: us-184,19SS* IN. LANDGREN, S., G. LILJESTRAND AND Y. ZOTTERMAN. Chemical transmission in taste fibre endings. Ada ghysio2. scadinau. 30: x05-114,Ig$4. LAWRENCE, M. AND C. L. BLANCRARD. Prediction of susceptibility to acoustic trauma by determination of threshold of distortion. I&&. Med. and Swg. 23: 1gp200, x9$4. ISI. LAWRENCE, M. AND E. G. WEVER. Effects of oxygen deprivation on structure of organ of Corti. A.M.A. Arch. Otolarpg. $5: 31-37, 1~52. 152. LEDOUX, A. Les liquides labyrinthiques. Acta oto-rhin-latmg. belg. 4: 216-223, 1950. 153. LEMPERT, J., E. G. WEVER AND M. LAWRENCE. The cochleogram and its clinical application. Arch. Otolaryng. 45: 6147, 1947. 154. LEMPERT, J., E. G. WEVER AND M. LAWRENCE. Are the membranous walls of the endolymphatic labyrinth permeable? Tr. Am. Acad. Oghth. $8: 46~ 150. 465, I$$. Iset. LEMPERT, J., D. WOLFF, J. H. T. RAMBO, E. T. WEVER AND M. LAWRENCE. New theory for correlation .of patholosr and the symptomatology of M&&e’s disease. A##, OtoZ. Rhb. & Laryng. 61: 717-737s 1952. 156. LERCHE, E. Masking (electrophysiological effect in animal experiments studies of the inner ear). PjEugcrs Arch. g6s. Physiol. 225: 417-448, rgs2. 157. LICKLIDER, J. C. R. A duplex theory of pitch perception, Ex#rietbtia 7: 128-134, 1951; also ‘Periodicity” pitch and “place” pitch. J. Acoust. Sot. Am. 26: 945, x9$4. 158. LINDSAY, J. R., H. F. SCHUKNECHT, W. D. NEFF AND R. S. KIMURA. Obliteration of the endolymphatic sac and the cochlear aqueduct. A**. Otol. Rhin. & Lary#g. 61: 697-716, rgsa. t$g. LING, G. AND R. W. GERARD. The normal membrane potential of frog sartorius fibers. J. CeZJ. & Comfi. Physiot. 34: 383-396,x940. rho. LURIE, M. H., H. DAVIS AND J. E. HAWKINS, Jn. Acoustic trauma of organ of Corti in guinea pig. Laryngoscop6 54: 375-386, 1944. 161. DE MARE, G. Audiometrischc Untcrsuchungefl. Goteborg: Elanders Boktryckeri Aktiebolag, 1939. 162. MEYER, M. F. Crucial experiments in cochlear mechanics. Am. J. Psychol. 66: 261-26g,rg$3. 162a. MEYER, M. F. How We Hear. Boston: C. T. Branford, xg$o. 163. MITARAI, G. AND YAGASARI, Y. Receptor potential in carp retina. Tr. XIV Jap. Med. Congress, 1g$$, Part II: pp. fQ-30. 164. MYGIND, S. H. Experimental histological studies on labyrinth. Acta otodary?zg. 33: 79-116, IQQ$. 165. MYGIND, S. H. Further labyrinthine studies. Acta oto-Zaryng. suppl. 68: I*, Ig& 166. NACHMANSOHN, D. AND I. B. WILSON. Molecular basis for generation of bioelectric potentials. In: OF INNER EAR 47 Electrochemistry is Biology and Me&&e, edit&l by T. Shedlovsky. New York: Wiley, 1g$$. 167. NEFF, W. D, Effects of partial section of auditory nerve. J. Camp. & PhysioC. PsydoZ. 4.t~ 2030215,1947. t67a. OTTOSON, D. AND G. SVAETICHIN. Electrophysiological investigations of frog retina. CoZd S$ri~g Harbor Symposia Quad. Biol. r7 : 169x73, 1952* 168. PERLMAN, H. nomena of the 7x0-718, Igq. 16g. PERLMAN, H. of living blood & Larpg. 64: B. AND T. J. CASE. Electrical phecochlea in man. Arch. OtoCarpg. 34: B. AND R. S. KIMURA. Observations vessels of cochlea. AM. Otof. Rhin. rt76rxg2, rg$$. 170. PESTALOZZA, G. AND H. DAVIS. Electrical responses of guinea pig to high audiofrequencies. Am. J. PhysioC. 18s: $g$+oo, x956. 171. PETERSON, L. C. AND B. P. BOGERT. A dynamical theory of the cochlea. J. Acoust. SOL Am. 22: 3693% wso. 172. PLOT& E. AND H. B. PERLMAN. A histochemical study of the cochlea. Luryfigosco@ 65: 291-312, rg$$. 173. PORTMANN, G., M. PORTMANN AND L. J. BARBE. Etude exp&imentale de l’action des ultrasons sur l’audition. Acta oto-Zarwg. suppl. IOO: rrg133, 1951. 174. PROSSER, C. L. Comparative physiology of newous systems and sense organs. Am. Ras. Physid. 16: X03-124, 1954. 175. PUMPHREY, R. J. Hearing. Symposia SOG. EZ~GT. Biol. IV. New York: Acad. Press, xg$o. 176. PUMPHREY, R. J. AND T. GOLD. Transient reception and the degree of resonance of human ear. N&sue 160: 124-125, 1947. 177. RAMBO, J. H. T., D. WOLFF AND G. FREEMAN. A research study of the effect of the autonomic nervous system on the internal ear. Tr. Am. Oto. Sot. 41: 244-268, x9$3. 178. RANKE, 0. F. Das Massenverh<nis zwischen Membran und FlUssigkeit im Innenohr. A&wt. 2. 7: I-II, Ig42. x79. RANKE, 0. F. Theory of operation of the cochlea: a contribution to the hydrodynamics of the cochlea. J. Acoust. Sac. Am. 22: 772-777, rgso. I&. RANKE, 0. F., W. D. KEIDEL AND H. G. WESCHKE. Cochleaeffekt bei Verschluss des runden Fensters. Akust. Beih. 3: 14501$8,1g$2. 181. RANKE, 0. F., W. D. KEIDEL AND H. G. WESCHKE. Die seitlichen Beziehungen zwischen Reiz und Reisfolgestrom (Cochlea-Effekt) des Meerschweinchens. 2. Biof. 105: 3&3go,xg$3. 182. RASMUSSEN, G. L. Olivary peduncle and other fiber projections of superior olivary complex. J. Camp. Neuro1. 84: 141-220, rg46. 183. RASMUSSEN, G. L. Further observations of aerent cochlear bundle. J. Com#. Ncurol. 99: 61-74~1953. I&. REBOUL, J. ThCorie des phbnom&nes mCcaniques se passant dans l’oreille interne. J. Phys. Rudism g: 18$-rg4,rg38. 185. REED, C. I. AND W. J. DAY. Do potentials of metabolic origin exist in the synovialis? Am. J. Phydol. 178: 135-137, 1954a 186. REGER, S. N. AND D. M. LIERLE. Changes in auditory acuity. Tr. Am. Ad. Opkth. $8: 433-438, 1954. DENNIS AND H. 187. REHM, W. S., W. H. SCHLESINGER. Electrical resistance of mammalian stomach. Am. J. Pkysiol. x81: 451-470, xg$$. HALLOWELL 48 188. RIESCO-MAcCLURE, J. S., H. DAVIS, B. E. GERNANDT AND W. P. COVELL. Ante-mortem failure of aural microphonic in guinea pig. PIOC. Suc.Exper. BioE.& Med. 71: I@-160, 1949. X~Q. RISKAER, N., E. CHRISTENSEN, I’. V. PETERSEN AND H. WEISMAN. Ototoxicity of neomycin. Ada Otdary?tg. 46: x37-152, 1956. rg0. DE ROSA, L. A. A theory as to the function of the Scala tympani in hearing. J. Acolrst. Sot. Am. IQ: 623628, 1947. 191. ROSENBLITH, W. A. Auditory masking and fatigue. J. Acod. .%?c. Am. 22: 792-800, 1950. 192. ROSENBLITH, W. A. Electrical responses from auditory nervous system. Ann. Otol. Rhin. & Laryng. 63: 839-86% 1954. 193. ROSENBLITH, W. A., R. GALAMBOS AND I. J. HIRSH. Effect of exposure to loud tones upon animal and human responses to acoustic clicks. Science nr: S%-S7& xoso* 194. ROSENBLITH, Electrical electrode W. A. AND M. R. ROSENZWEIG. responses to acoustic clicks: Influence of location in cats. J. Acoust. Sot. Am. 23: 583-588, 1951. 195. ROSENBLUETH, A. AND J. V. LUCO. Local responses of myelinated mammalian axons. 3. CecI. & camp. PhysioC. 36: o&331, 1950. 196. ROSENBLUETH, A. AND J. GARCIA RAMOS. Local responses of mammalian spinal roots to alternating sinusoidal currents. J. Cell. & Covzp. Physiol. 38: 321-345, 1951. 197. ROSENBLUETH, A. AND J. GARCIA RA&IOS. Further study of local responses of axons. J. Cell. & Cm@. Physiol. 39: 109-145, 1952. 198. ROSENZWEIG, M. R. AND W. A. ROSENBLITH. Some electrophysiological correlates of perception of successive clicks. J. Acosut. SOC. Am. 22 : 878-880, 199. ROSENZWEIG, M. R. AND W. A. ROSENBLITH. Responses to successive auditory stimuli at cochlea and at auditory cortex. Psychol. Monographs: General and Applied. 67: 1-26, 1953. 200. RmDI, L. Some animal experimental findings on the functions of the inner ar. AM. OtoZ. Rhin. & Larpg. 60: 993-1016, 1951. 201. RUEDI, L. AND W. FURRER. Das Abstische Tratima. Basel: Karger, 1947. 202. RtkDI, L., W. FURRER, F. LUTHY, G. NAGER AND B. TSCHIRREN. Further observations concerning toxic effects of streptomycin and quinine on auditory organ of guinea pigs. Laryngoscope 62 : 1952. 203. SAXI?& A. Some observations on the physiology of the endolymph stream. Acta oto-ZaryHg. suppl. 74: 87-102, 194% 204. SCHUKNECHT, Tr. Am. Acad. 205. SCHUKNECHT, 65: 402-49, H. F. Lesions of the organ of Corti. Ophth. 366-383, 1953. H. F. Presbycusis. Laryngoscupe r955. 206. SCHUKNECHT, H. F. Ablation therapy for relief of M&i&e’s disease. Laryngoscope 66: 859-870, 1956. da. SCHUKNECHT, H. S. AND R. S. HIMURA. Functional and histological findings after obliteration of the periotic duct and endolymphatic sac in sound conditioned cats. Laryngosco~e 63: 117~1192, 1953. 207. SCHUKNECHT, H. F. AND W. D. NEFF. Hearing losses after apical lesions in the cochlea. Ada otolaryng.42: VOlwn6 37 PERLMAN. An experimental study of auditory damage following blows to the head. A%s. Olol. Rlrin. & LWy?Lg. 60: 273-289, 1951. 209. SCHUKNECHT, H. F. AND S. SUTTON. Hearing losses after experimental lesions in basal coil of cochlea. AXA. Arch. Otolaryng. 57: 129-142,19~3. 210. SCHUKNECHT, H. F. AND R. C. WOELLNSR. Hearing losses following partial section of cochlear nerve. Larmgoscope 63 : 441-465, 1953; also, An experimental and clinical study of deafness from lesions of co&ear nerve. 3. Laryng. & Otot. 49: 75-97,1955. 2x1. SEYMOUR, J. C. AND J. W. TAPPIN. Effect of sympathetic stimulation upon cochlear microphonic potentials. Proc. Roy. SW. Med. 44: 755-759, 1951. 212. SHEDLOVSKY, T. Electromotive force from proton transfer reactions: a model for bioelectric phenomena. Cold Spring Harbor Symposia Quant. Biot. 27: 97102,X952. 213. SHEDLOVSKY, T. (Ed.). Electrochemistry in Biology ad Medicine. New York: Wiley, 1955. 214. SHIMAMOTO, T., H. YAMASAKI, M. INOUE AND ‘I’. KONISHI. A proposal on the cause of M&i&e’s syndrome and its experimental basis. PTOC. Jap. Acad. 30: IO&1011, 1954. 215. SKOUBY, A. I? AND K. ZILSTORFF-PEDERSEN. Influence of acetylcholine, menthol and strychnine on taste receptors in man. Acta fihysiot. scandinarr. 34: 250-256, ~55. 216. SMITH, C. A. Capillary areas of cochlea in guinea pig. Laryngoscope 6x : 1073-10~5, 1951. 217. SMITH, C. A. Electron microscopic studies of organ of Corti. Anat. Rec. 121: 451,1955. 218. SMITH, trolytes C. A., LOWRY, of labyrinthine 14I-G3, 219. SMITH, 1954. C. A., M-L. of endolymph trolytes I950* 333-351, DAVIS 263-274,x952. 208. SCHUKNECHT, H. F., W. D. NEFF AND H. B. 0. H. AND M-L. WU. fluids. tiryngoscupe WU AND 0. H. LOWRY. Elecand perilymph. Scimce 116: 529, 19S2. 220. SMITH, K. R. Histological function of tonal 304-317, 221. SMITH, Elec6.+: frequency. changes in cochlea as a J. Ezpcr. Psychok 37: W47a K. R. AND E. G. WEVER. Functional and histological effects of a high-frequency stimulus. J. EXPGI. Psychol. 39: 238-241, 1949. 222. STEVENS, S. S. (Ed.). Handbook of Exp8timmtaf Psychology. New York: Wiley, 1~51. 223. STEVENS, S. S., J. G. C. LORING AND D. COHEN. Bibliography on Hearing. Cambridge, Mass. : Harvard, 19% 224. STEVENS, S. S. AND H. DAVIS. Hearittg: Its Psychology ad Physiofogy. New York: Wiley, 1938. 225. SUTTON, S. AND H. F. SCHUKNECHT. Regional hearing losses from induced cochlear injuries in experimental animals. Am. Otot. Rhin. & Laryng. 63 : 727753, 226. SUMI, x954* T., Y. KATSUKI AND H. UCHIYAMA. nerve fibers. Pm. Jap. Acad. 32: 67-71, Cochlear 1956. 227. TASAKI, nerve fibers I. Nerve impulses in individual of guinea pig. J. Nclrrophysiol. auditory 16: 97- 122, 1954. 227a. TASAKI, 228. TASAKI, individual stimulation 158,195s. 229. TASAKI, Exploration I. Hearing. Ann. Reu. PhyrioZ. In press. I. AND H. DAVIS. Electric responses of nerve elements in cochlear nucleus to sound (guinea pig). J. Ncuruphysiol. 18: ISII., H. DAVIS AND D. H. ELDREDGE. of cochlear potentials in guinea pig with January 1957 BIOPHYSICS Al?D PHYSIOLOGX a microelectrode. J. Acmst. Sot. Am. 26: 765-773, 1954. TASAKI, I., H. DAVIS AND J-P. LEGOUIX. Spacetime pattern of co&ear microphonics (guinea pig), as recorded by difIerentia1 electrodes. 1. Awust. Sot. Am. 24: sos-5x9, rgS2. I. AND C. FERNhDEZ. M&cations of 231. TASAKI, cochlear microphonics and action potentials by KC1 solution and by direct currents. J. NeuropkytiZ. IS: 497-512, 1952. 232. TONNDORF, J. Analogy between fluid motion within the cochlea and formation of surf on sloping beaches and its significance for mechanism of cochlear stimulation. A##. Otol. &in. & &zfmg. 65: 48&506,1g36. TONNDORF, J. AND F. A. BROGAN. Two forms of 2339 change in co&ear microphonics: parallel shift in stimulus intensity and truncation of gradient curves. USAF School of Aviation Med. Project no. II-27-oo1, Rep. no. 6, 1952. TUNTURI, A. R. Physiological determination of boundary of acoustic area in cerebral cortex of dog. Am. J. PbysioL x60: 3gs-@x, xgs. A. R. Analysis of cortical auditory re235* TUNTURI, sponses with the probability pulse. Am. J. Pbysiot. 181: 630638,rgSS. 236. USSING, H. H. AND K. ZERAHN. Active transport of sodium as source of electric current in short-circuited isolated frog skin. Acta pbysiot. scadi~a. 23: 11o-127, 1951. T., G. VILSTRUP AND C. E. JENSEN. 237e VILSTRUP, On the chemistry of the endolymph. Acta ok-&rwg. suppl. xog: 2oo-2o1, 1933. 238. DL VRIES, HL. Brownian movement and hearing. Pbyticu 14: Ag-60, Igt@. HL. Strulctur und Lage der Tektorial239. DE VRIES, membran in der Schnecke, untersucht mit aeueren Hilfsmitteln. Acta oto-kuyng. 37: 334-338, 1~. DE VRIES, HL. Brownian motion and transmission of energy in cochlea. J. Acoust. Sot. Am. 14: 527333, 249* 250. 251. 252. 253. 256. 2s7* Ill.: 259. 260. 12: 268-280, E. G. Width of basilar membrane in man. Rbin. & Larysg. 47: 37-47, I&. E. G. Tbmy of Eeari?sg. New York: Wiley, E. G., C. W. BRAY AND M. LAWRENCE. of tones in cochlea. J. Acowt. Sac. Am. a61. 26% Ig@. WEVER, 2441 E. G., C. W. BRAY AND M. LAWRENCE. Nature of cochlear activity after death. Am. OtoC. Rhin. & Laryng. $0: 3x7-329, IWI. WEVER, E. G. AND M. LAWRENCE. Tonal interference in relation to cochlear injury. J. Exper. Psychol. 29: 283-295, rgg. 246. WEVER, E. G. AND M. LAWRENCE. Patterns of response in cochlea. J. Awnst. Sot. Am. 21: 127-134, w49. WEVER, E. G. AND M. LAWRENCE. The place principle in auditory theory. Proc. Nut. Ad. SC. 38: 13393% Isw- 263. *. a 266. INNER EAR 49 WEVER, E. G. AND M. LAWRENCE. Sound conduction in cochlea. Ann. Otol. Rhiti. & Laryng. 6x: 824835, 1952. WEVER, E. G. AND M. LAWRENCE. Pkysiokgicul Acowtic~. Princeton, N. J.: Princeton, 19%. WEVER, E. G. AND M. LAWRENCE. Patterns o# injury produced by over-stimulation of ear. J. Acorrst. Sot. Am. 27: 853-858, 1955. WEVER, E. G., M. LAWRENCE AND G. VON BRKI%Y. A note on recent development8 in auditory theory. Proc. Nat. Ad. SC. w: @-~12,rg~. WEVER, E. G.,M. LAWRENCE, R. W.HEMPHILL AND C. B. STRAUT. Effects of oxygen deprivation upon cochlear potentials. Am. J. Pbysioc. rsg: rgga, 1949. WEVER, E. G., M. LAWRENCE AND W. E. RAHM, JR. Phase characteristics of ear. Proc. Nat. Acad. SC. 4.0: 209-218, 19s. WEVER, E. G. AND W. D. NEFF. A further study of etlects of partial section of auditory neme. J. Camp. Physiol. Psychol. w: 217-226, rg47. WEVER, E. G. AND K. R. SMITH. Co&ear impairment as a function of tonal frequency. J. Exptt. Psycbol. 34: 23p245, 1~. WEVER, E. G. AND J. A. VERNON. Sensitivity of turtle’s ear as shown by its electrical potentials. Proc. Nat. Acd. SC. 42: tx312o,rgs6. WILLIAMS, H. L. M&s&6’s Disuasu. Springfield, Thomas, 258. WING, cochlear 19% WEVER, Anrr. otoc. WEVER, w49* WEVER, Interference OF 1952. K. G. A progress report on hypoglycemia and microphonics. USN Submarine Base, Project NM 002 or5.1g.01, Rep. no. 25, 19%. WING, K. G., J. D. HARRIS, A. STOVER AND J. H. BROUILLETTE. Effects of changes in arterial oxygen and carbon dioxide upon cochlear microphonics. 1. Camp. & Pbysiot. PsycboL. 46: 352-357, 1953. WISLOCKI, G. B. AND A. J. LADMAN. Selective staining of otolithic membranes, cupulae and tectorial membrane of inner ear. J. Anot. 8g: 3-12,195s. WOLFF, D. Organ of Corti as we see it today, one hundred years after its discovery. Arch. Otokyng. 56:~88600, w2. WOODBURY, J. W. AND D. M. MCINTYRE. Electrical activity of single muscle cell8 of pregnant uteri studied with intracellular nltramicroelectrodes. Am. J. Pbysiot. 177: 355-360, x954. ZWISLOCKI, J. Theorie der Schneckenmechanik. Acta oto-Zaryng. supp. 72: 1-76, rgq8. ZWISLOCKI, J. Theory of acoustical action of cochlea. 1. Acoust. Sot. Am. 22: 778-784, rgso. ZWISLOCKI, J. Review of recent mathematical theories of cochlear dynamics. J. Acoust. Sot. Am. 25: 743-751, 1953. ZWISLOCKI, J. Wave motion in cochlea caused by bone conduction. J. Acmst. Sot. Am. 25: g86+3g, 1953.