Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Transposable element wikipedia , lookup
Epigenomics wikipedia , lookup
Cell-free fetal DNA wikipedia , lookup
Designer baby wikipedia , lookup
DNA supercoil wikipedia , lookup
Extrachromosomal DNA wikipedia , lookup
Transcription factor wikipedia , lookup
Metagenomics wikipedia , lookup
Nucleic acid double helix wikipedia , lookup
History of genetic engineering wikipedia , lookup
Microevolution wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
Human genome wikipedia , lookup
DNA polymerase wikipedia , lookup
Expanded genetic code wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Long non-coding RNA wikipedia , lookup
Point mutation wikipedia , lookup
Transfer RNA wikipedia , lookup
Short interspersed nuclear elements (SINEs) wikipedia , lookup
Non-coding DNA wikipedia , lookup
RNA interference wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Genetic code wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
Messenger RNA wikipedia , lookup
Polyadenylation wikipedia , lookup
Nucleic acid tertiary structure wikipedia , lookup
RNA silencing wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Deoxyribozyme wikipedia , lookup
RNA-binding protein wikipedia , lookup
History of RNA biology wikipedia , lookup
Epitranscriptome wikipedia , lookup
1. Genes and RNA
The initial products of all genes is a sequence of ribonucleic acid
(RNA).
RNA is produced by a process that copies the nucleotide sequence in
DNA. Since this process is reminiscent of transcribing (copying)
written words, the synthesis of RNA is called transcription.
The DNA is said to be transcribed into RNA, and the RNA is called
a transcript.
One way to think about the different biological roles of DNA and
RNA is to consider that the DNA (that is, the genome) is the
instruction manual for producing all the RNAs that the cell needs,
whereas RNA is the erasable readout of those parts of the manual
relevant to any given task.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
2. Properties of RNA
Although RNA and DNA are both nucleic acids, RNA
differs in several important ways:
1. RNA is a single-stranded nucleotide chain, not a double helix. One
consequence of this is that RNA can form a much greater variety of complex
three-dimensional molecular shapes than can double-stranded DNA.
2. RNA has ribose sugar in its nucleotides, rather than deoxyribose. As the
names suggest, the two sugars differ in the presence or absence of just one
oxygen atom. Analogous to the individual strands of DNA, there is a
phosphate-ribose backbone to RNA, with a base covalently linked to the 1
position on each ribose.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
3. Uracil instead of thymine
The nucleotides of RNA carry the
bases adenine, guanine, and cytosine,
but the pyrimidine base uracil
(abbreviated U) is found in place of
thymine:
However,
uracil forms
hydrogen bonds with
adenine just as thymine does.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
4. Classes of RNA
RNAs can be grouped into two general classes:
Some RNAs are intermediaries in the process of decoding genes
into polypeptide chains; these molecules are called
"informational" RNAs.
In the other class, the RNA itself is the final, functional product.
These RNAs are called "functional" RNAs
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
5. Informational RNAs
For the vast majority of genes, the RNA is only an intermediate in
the synthesis of the ultimate functional product, which is a protein.
The informational RNA of this vast majority of genes is always
messenger RNA (mRNA).
In prokaryotes, the transcript, as it is synthesized directly from the DNA (the
primary transcript), is the mRNA. In eukaryotes, however, the primary
transcript is processed through modification of the 5’ and 3’ ends and removal
of pieces of the primary transcript (introns). At the end of this pre-mRNA
processing, an mRNA is produced.
The sequence of nucleotides in mRNA is converted into the sequence
of amino acids in a polypeptide chain by a process called translation.
In this connection the word translation is used in much the same way
as we use it in translating a foreign language: the cell has a way of
translating the language of RNA into the language of polypeptides.
Proteins are made up of one or more polypeptide chains.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
6. Functional RNAs
Functional RNAs action is purely at the level of the RNA; they are
never translated into polypeptides. Each class of functional RNA is
encoded by a relatively small number of genes (a few tens to a few
hundred). The main classes of functional RNAs contribute to various
steps in the informational processing of DNA to protein. Two classes
of functional RNAs are found in all organisms:
Transfer RNA (tRNA) molecules act as transporters that bring amino acids to
the mRNA during the process of translation (protein synthesis). The tRNAs are
general components of the translation machinery; they can bring amino acids to
the mRNA of any protein-coding gene.
Ribosomal RNAs (rRNAs) are components of ribosomes, which are large
macromolecular assemblies that act as guides to coordinate the assembly of the
amino acid chain of a protein. Ribosomes are composed of several types of
rRNA and about 100 different proteins. As in the case of tRNA, the rRNAs are
general translational components that can be used to translate the mRNA of any
protein-coding gene.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
7. One DNA strand is the template
Transcription relies on the complementary pairing of bases. The two
strands of the DNA double helix separate locally, and one of the
separated strands acts as a template (alignment guide) for RNA
synthesis. In the chromosome overall, both DNA strands are used as
templates, but in any one gene only one strand is used, and in that
gene it is always the same strand.
One or the other DNA strand is used as transcriptional template.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
8. 5’3’
RNA growth is always in the 5’3’ direction; in other words, nucleotides are always
added at a 3’ growing tip:
RNA polymerase moves always from the 3’ end of the template strand, creating an
RNA strand that grows in a 5’3’ direction (since it must be antiparallel to the
template strand). Some genes are transcribed from one strand of the DNA double helix;
other genes use the other strand as the template
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
9. Transcription in action
Transcription of ribosomal RNA (rRNA) genes in
the developing egg cell of the spotted newt
Eukaryotes have several hundred
identical genes encoding
ribosomal RNA. The long
filaments are DNA molecules
coated with proteins. The fibers
extending in clusters from the main
axes are molecules of ribosomal
RNA which will be used in the
construction of the cell's ribosomes.
Transcription begins at one end of
each gene, with the RNA molecules
getting longer as they proceed
toward completion. Note the large
number (up to 100) of RNA
molecules that are transcribed
simultaneously from each gene.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
10. RNA Polymerases
In most prokaryotes, a single RNA polymerase does the job of
transcribing all types of RNA.
Eukaryotes have three different RNA polymerases, which specialize
as follows:
1. RNA polymerase I (Pol I) transcribes rRNA genes.
2. RNA polymerase II (Pol II) transcribes protein-coding genes.
3. RNA polymerase III (Pol III) transcribes other functional RNA genes (for
example, tRNA genes).
In eukaryotes, transcription of nuclear chromosomes takes place
entirely within the nucleus, and the transcripts then move through
nuclear pores out into the cytoplasm, where translation occurs.
Since prokaryotes have no nucleus, there is no comparable movement
of transcripts, and translation can take place immediately, right on the
growing transcript.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
11. Three stages of transcription
Transcription is usually described in terms of three
distinct stages:
Initiation
Elongation
Termination
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
12. INITIATION
A DNA sequence to which RNA polymerase binds to initiate
transcription is termed a promoter.
A promoter is part of the regulatory region adjacent to the coding region of a
gene. Since an RNA transcript is made in the 5’3’ direction, the convention is
to view the gene in the 5’3’ orientation, too (the orientation of the
nontemplate strand), even though transcription actually starts at the 3’ end of the
template strand. By convention the first-transcribed end of the gene is called the
5’ end. Using this view, the promoter is at the beginning of the gene and, so, is
said to be at the 5’ end of the gene, and the regulatory region is called the 5’
regulatory region
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
13. The promoter
Promoter sites have regions of similar sequences, as indicated by the
yellow region in the 13 different promoter sequences in E. coli. Spaces
(dots) included to maximize homology at consensus sequences. The gene
governed by each promoter sequence is indicated on the left. Numbering
is given in terms of the number of bases before () or after (+) the RNA
synthesis initiation point.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
14. The TATA box
Two regions of partial similarity appear in virtually all promoters.
These regions have been termed the -35 (minus 35) and -10 regions
because of their locations relative to the transcription initiation point.
RNA polymerase scans the DNA for a promoter sequence, binds to the
DNA at that point, then unwinds it and begins the synthesis of an RNA
molecule at the transcriptional initiation site. Hence, we see that the
principle of DNA binding is a result of interactions between the
protein (here, the RNA polymerase) and a specific base sequence in
the DNA.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
15. RNA polymerase in bacteria
Schematic diagram of prokaryotic RNA polymerase. The
core enzyme contains two a polypeptides, one b polypeptide,
and one b’ polypeptide. The addition of the s subunit allows
initiation at promoter sites.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
16. The s factor
In order to recognize their promoters, bacterial RNA polymerase enzymes require a
specialized subunit called the sigma factor (σ), which directly contacts the
promoter sequence. The complex formed by the sigma subunit with the remaining
polymerase core subunits constitutes the bacterial holoenzyme.
Bacteria contain a variety of sigma factors that specifically recognize different
promoter sequences. It is therefore the sigma factor that determines which genes are
transcribed.
All cells have a primary sigma factor, which
directs transcription from the promoters of
essential housekeeping genes, and a variable
number of alternative sigma factors whose
levels or activities are increased in response
to specific signals. E. coli, a symbiotic
bacterium leading a relatively sheltered life in
the gut of other organisms, has only 7 sigma
factors.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
17. Structure of a bacterial RNA polymerase
The structure of the T. aquaticus
holoenzyme shows how three structural
domains of the sigma subunit bind to the
core enzyme in a position to recognize the
promoter elements. The DNA is
numbered relative to the transcription start
site at +1. The σ2 domain recognizes the 10 region (red), while the σ3 domain binds
to the flanking base pairs of the extended
-10 region. The σ4 domain, which binds to
the -35 element (red), is anchored to a
flexible flap of the β subunit that may
allow movement of the σ4 subunit to
allow for different spacings between the
-35 and -10 regions.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
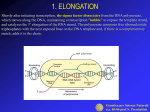
18. ELONGATION
Shortly after initiating transcription, the sigma factor dissociates from the RNA
polymerase, which moves along the DNA, maintaining a transcription "bubble" to
expose the template strand, and catalyzes the 3’ elongation of the RNA strand. The
polymerase compares free ribonucleotide triphosphates with the next exposed base on
the DNA template and, if there is a complementary match, adds it to the chain.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
19. TERMINATION
Specific nucleotide sequences in the DNA act as
signals for RNA chain termination. In the mechanism
called direct termination, the termination signal
consist of about 40 bp containing a GC-rich
palindrome, followed by an oligo A region, which
forms a local stem-loop structure in the RNA. The
resulting double-stranded RNA section is called a
hairpin loop. It is followed by the terminal run of U's
that correspond to the A residues on the DNA
template. This sequence disrupts the base pairing of
newly synthesized RNA with the DNA template,
forcing the RNA and the polymerase to fall off.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
20. RNA Processing in Eukaryotes
Transcription works in much the same way in eukaryotes as in
prokaryotes; that is, there are specific promoter sequences to which
the RNA polymerase binds, and the polymerase moves along the gene
synthesizing RNA in the 5’3’ direction.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
21. The RNA polymerase II holoenzyme
Model of RNA Polymerase II
Transcription Initiation
Machinery.The machinery
depicted here encompasses
over 85 polypeptides in 10
(sub) complexes: core RNA
polymerase II (RNAPII)
consists of 12 subunits; TFIIH,
9 subunits; TFIIE, 2 subunits;
TFIIF, 3 subunits; TFIIB, 1
subunit, TFIID, 14 subunits;
core SRB/mediator, more than
16 subunits; Swi/Snf complex,
11 subunits; Srb10 kinase
complex, 4 subunits; and
SAGA, 13 subunits.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
22. mRNA maturation
In eukaryotes, the initial product of transcription, the primary RNA
transcript, is processed in several ways before its transport to the
cytosol. These processing steps are all performed by specific proteins
that bind to the RNA. Until it reaches its final, mature form, the
primary transcript is sometimes called pre-mRNA.
First, during transcription, a cap consisting of a 7-methylguanosine
residue is added to the 5’ end of the transcript, linked by a
triphosphate bond.
Then an AAUAAA sequence near the 3’ end is recognized by an
enzyme that cuts off the end of the RNA approximately 20 bases
farther down. At this time a stretch of 150 to 200 adenine nucleotides
called a poly(A) tail is added at the cut 3’ end.
Next, a crucial splicing step removes any introns from the RNA
transcript, converting pre-mRNA into mature mRNA.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
23. Mechanism of exon splicing
For many eukaryotic genes the capped and tailed transcripts are shortened by the
elimination of internal introns before transport into the cytoplasm.
Exon-intron junctions of mRNAs show specific sequences that are highly conserved,
i.e., they are the same in most introns in most species. There is a GU at the 5’ splice
site of the intron and an AG at the 3’ splice site in virtually all cases examined ("the
GU-AG rule")
Consensus sequences of 5’ and 3’ splice
junctions in eukaryotic mRNAs. Almost
all introns begin with GU and end with
AG. From the analysis of many exon
intron boundaries, extended consensus
sequences of preferred nucleotides at
the 5’ and 3’ ends have been
established. In addition to AG, other
nucleotides just upstream of the 3 splice
junction also are important for precise
splicing.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
24. The spliceosome
Other less well conserved sequences are found flanking these. These
common configurations of the pre-mRNA are recognized by small
nuclear ribonucleoprotein particles, or snRNPs, which catalyze the
cutting and splicing reactions.
During the process of splicing,
the snRNPs, the primary
transcript, and associated
factors all come together to
form a high-molecular-weight
(60S) ribonucleoprotein
complex, called a
spliceosome, which catalyzes
the splicing reactions.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
25. The translation
At INITIATION, the ribosome
recognizes the starting point in a
segment of mRNA and binds a
molecule of tRNA bearing a
single amino acid. In all bacterial
proteins, this first amino acid is Nformylmethionine.
In ELONGATION, a second
amino acid is linked to the first
one. The ribosome then shifts its
position on the mRNA molecule,
and the elongation cycle is
repeated.
At TERMINATION, when the
stop codon is reached, the chain
of amino acids folds
spontaneously to form a protein.
Subsequently, the ribosome splits
into its two subunits, which rejoin
before a new segment of mRNA is
translated.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
26. Ribosomes are protein factories
The meeting place for amino acid bound tRNAs and mRNA is the ribosome.
Ribosomes are large macromolecular assemblies acting like complex subcellular
machines. Each ribosome consists of a large and a small subunit, which shows slight
differences between prokaryotes and eukaryotes. Each subunit in turn is composed
of several rRNA types and as many as 50 proteins.
Ribosomes contain specific sites that enable them to bind to the mRNA, the tRNAs,
and other specific protein factors, all required for protein synthesis.
The addition of a single
amino acid to the
growing polypeptide
chain in the course of
translation of mRNA
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
27. The tRNAs
The structure of an alanine tRNA,
showing the aminoacyl-tRNA
binding to its correct codon in
mRNA. Some nuclotides carry
rare modified bases
The basis for the specificity between codon and
amino acid lies in the structure of transfer RNA
(tRNA) molecules.
A molecule of tRNA has a clover-leaf shape
consisting of four double-helical stems and three
single-stranded loops.
The middle loop carries a nucleotide triplet called
the anticodon, whose job it is to bind with a
specific codon in the mRNA by specific RNA-toRNA base pairing. Since codons in mRNA are
read in the 5 3direction, anticodons are oriented in
the 3 5 direction.
Each tRNA is specific for only one amino acid and
carries that amino acid attached at its free 3 end.
Amino acids are added to the tRNA by enzymes
called aminoacyl-tRNA synthetases. Each amino
acid has a specific synthetase that links it to only
those tRNAs that recognize the codons for that
amino acid.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini
28. Chain termination
Three codons of the genetic code, UAG, UGA, and UAA, do not
specify an amino acid. These are called stop codons or termination
codons. They can be regarded as punctuation marks ending the
message encoded in the mRNA. Stop codons often are called
nonsense codons.
The three stop codons are not recognized by a tRNA, but instead by
protein factors called release factors. When the peptidyl-tRNA is in
the P site, the release factors bind to the A site in response to the chain
terminating codons. The polypeptide is then released from the P site,
and the ribosomes dissociate into two subunits, ending translation.
In one way of analyzing DNA sequences to look for potential genes,
computers are programmed to look for open reading frames (ORFs),
which are long DNA sequences beginning with an initiation codon
(for example 5-ATG-3) and ending with one of the three stop codons.
Genetica per Scienze Naturali
a.a. 03-04 prof S. Presciuttini