Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Tissue engineering wikipedia , lookup
Cytoplasmic streaming wikipedia , lookup
Cell growth wikipedia , lookup
Cell culture wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell encapsulation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cell nucleus wikipedia , lookup
Cell membrane wikipedia , lookup
Signal transduction wikipedia , lookup
Cytokinesis wikipedia , lookup
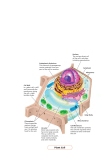
CELL STRUCTURE I Diane L.M. Hickson-Bick, Ph.D. Goals and Objectives: 1. Describe the structure and function of the major components of the cell. 2. Understand the molecular arrangement of cell organelles and associated structures. Key Words: Cytoplasm, plasma membrane, mitochondria, organelle, peroxisome, lysosome. Cells: Structural units of living organisms. A mass of protoplasm containing a nucleus partitioned from the environment by a selectively permeable phospholipid/protein membrane. > 200 different cell types in humans. All share common features: usually a nucleus containing nucleoplasm surrounded by cytoplasm. CYTOPLASM A matrix (cytosol) containing organelles, the cytoskeleton and deposits of carbohydrates, lipids and proteins; separated from the extracellular environment by the plasma membrane (plasmalemma). Plasma Membrane (PM) A limiting membrane composed of phospholipids, cholesterol, proteins and oligosaccharide chains. It has several roles, essentially: Acts as a selective barrier allowing material into and out of the cell thereby regulating the intracellular environment distinct from the extracellular environment. Functions in recognition, regulation and controls interactions with the extracellular environment. Structure of the PM 7.5-10nm thick so only visible by EM Electron lucent layer sandwiched between inner and outer electron-dense lines = unit membrane Roughly 45% lipid, 50% protein and 5% carbohydrate. Cholesterol is present and can increase the fluidity of the membrane by breaking up the orderly packing of the long fatty acid chains of phospholipids. Phospholipids are categorized according to the base that forms the hydrophilic head group, e.g. phosphatidylcholine, phosphatidylserine, phosphatidylethanolamine. Phospholipids are most stable when the hydrophobic fatty acid side chains are directed towards the center of the membrane whilst their hydrophilic head groups are directed outward towards the aqueous environment. The lipid composition of each half of the bilayer is different. Glycolipids possess oligosaccharide side chains that extend outward from the surface towards the extracellular milieu. Protein components are either integral i.e. immersed in the lipid bilayer, or peripheral i.e. lie adjacent to the PM surface. Integral Proteins: Usually transport proteins (e.g. Na+K+ ATPase), ion selective channels, receptors or anchoring structural proteins.Some proteins can span the bilayer whilst others are only partially embedded. Polysaccharide may project from the external surface of integral proteins and form a cell surface coat, the glycocalyx. The presence of carboxyl groups and sulphate ions means the glycocalyx bears an overall negative charge. Activities of the PM Endocytosis: Mass uptake of material through the PM. Exocytosis: Bulk release of material through the PM. Pinocytosis: “Cell drinking”. Small invaginations of the PM entrap extracellular fluid and anything soluble in this fluid. Pinocytotic vesicles pinch off and eventually fuse with lysosomes. Phagocytosis: “Cell Eating” Certain cells are specialized for removing foreign cells, bacteria, fungi, damaged cells etc. Cytoplasmic processes extend and surround the target, the edges fuse and enclose it in an intracellular phagosome. Membrane Trafficking: During endocytosis parts of the PM become endocytic vesicles; during exocytosis the membrane is returned to the cell surface. Receptor-Mediated Endocytosis: Receptors on cell surface bind a ligand causing the receptors to accumulate in coated pits. The cytoplasmic surface of the PM at these points contain a protein called clathrin which forms a lattice meshwork The coated pit invaginates and pinches off from the PM forming a coated vesicle which contains the receptor with a bound ligand The vesicles then lose their clathrin coat and fuse with endosomes. Endosomes are a system of vesicles and tubules located within the cytosol near the cell surface (early endosomes) or deeper within the cell (late endosomes). Clathrin recycles to the PM. Endosomes contain ATP-driven proton pumps within their membranes so the pH within the fused vesicles drops causing dissociation of ligand and receptor complexes. Receptors may recycle to the PM while the ligand is now engulfed in a late endosome which can fuse with lysosomes and complete digest the ligand. Figure 2—4. Schematic representation of the endocytic pathway and membrane trafficking. Ligands, such as hormones and growth factors, bind to specific surface receptors and are internalized in pinocytotic vesicles coated with clathrin and other proteins. After the liberation of the coating molecules, the pinocytotic vesicles fuse with the endosomal compartment, where the low pH causes the separation of the ligands from their receptors. Membrane with receptors is returned to the cell surface to be reused. The ligands typically are transferred to lysosomes. The cytoskeleton with motor proteins is responsible for all vesicle movements described. MITOCHONDRIA Spherical or filamentous, o.5-1μm wide and up to 10μm long Abundance depends on energy demands of cell Composed of an outer (OMM) membrane and an inner membrane (IMM). The IMM projects folds or cristae into the interior of the mitochondrion increasing the surface area. Two functional spaces, the intermembrane space and the matrix Amorphous matrix is protein rich and contains DNA, tRNA, rRNA and mRNA. Also location of the enzymes of the citric acid cycle and fatty acid β-oxidation Mitochondrial membranes have large protein complexes. IMM associated with enzymes and other components of the electron transport chain. Globular structures projecting into the matrix from the IMM represent the ADP to ATP phosphorylating system. Many mitochondrial proteins transcribed in the nucleus and imported into the mitochondrion. RIBOSOMES Small, electron-dense particles, 20 x 30 nm in size. 4 types of rRNA and approx. 80 proteins In eukaryotic cells the ribosomes are composed of two rRNA molecules. Protein components synthesized in the cytoplasm, rRNA in the nucleus. Intensely basophilic because of numerous phosphate groups. Stain with basophilic dyes like hematoxylin, toluidine blue and methylene blue. Individual ribosomes held together by a mRNA strand to form polyribosomes. Ribosomes decode the mRNA message for translation into protein. Proteins synthesized for use within the cell are made on polyribosomes within the cytoplasm. Proteins to be secreted or stored a synthesized on polyribosomes associated with the endoplasmic reticulum (see later notes). ENDOPLASMIC RETICULUM (ER) Anastomosing network of interconnecting channels and sacs formed by a continuous membrane which encloses a space called the cisterna. Two Types of ER: 1. Rough ER (RER): Prominent in cells specialized for protein secretion. Parallel stacks with flattened cisternae. Rough alludes to the presence of ribosomes on the cytosolic side of the ER. Rough ER functions to segregate proteins destined for export (see figure 2-8 above). mRNA’s for these proteins have a 20-25 amino acid sequence on the 5’ end which interacts with proteins and RNA and allows translated sequence to traverse the membrane and enter into the cisterna. 2. Smooth ER (SER): Lacks ribosomes and appears smooth and glandular. Cisternae more tubular. SER is continuous with RER. SER associated with steroid hormone and phospholipid synthesis, degradation of certain hormones. In muscle cells SER is specialized for its role in the sequestration and release of Ca2+ during contraction/relaxation. This specialized SER is called the sarcoplasmic reticulum (SR). GOLGI APPARATUS Completes post-translational modification of proteins, packages and directs them to their ultimate destination. Composed of polarized, smooth, membrane-limited cisternae. Budding of vesicles shuttles newly synthesized proteins from one face towards the other. cis face (convex) receives vesicles from RER Proteins released in vesicles from the trans face (concave). Specific enzymes and proteins are located at different locations within the Golgi. LYSOSOMES Sites of intracellular digestion. Membrane limited containing many hydrolytic enzymes and acidic pH. Very abundant in cells with phagocytic activity e.g. macrophages. Spherical, 0.05-0.5μm in diameter. Lysosomal membranes and proteins packaged in trans-Golgi network and delivered to late endosomes forming endolysosomes. These then mature into lysosomes. Primary lysosomes: Quiescent or not involved in digestion. Secondary lysosome: Primary lysosome fused with a phagosome, 0.2-2μm in diameter with heterogenous appearance (heterophagy). Primary lysosome can fuse with a cytoplasmic organelle needing digestion (autophagy). Residual Body: After digestion most soluble material diffuses out of the secondary lysosome. Indigestible material remains in vacuolar structures = residual bodies. End products of both auto- and heterophagy is often a yellow pigment, lipofuscin. This can be seen with a light microscope and increases with age. PROTEOSOMES Multi-proteases complexes that digest proteins targeted for destruction by ubiquination. Responsible for moving excess proteins or proteins that are incorrectly folded. PEROXISOMES Spherical 0.5-1.2 μm in diameter Oxidize organic materials by transferring hydrogen atoms to molecular oxygen Resulting hydrogen peroxide is toxic and removed by catalase present in peroxisome. β-oxidation of long chain and branched fatty acids occurs here. SECRETORY VESICLES OR GRANULES Found in cells that store a product until its release. Membrane bound Secretory vesicles containing digestive enzymes are called zymogen granules. CYTOPLASMIC INCLUSIONS Usually transitory components containing accumulated metabolites Lipids: Spherical droplets of triglyceride, non-membrane bound. Carbohydrate: Non-membrane bound glycogen deposits. Single 20-30nm particles (β glycogen) or rosettes (α glycogen) THE CYTOSKELETON A complex network of microtubules, actin filaments and intermediate filaments. Provides shape and allows movement of organelles and intracytoplasmic vesicles within cell. Functions in movement of whole cells. Microtubules Cylinders composed of tubulin found in any orientation within the cytoplasm Converge on centrosome and terminate near centrioles Conspicuous in dividing cells. Subunit is a heterodimer of α and B tubulin each has a mass of about 50KD. Tubulin subunits dimerize under the correct conditions Play role in cell shape maintenance, development and intracellular transport. Microtubules form basis for many complex cytoplasmic structures including: Centrioles: Cylindrical bodies (015 x 0.3-0.5μm) of short mictrotubules. Each centriole is comprised of 9 sets of microtubules arranged in triplets. Near nucleus in non-dividing cells is the centrosome which is a pair of centrioles with the long axis of each at right angles to each other. Before cell division each centrosome duplicates, during mitosis the centrosomes move to opposite poles where they become organizing center for the microtubules of the mitotic spindle. Primary Cilia: Found on virtually every cell. Very important during development. Nonmotile, no central pair of microtubules and lack dynein (9+0 arrangements). Very short and do not always project beyond the cell surface. Believed to act as receptors e.g. in the kidney mechanoreptors detecting fluid flow. Several diseases, ciliopathies, stem from defects in primary cilia. Cilia and Flagella: Motile processes covered by plasma membrane. Highly organized with microtubular core. Ciliated cells possess large numbers each 2-3 μm long. Flagellated cells (spermatozoa) have only one flagellum ~100μm long. Cilia and flagella have same core organization, 9 peripheral + 2 central pairs of microtubules = Axoneme. Protein bridges (nexins) link adjacent peripheral pairs. Microtubules of each pair not identical. A= a microtubule of 13 heterodimers with dynein (ATPase) arms; B = a microtubule of 10 heterodimers. At the base of each cilia or flagellum is a basal body that is essentially similar to a centriole and controls assembly of the axoneme. Microfilaments (Actin Filaments) Present in muscle cells as 5-7nm in diameter filament of globular subunits arranged in a double stranded helix. Intimately associated with the contractile process. Actin filaments present in all cells, especially near the PM but the type of actin can vary. Important in defining cell shape, motility and movement of organelles. Microfilaments form the terminal web in the apical cytoplasm of some epithelial cells. ATP hydrolysis required for the polymerization of actin monomers. 1. Intermediate Filaments Filaments 10-12nm in diameter. Several different proteins associated with these proteins, dependent on cell type Filament Type Cell Type Examples Function Keratins Epithelium Keratizing and nonTerminate in desmosomes and keratizing epithelia sites of cell-cell adhesion. Bundles visible under light microscope called tonofilaments Vimentin Mesenchyme Fibroblasts, Attach nucleus to plasma chondroblasts, membrane. macrophages. Desmin Muscle Striated and smooth Provides framework for the muscle attachment of contractile proteins. Glial fibrillary Glial cells Astrocytes (Leydigs) Mechanical strength of cells? acidic proteins Neurofilaments Neurons Nerve cell bodies and Internal support for neuron processes processes and maintaining gel state of cytoplasm. Cell Structure II Diane L.M. Hickson-Bick, Ph.D. Reading: Gartner and Hiatt Chapter 1; Gartner and Hiatt, Chapter 1, (Or Junquiera Basic Histology), Chapter 3; Klein and McKenzie pp 47-58 and look at first 3 chapters too Objectives: Understand the arrangement and function of the components of the cell’s nucleus. Understand the elements of cell division and the cell cycle. Describe the multiple forms of cell death. Describe the function and composition of cell surface specializations Describe composition and function of different intercellular junctions. Key Words: Nucleus, chromatin, cell division, cell death, cell cycle, microvilli, intercellular junctions, basement membrane, basal lamina. THE CELL NUCLEUS The nucleus houses the genetic machinery. It does not synthesize protein. Usually a round or elongated structure, often at the center of the cell. Main components are: nuclear envelope, chromatin, the nucleolus, and nuclear matrix. Rat Pituitary Cell EM n = nucleolus NE = nuclear envelope N = Nucleoplasm GA = Golgi apparatus rER = rough endoplasmic reticulum. Nuclear Envelope 2 parallel unit membranes separated by a space called the perinuclear cisterna. Impermeable to ions and proteins of all sizes. Internal membrane associated with the fibrous lamina = 3 main proteins, lamins A, B and C. In non-dividing cells chromosomes are associated with the fibrous lamina. Polyribosomes attached to outer membrane. Inner and outer membranes fuse at nuclear pore. Nuclear pores provide pathways between nucleus and cytoplasm. Complex protein structures ~150nm in diameter; >100 proteins. Chromatin Coiled strands of DNA bound to basic proteins (histones) Two types of chromatin distinguishable in non-dividing cells. Heterochromatin = electron dense and appears as coarse clumps in electron microscope or basophilic clumps by light microscopy. Composed of unexpressed DNA Euchromatin = Less coiled portions of chromosomes and visible as finely dispersed granular material. Site of active transcription. In dividing cells chromosomes become supercoiled and densely staining Human cell has 22 pairs of chromosomes grouped by size and position of the centromere and 2 sex chromosomes = diploid state. In female cells one X chromosome remains euchromatic while other remains condensed and inactive. Nucleolus Spherical structure up to 1mm in diameter. Rich in rRNA and protein All protein components synthesized in the cytoplasm and imported. 3 basic components seen by EM: 1) Nucleolar organizer DNA: Base sequences coding for rRNA 2) Pars Fibrosa: Densely packed ribonucleoprotein fibers, primary transcripts of rRNA genes 3) Pars Granulosa: 15-20nm granules representing maturing ribosomes. CELL DIVISION (MITOSIS) A parent cell divides into two daughter cells each of which receives a chromosome set identical to the parent cell. INTERPHASE: The phase between two mitoses. Its duration can vary greatly. DNA replication occurs here. PROPHASE: Gradual coiling of chromatin, chromatin condenses to form chromosomes of 2 chromatids bound at the centromere; microtubules disintegrate; centrosomes with centrioles separate and migrate to opposite poles of the cell; nucleolus and nuclear membrane disperse. METAPHASE: Chromosomes migrate to equatorial plane where each divides longitudinally to form 2 chromosomes called sister chromatids. Chromatids attach to the microtubules of the mitotic spindle at the electron dense kinetochore. ANAPHASE: Sister chromatids separate and migrate to opposite poles of the cell pulled by microtubules. Centromeres move away from the center. TELOPHASE: Nuclei reappear in daughter cells, chromosomes uncoil and nuclear membrane and nucleolus reform. Cleavage furrow develops between daughter cells. CELL CYCLE Cell Cycle = Interphase + 4 phases of mitosis. Interphase can be divided into 3 phases: G1(presynthesis) S (DNA synthesis) G2 (post-DNA duplication). Cell cycle can be temporally or permanently suspended. Such cells are referred to as being in G0. CELL DEATH APOPTOSIS: Programmed cell death. Energy requiring process. Helps remove injurious cells from an organ. Complex pathways can be activated ultimately resulting in a compacted (pyknotic) nucleus, specific cleavage of the chromatin, blebbing of the plasma membrane. No rupture of the PM to release intracellular components into extracellular space therefore no activation of the immune system ONCOSIS/NECROSIS: Accidental death caused by pathological processes. Cells swell and release intracellular components into extracellular space stimulating macrophages and downstream immune responses often leading to inflammation CELL SURFACE SPECIALIZATIONS Many cells are polarized with an apex (top), lateral (side) and basal (bottom) domains. MICROVILLI Fingerlike projections (1 x 0.1μm) on the apical surface of cells Increase surface area of cells by up to 25X. Common on epithelial cells but especially on absorptive cells where it has a thick glycocalyx. Microvilli + glycocalyx = Brush Border Core of actin filaments linked by other proteins. Actin filaments extend into the body of the cell where they form a network, the terminal web. PRIMARY CILIA,CILIA and FLAGELLA – Principle protein tubulin. See previous notes from CELL STRUCTURE I. STEREOCILIA Misleading name. Long non-motile microvilli (actin core) found on epithelial cells of the epididymis and hair cells of the inner ear. Absorptive function in epididymis and sensory function in the ear. MICROPLICAE Fold like extensions of cytoplasm present on luminal epithelial cells in places like the cornea, esophagus and renal collecting ducts. Increase surface area and the adherence of surface fluid layers. INTERCELLULAR JUNCTIONS Structures that contribute to cohesion and communication between cells. Very prominent in epithelia subjected to traction and pressure. Tight Junctions (Zonula occludens) Most apical junction, divides apical and basolateral domains. Forms a band completely encircling cell. Membrane fusion between adjacent cells closes off intercellular spaces. Anastomosing series of ridges and grooves forms a netlike structure. Number of fusion sites correlates to relative leakiness of an epithelium. Intermediate Junctions (Zonula Adherens) Basal to tight junctions Actin filaments arising from the terminal web insert into electron dense plaques on the surface of junctional membranes. Gap Junctions Found in most tissues along the lateral membranes of cells Represent a close (2 nm) apposition of two adjacent membranes. Aggregates of intramembrane particles in circular patches Gap junction protein units called connexins. These form hexamers with hydrophilic pores in the center (connexons) Connexons project through plasma membrane and join in mirror symmetry with connexons in adjacent cells. Site of exchange between cells for electrical coupling and cell-cell communications. Molecules >1000 Daltons cannot pass through. Desmosomes (Macula Adherens) Complex disk-shaped structures at the surface of one cell directly opposite an identical structure on an adjacent cell. Cell membranes at these junctions usually very straight. On cytosolic sides tonofilaments converge and form hair-pin loops near the dense plaques of the desmosome. Plaques contain many protein including vimentin, desmin and cadherins Rows of desmosomes lie basal to the intermediate junctions of absorptive or secretory epithelial cells. Numerous desmosomes in protective or multilayered epithelia like skin. Hemidesmosomes Basal structure = Half a desmsome. Anchor cell by type VII collagen to the subjacent basal lamina (see below). Proteinaceous plaques in these structures contain many integrins. Membrane Infoldings Deep infoldings of the basal membrane which partition the cytoplasm into alcoves Alcoves often contain many mitochondria. Increase basal surface area for active pumping of solute to generate an extracellular osmotic gradient. Found in epithelial cells of the renal system, ducts of sweat glands and ciliary process of the eye. BASAL LAMINA AND BASEMENT MEMBRANE Basal Lamina: A sheet-like extracellular structure at the basal surface of epithelial cells where they contact the subjacent connective tissue. Visible by EM as an electron dense layer 20-100nm thick = the lamina densa. Lamina densa contains type IV collagen and laminins. An electron-lucent layer may be present on one or both sides of the lamina densa. This is the lamina rara (lamina lucida). Basement Membrane: Visible with light microscope. Formed by either the fusion of: (A) two basal laminae OR (B) a basal laminar and a reticular laminar (laminar reticularis). (A): The fusion of a basal lamina formed by an epithelial cell and a basal lamina of an endothelial cell layer. Especially visible in the kidney glomerulus. And in the alveoli of the lung. Arranged as a thick lamina densa with a lamina rara on either side. (B). More common form of basement membrane composed of a basal lamina plus a lamina reticularis. Lamina reticularis is formed by fibroblasts subjacent to the basal lamina. Lamina reticularis contains fibronectin and type III collagen. Type VII collagen fibrils bind the basal lamina to the subjacent collagen. ___________________________________________________________________________ CELL STRUCTURE LAB – DEMO SLIDE OF MITOSES at front of lab! SLIDE 1 – LIVER – H&E Tissue is embedded in paraffin and Hematoxylin (blue-purple) and eosin (red) show. Hematoxylin binds to RNA and DNA, eosin binds to proteins. Look at the large, round/polygonal cells, which are hepatocytes. The cytoplasm stains pink and the nucleus blue. Try to find these: Plasma Membrane Nuclear envelope Cytoplasm Euchromatin Heterochromatin Nucleolus Rough endoplasmic reticulum Look for a yellow substance. This is housed within lysosomes. What’s this stuff called? SLIDE 2 – EPIDIDYMIS Locate the cells lining the cut tube (lumen) of the epididymis (Hollow round structures). These are the epithelial cells. Locate the nucleus and the luminal (apical) cell membrane. Between the membrane and nucleus is an area that is poorly stained and looks ‘washed out’. This is the GOLOGI complex. Next look at the apical surface and projecting into the lumen there are some long extensions. These are misnamed STEREOCILIA; they are NOT cilia but actually long microvilli. What protein is in the core of these structures? Look for small dark dots at the lumen where two cells meet. This is the TERMINAL BAR and it corresponds to the intermediate junction and is visible with a light microscope. SLIDE 4 – SMALL INTESTINE Find the luminal surface. Go to higher power, and look at the apical surface where there should be a pink line; this is the BRUSH BORDER and is formed by millions of MICROVILLI. How are these microvilli anchored into the cell? SLIDE 12 – TRACHEA Once again go to the apical/luminal surface. You should find some ‘eye-lash’ like extensions from the surface. These are motile CILIA. What’s in the core of these structures? MICROSCOPE HINTS Carry by the arm and support the base Plug it in (Don’t laugh, this often happens) Place slide on stage with coverslip facing UP Start with the low magnification objective. Lower the lens with the coarse knob. Do NOT use oil! Bring slide into focus while looking through the eyepieces. Check to see if you have a pointer on your microscope….just so you know. Make sure that you have a clear view through BOTH eyepieces. Go from 4X to 10X and up to 40X. The slide should remain in focus (for the most part) even after you have moved the lens. Use the fine adjustment knob for small changes. DO NOT USE THE 1OOX LENS AND IMMERSION OIL. THE OIL GUMS UP THE WORKS AND WE ONLY USE OIL FOR THE BLOOD LABS. IT IS SAID: LOW POWER FOR HIGH POWER MINDS, HIGH POWER FOR LOW POWER MINDS