Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Single-unit recording wikipedia , lookup
Synaptogenesis wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Electrophysiology wikipedia , lookup
Multielectrode array wikipedia , lookup
Neural coding wikipedia , lookup
Nervous system network models wikipedia , lookup
Development of the nervous system wikipedia , lookup
Apical dendrite wikipedia , lookup
Chemical synapse wikipedia , lookup
Neuroanatomy wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Synaptic gating wikipedia , lookup
Subventricular zone wikipedia , lookup
Anatomy of the cerebellum wikipedia , lookup
Circumventricular organs wikipedia , lookup
Optogenetics wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
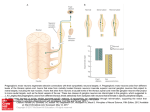
1301 The Journal of Experimental Biology 202, 1301–1309 (1999) Printed in Great Britain © The Company of Biologists Limited 1999 JEB2092 PHYSIOLOGY OF ELECTROSENSORY LATERAL LINE LOBE NEURONS IN GNATHONEMUS PETERSII YOSHIKO SUGAWARA1,*, KIRSTY GRANT2, VICTOR HAN3 AND CURTIS C. BELL3 1Department of Physiology, Teikyo University School of Medicine, Kaga 2-11-1, Itabashi-ku, Tokyo 173-8605, Japan, 2Institut Alfred Fessard, Centre National de la Recherche Scientifique, 91190 Gif-sur-Yvette, France and 3Neurological Sciences Institute, Oregon Health Science University, Portland, OR 97209, USA *e-mail: [email protected] Accepted 16 February; published on WWW 21 April 1999 Summary In mormyrid electric fish, sensory signals from to the ELL molecular layer in vitro evokes an excitatory electroreceptors are relayed to secondary sensory neurons postsynaptic potential (EPSP), generally followed by an in a cerebellum-like structure known as the electrosensory inhibitory postsynaptic potential (IPSP), in the efferent lateral line lobe (ELL). Efferent neurons and interneurons neurons. In MG cells, the same stimulation evokes an of the ELL also receive inputs of central origin, including EPSP, often followed by a small IPSP. Synaptic electric organ corollary discharge signals, via parallel fibers transmission at parallel fiber synapses is glutamatergic and and via fibers from the juxtalobar nucleus. To understand is mediated via both N-methyl-D-aspartate (NMDA)- and the cellular mechanisms of the integration of sensory inputs (AMPA)-type glutamate receptors. The inhibitory and central inputs in the ELL, the intracellular activity and component of the parallel fiber response is GABAergic. It ionic properties of the efferent projection neurons and is probably mediated via the stellate neurons and the MG interneurons were examined in an in vitro slice cells, which are themselves GABAergic interneurons preparation. intrinsic to the ELL network. We focus here on the electrophysiological properties of A hypothetical neural circuit of the intrinsic connections the efferent neurons of the ELL network, the large of the ELL, based on the known morphology of projection fusiform cells and large ganglion cells, and on a class of γneurons and medium ganglion interneurons, is presented. aminobutyric acid (GABA)-ergic interneurons known as This circuit includes an excitatory and an inhibitory medium ganglion (MG) cells. In response to current submodule. The excitatory submodule is centered on a injection through a recording pipette, both types of efferent large fusiform cell and appears to relay the sensory input neuron fire a large narrow spike followed by a large as a positive ‘ON’ image of an object. The inhibitory hyperpolarizing afterpotential. The MG cells fire a complex submodule is centered on a large ganglion cell and relays spike which consists of small narrow spikes and a large a negative ‘OFF’ image to the next higher level. We suggest broad spike. Although the forms of the action potentials in that MG cells exert an inhibitory bias on efferent neuron efferent neurons and in MG cells are different, all spikes types and that the ELL network output is modulated by the are mediated by tetrodotoxin (TTX)-sensitive Na+ dynamically plastic integration of central descending conductances and spike repolarization is mediated by signals with sensory input. tetraethylammonium (TEA+)-sensitive K+ conductances. In the presence of TEA+, substitution of Ba2+ for Ca2+ in the bath revealed the presence of a high-voltage-activated Key words: mormyrid, electric fish, electrosensory lateral line lobe, corollary discharge, Na+-dependent dendritic action potential, Ca2+ conductance. excitatory submodule, inhibitory submodule, Gnathonemus petersii. Stimulation of parallel fibers conveying descending input Introduction Weakly electric mormyrid fish use their electric sense for electrolocation and electrocommunication by detecting the electrical current flow through specialized cutaneous electroreceptors generated by the fish’s own electric organ discharge (EOD). Three classes of electroreceptor are present: mormyromasts, which serve in active electric imaging of the environment; ampullary receptors, which are sensitive to low-frequency electric events in the water; and Knollenorgans, whose primary role is communication. Centrally projecting fibers from electroreceptors terminate in the electrosensory lateral line lobe (ELL) (Maler, 1973; Bell and Russell, 1978a). Primary afferents from mormyromast and ampullary electroreceptors terminate in separate zones of the cortex of the ELL (Bell et al., 1989), while primary 1302 Y. SUGAWARA AND OTHERS afferents from Knollenorgan receptors terminate in the nucleus of the ELL (nELL) (Bell and Grant, 1989). All three classes of electroreceptor respond to the fish’s own EOD and to stimuli from the environment (Szabo and Hagiwara, 1967; Bell and Russell, 1978b; Bell and Szabo, 1986). The self-induced afferent signals are known as ‘reafferent’ signals (von Holst and Mittelstaedt, 1950), and their significance to the animal is different in each of the three electrosensory pathways. In the context of the Knollenorgan and ampullary electroreceptive pathways, reafferent input may potentially cause interference with the reception of exafferent electrosensory signals (those originating from the environment, rather than being a result of the animal’s ‘autostimulation’). For the mormyromast electrosensory pathway, however, reafferent electrosensory input is of prime interest to the active imaging of the environment. Each pathway-specific zone of the ELL also receives corollary discharge input associated with the motor command that drives the electric organ (electric organ corollary discharge, EOCD) (Zipser and Bennett, 1976; Bell, 1982). Thus, with each EOD, the neurons of the ELL are affected both by reafferent electrosensory signals and by descending EOCD signals. The interactions between reafferent and EOCD signals are different in each of the electrosensory zones of the ELL (Bell and Szabo, 1986). Those interactions revealed in studies in vivo are briefly described as follows. Afferent signals from Knollenorgan receptors encode information about the timing of brief electrical signals, such as the EODs of other fish, and evoke excitatory postsynaptic potentials (EPSPs) in the neurons of the nucleus of the ELL (Bell et al., 1989). The corollary discharge to the nELL induces a brief and precisely timed inhibitory postsynaptic potential (IPSP) in the nELL neurons, which completely blocks the electrosensory-reafferent-driven EPSP (Zipser and Bennett, 1976; Russell and Bell, 1978; Szabo et al., 1979). Complete blockade lasts for only 1–2 ms, but this brief corollary discharge inhibition is sufficient to block the reafferent response and to remove the interference that would have been caused by reafferent input (Bell and Grant, 1989). Ampullary electroreceptors are specialized for measuring low-frequency electrical signals (Kalmijn, 1974). Afferent fibers from ampullary electroreceptors project to the ventrolateral zone of the ELL (VLZ in Fig. 1). Primary afferents discharge tonically with a frequency that may be modulated up or down by external stimuli. The fish’s own EOD causes an acceleration followed by a deceleration of the tonic discharge rhythm. A brief EOCD inhibition, such as in the Knollenorgan region, would not be effective in inhibiting reafferent ampullary input. The corollary discharge input to the ampullary zone evokes a more complex, temporally structured series of excitatory and inhibitory events which selectively filter the longer-lasting reafferent input (Bell, 1981). Thus, under stable conditions, the corollary discharge effect counterbalances or minimizes sensory reafferent inputs. In the mormyromast electrosensory pathway, reafferent responses convey actively sought information about the environment. Mormyromast electroreceptors serve active electrolocation by measuring the intensity of current flow induced by the fish’s own EOD. Object images are generated by the distortions that they impose on the current flow within the fish’s own electric field. Repetitive association of reafferent electrosensory signals with the descending EOCD signal SM Voltage LC Current injection SD EGp PF MZ DLZ Fig. 1. Diagram of a transverse section of the electrosensory lateral line lobe (ELL) to show the positions of the recording and stimulating electrodes in vitro. DLZ, dorsolateral zone of ELL; EGp, eminentia granularis posterior; LC, lobus caudalis; MZ, medial zone of the ELL; PF, parallel fibers; VLZ, ventrolateral zone of the ELL; SD, SM, recording electrodes in the deep fiber and molecular layers, respectively. VLZ Afferents from electroreceptors Physiology of the mormyrid ELL 1303 modulates the effect of the EOCD-driven input to the ELL networks. This plasticity follows an anti-Hebbian learning rule (Hebb, 1949; Bell, 1981, 1990; Bell et al., 1992; Bell and Grant, 1992), such that the EOCD-driven feedback to the ELL generates a negative image of the recent reafferent responses and acts as an adaptive filter of actual or current reafferent responses (Bell and Grant, 1992; Bell et al., 1993, 1997a). One plausible site for the adaptive filter is thought to be at the synapses between parallel fibers and the medium ganglion cells of the ELL (Bell et al., 1997b). Our goals are to understand the adaptive filtering of the electrosensory inputs by descending signals on the basis of the cellular mechanisms of electrogenesis in the ELL neurons, focusing particularly on the mormyromast region. In this paper, we give a brief description of the basic anatomical organization and the electrophysiological properties of the efferent projection neurons and GABAergic interneurons of the ELL cortex. Both in vivo and in vitro intracellular recordings have revealed that the forms of action potentials and the firing properties are different in these cell types. On the basis of evidence obtained from the brain in vivo and from in vitro slices of the ELL, we will discuss a hypothetical model of electrosensory processing in the ELL. Materials and methods The results presented in Figs 2–4 were obtained by using patch recording pipettes in the whole-cell configuration in slices of the ELL. The ELL was excised from Gnathonemus petersii, anesthetized with MS-222 (Sandoz) (100 mg l−1), and sliced at a thickness of 300 µm in low-[Na+] ACSF (see below) in which half the Na+ had been replaced with iso-osmotic sucrose as described previously (Grant et al., 1998). After incubation for 1–2 h in low-[Na+] ACSF, the slice was placed in the recording chamber and perfused with standard ACSF. The composition of standard ACSF was as follows (in mmol l−1): 125 NaCl, 2.5 KCl, 1.25 NaH2PO4, 2.0 CaCl2, 1.6 MgSO4.7H2O, 24 NaHCO3 and 10 glucose, pH 7.2–7.4. The ACSF was bubbled with 95 % O2 and 5 % CO2. Parallel fiber stimulation was applied through a monopolar tungsten wire electrode in the molecular layer. Primary afferent fibers were stimulated with a similar electrode in the deep fiber layer. The responses of ganglion layer neurons of the ELL were recorded using a patch pipette (resistance 10–20 MΩ) filled with an internal pipette solution which contained (in mmol l−1): 140 potassium gluconate, 3.0 KCl, 10 Hepes, 10 EGTA, 1.0 CaCl2, 1.0 MgCl2 and 110 µg ml−1 nystatin (Sigma). The pH was adjusted to 7.3. The response was amplified with a patch-clamp amplifier (Nihon Koden, CEZ-2300), stored on a digital tape recorder (Sony, PC204A) and analyzed offline. The intrinsic network of the ELL: efferent neurons and medium ganglion cells (GABAergic interneurons) The ELL cortex has six layers: the molecular, ganglionic, plexiform, granule, intermediate and deep fiber layers. Primary afferents from the A and B receptor cells of mormyromast electroreceptors terminate in the granule layer of the ELL medial zone (MZ in Fig. 1) and dorsolateral zone (DLZ in Fig. 1) respectively. Sensory signals transmitted to the granule cells are relayed to efferent projection neurons and inhibitory interneurons, including medium ganglion (MG) cells. Large ganglion (LG) cells and large fusiform (LF) cells were recorded in the ganglionic cell layer and in the plexiform or granule layers, respectively (Grant et al., 1996; Meek et al., 1996; Han et al., 1999). The efferent axons of the LG cells and the LF cells project to the preeminential nucleus and to the lateral nucleus of the torus semicircularis. The LF and LG cells, also referred to as E-cells and I-cells, respond to stimuli in the center of their electroreceptive field with increased activity and decreased activity, respectively (Bell et al., 1997a). Medium ganglion, cells are GABAergic interneurons, as shown by immunohistochemistry (Meek et al., 1996) and pharmacology (Grant et al., 1998). The cell bodies of the MG cells are found in the ganglionic cell layer and are of two types: MG1 cells with basal dendrites confined to the plexiform layer and axonal terminals in the ganglion, plexiform and granule layers; and MG2 cells with basal dendrites in the granule layer and axonal terminals confined to the ganglion and plexiform layers (Meek et al., 1996; Han et al., 1999). The MG cells are 7–10 times as numerous as the LG cells (Meek et al., 1996; Han et al., 1999). The EOCD signals from the EOD motor command nucleus arrive by at least two pathways (Bell et al., 1983). The first arises via the paratrigeminal command-associated nucleus, which projects to the eminentia granularis posterior (EGp in Fig. 1). The granule cells of the EGp in turn give rise to the parallel fibers that project to the outer two-thirds of the molecular layer of the ELL (Maler, 1973). The second EOCD pathway arises via the mesencephalic command-associated nucleus, which projects to the juxtalobar nucleus via the juxtaleminiscal region (Bell et al., 1995; Bell and von der Emde, 1995). The cells of the juxtalobar nucleus in turn project to the mormyromast region of the ELL, primarily to the granular layer, although some terminals are also found in the plexiform and ganglion layers (Bell et al., 1995; Bell and von der Emde, 1995). Thus, integration of electrosensory signals with the descending EOCD inputs probably takes place in at least two stages: an early stage, in the granular layer, with the EOCD inputs from the juxtalobar nucleus, which convey accurate information on the timing of the EOD motor command (Bell et al., 1995; Bell and von der Emde, 1995); and a later stage in the molecular layer at the level of the spiny dendrites of the efferent cells and MG cells, which receive parallel fiber input. Parallel fibers convey descending signals from the EGp (Bell et al., 1981, 1995; Bell and von der Emde, 1995; Meek et al., 1996). In the in vitro slice preparation, integration in the molecular layer of the ELL has been examined by applying field stimulation to the parallel fiber that mimics descending input, without the influence of EOCD-driven effects on the early stage in the granule layer (Fig. 1). Action potentials in LG, LF and MG neurons in vitro Intracellular recordings from efferent neurons and interneurons, subsequently identified by labeling with biocytin, 1304 Y. SUGAWARA AND OTHERS A Fig. 2. Action potentials and synaptic responses of efferent neurons to molecular layer stimulation and to current injection through the recording pipette. (A–C) Current injection through the recording pipette evoked bursts of large narrow spikes. Note that the bursting spikes were induced on the depolarizing phase of an oscillatory membrane fluctuation (open arrow). Membrane potential -72 mV was −72 mV. The intensity of the stimulus is shown to the right of each trace. (C) At high intensity, the bursts changed to repetitive D firing. (A) Molecular layer stimulation (SM) evoked an excitatory postsynaptic potential (EPSP) and large narrow spikes, sometimes followed by an inhibitory postsynaptic potential (IPSP) (filled arrow). Two superimposed traces show that the occurrence of an IPSP suppresses the second and third spikes. (E) A single sweep showing an IPSP on an expanded scale. C B 100 pA 255 pA 110 pA 20 mV 50 ms E 10 mV 20 mV SM 20 ms showed that the form of action potentials in these two types of neurons is different in vivo (Bell et al., 1997a) and in vitro (Grant et al., 1998). LG and LF efferent neurons fire large narrow spikes. In results obtained by using a sharp microelectrode in the in vitro slice preparation, Grant et al. (1998) reported that in LG cells these action potentials were 25–50 mV in amplitude (35.7±2.2 mV, mean ± S.E.M., N=25) and 1–2 ms in duration; in LF cells, action potentials were 30–60 mV in amplitude (42.2±3.8 mV, N=15) and 1–2 ms in duration. Large hyperpolarizing afterpotentials follow these spikes in both LG and LF neurons. There was little difference between the forms of action potentials and their large hyperpolarizing afterpotentials in LG and LF efferent neurons. In contrast, intracellular recordings from MG cells in vitro showed two types of spikes: a large broad spike of 25–60 mV in amplitude (49.8±1.7 mV, N=28) and 10–20 ms in duration with a small after-hyperpolarization, and a small narrow spike 1–10 mV in amplitude (5.8 mV ±0.4 mV, N=7) and 1–2 ms in duration. Small narrow spikes also occurred in the absence of the large broad action potentials. Under some conditions, a medium-sized broad spike, 8–20 mV in amplitude and 4–6 ms in duration, was also seen in isolation (Bell et al., 1997a; Grant et al., 1998). This suggests that each component of the complex spike may have a different origin and a different role in cell activities. As described in a previous study (Bell et al., 1993), the expression of plasticity appeared to depend on the activation of a large broad spike in the postsynaptic neuron. Two types of MG neuron were distinguished on the basis of cell morphology, MG1 and MG2 cells. No differences were observed, however, in the recorded electrophysiological properties of these two cell types. Responses around the threshold for firing were examined by using a patch pipette whose tip resistance was lower than that of a sharp microelectrode. In the efferent neurons, current injection through a recording patch pipette evoked a slow SM 20 ms depolarizing potential (open arrow in Fig. 2A). Injection of low-intensity current evoked an oscillatory membrane fluctuation leading to a large narrow spike or to bursts of several spikes. With increased current intensity, the number of spikes in a burst increased and the inter-burst interval decreased (Fig. 2B). Even stronger stimulus intensity evoked a repetitive tonic discharge (Fig. 2C). The latency to the first spike gradually decreased with the increase in current intensity (Fig. 2A,B). Latency is plotted against current intensity in Fig. 3C (open squares). However, the initial frequency, calculated from the interval between the first and the second spikes of the response train, appeared to be fixed (Fig. 3D; open squares). Thus, in efferent neurons, current injection initially evoked an oscillatory membrane depolarization which then induced a large narrow spike. An increase in stimulus current intensity evoked an increase in the amplitude of membrane fluctuation and a shortening of the burst interval. With large depolarizations of more than approximately −50 mV, the burst firing pattern changed from bursting to repetitive firing (Fig. 2C). Therefore, the fixed first interval suggests that the conductances underlying spike generation in efferent neurons may be saturated. Further analysis of the ionic currents involved is required. Molecular layer stimulation in vitro excites parallel fibers and evokes short-latency EPSPs in efferent neurons which are sufficient to trigger large narrow spikes (Fig. 2D). In many efferent cells, IPSPs follow the initial EPSP with a latency of 2–3 ms or more and may be sufficient to block later spikes of the response (Fig. 2D,E). Grant et al. (1998) reported that EPSP–IPSP sequences of this sort were observed in approximately 48 % of the efferent neurons recorded; an EPSP only was obtained in 45 % of efferent cells and an IPSP only was found in 7 % of the efferent neurons. IPSPs recorded in LG and LF neurons may result from parallel fiber excitation of GABAergic stellate cells in the molecular Physiology of the mormyrid ELL 1305 layer, whose axons terminate on the apical dendrites of these neurons, and from the excitation of neighboring MG neurons, whose axons terminate on the somata of LG and LF neurons. In MG cells, current injection through the recording pipette in vitro evoked both small narrow and large broad spikes (Fig. 4A–C). The threshold for initiation of small spikes was always lower than that for the large broad action potentials. The broad spike thus always arose from a small spike (Fig. 4C,D), being initiated increasingly close to the peak of the small spike as current intensity increased. The latency of both spike types decreased with increasing current intensity (Fig. 3C). At high current intensity, the frequency of both small narrow and large broad spikes increased (Fig. 3D), as calculated from the interval between the first and second spikes. This is in contrast to the behavior of the large narrow spikes in efferent neurons. Since the curve showing frequency as a function of current intensity for small spikes showed a steeper slope (filled triangles and circles in Fig. 3D) than that for large broad spikes (open circles and triangles in Fig. 3D), the generation of small spikes seemed to be sensitive to the stimulus intensity. In MG cells, parallel fiber stimulation evoked an EPSP from which several sizes of small spikes and large broad spikes could arise (Fig. 4D,E). When the tungusten stimulating electrode was moved a small distance (100 µm) along the parallel fibers, a small IPSP was evoked at the time that the second small spike arose (Fig. 4F). This supports the view that the MG cells also receive inhibitory inputs from the stellate cells or from neighboring MG cells, as shown in the morphological study of Meek et al. (1996). Voltage-dependent channels in efferent neurons and medium ganglion cells The addition of 0.3 µmol l−1 tetrodotoxin (TTX) to the bath perfusion medium abolished all regenerative spikes in both efferent neurons and medium ganglion cells. This means that Na+ is the primary carrier of the inward current during the large narrow spike, the large broad spikes and the small narrow spikes. In efferent neurons, after suppression of large narrow spikes by TTX, current injection induced large depolarizing plateau potentials when Ca2+ was replaced with Ba2+ in the presence of the K+ channel blocker tetraethylammonium (TEA+; 25 mmol l−1). In medium ganglion cells, after replacement of Ca2+ with Ba2+ and in the presence of TEA+, a long depolarizing potential followed the broad spike. In both cell types, these large, slow potentials were blocked by the addition of 0.1 mmol l−1 Cd2+. Since Ba2+ is a potent charge carrier through Ca2+ channels and Cd2+ suppresses Ca2+ Frequency (Hz) Latency (ms) B A Fig. 3. Relationships First frequency First frequency between the intensity of the stimulus current Latency Latency injected through the patch pipette and spike generation in efferent and medium ganglion (MG) cells. (A) Burst firing of an efferent cell 20 mV 20 mV in response to a 20 ms 50 ms suprathreshold current pulse shown in Fig. 2A,B. Large and small open 500 400 squares show the C 350 D measured points for latency and first 300 100 frequency, respectively. ■■ 250 (B) Responses of an MG 200 cell to current injection. Filled symbols indicate 150 10 small spikes and open 100 symbols indicate large broad spikes. (C) Plot of 50 first spike latency against l 0 1 stimulus current intensity. 50 100 150 200 250 300 350 0 50 100 150 200 250 300 350 0 Open squares indicate Current (pA) Current (pA) large narrow spikes of an efferent cell. Circles indicate one MG cell and triangles indicate a second MG cell. The large narrow spikes in the efferent cells had a lower stimulus threshold than the small narrow spikes or large broad spikes in MG cells. Note that the small narrow spikes in MG cells were evoked with a lower stimulus intensity than the large broad spikes. (D) Plot of initial frequency (the inverse of first interspike interval) against injected current intensity for an efferent cell and two MG cells (symbols as in Fig. 3C). Note that the initial frequency of the small spikes in the MG cells increased with current intensity, while the initial frequency was almost constant in the efferent cell, probably due to an activation of oscillatory membrane fluctuation. 1306 Y. SUGAWARA AND OTHERS Fig. 4. Responses of medium C B A ganglion (MG) neurons to molecular layer stimulation and to current injection through the recording pipette. (A–C) Current injection through the recording pipette evoked bursts of small narrow spikes. Membrane potential was −65 mV. 100 pA 125 pA 175 pA The intensity of the stimulus is shown 20 mV to the right of each trace. (A,B) Low- -65 mV intensity current injection evoked 50 ms small narrow spikes. (C) An increase F E* D in current intensity induced repetitive firing of small narrow spikes and a large broad spike. (D) Low-intensity molecular layer stimulation (SM1) 10 mV 10 mV evoked only a small spike (asterisk). Higher-intensity stimulation evoked * a larger excitatory postsynaptic 20 mV 20 ms 20 ms potential (EPSP) inducing small SM1 SM2 narrow spikes and a large broad 20 ms spike. (E) The response marked with SM1 an asterisk in D shown on an expanded scale. (F) When the stimulating tungsten electrode was moved by 100 µm (SM2), low-intensity molecular layer stimulation induced a small inhibitory postsynaptic potential (IPSP) (arrow). Two traces with and without an IPSP are superimposed. conductances (Hagiwara and Barley, 1981), these results suggest that the slow depolarizing potentials in both types of neuron are induced by activation of a Ca2+ conductance. However, the stimulus threshold for activation of this Ba2+ spike was approximately 3–4 times greater than that for the Na+ spike. The high threshold for these slow potentials suggests the activation of high-voltage-activated Ca2+ channels. The high threshold could also be partly due to a distal distribution of Ca2+ channels in the apical dendrites. Ion channel distribution and function The relative distribution of ion channel densities cannot be established with the available data. However, it appears that voltage-sensitive Na+ channels must be present in the axonal, somatic and proximal dendrite membranes, as in hippocampal pyramidal neurons (Turner et al., 1991; Jaffe et al., 1992), in neocortical pyramidal neurons (Regehr et al., 1993; Stuart and Sakmann, 1994) and in pyramidal neurons of the gymnotid ELL (Turner et al., 1994). ELL efferent neurons have relatively large myelinated axons. Large narrow spikes are probably evoked as propagating spikes in these axons with a low stimulus threshold. The spike then invades the soma antidromically and possibly also the apical dendritic tree. Broad dendritic spikes may be present in the apical dendrites of efferent neurons but are obscured in somatic recordings by the presence of K+ channels. The large after-hyperpolarizations suggest a high density of K+ channels in these efferent neurons. In MG cells, the small narrow spikes are probably evoked in the axon and the large broad spikes probably arise in the soma or proximal dendrites. These suggestions are based on the following reasoning. (1) The small narrow spike has a lower threshold than the large broad spike. Action potential initiation occurs at the point of lowest threshold in the neuron, usually in the initial segment of the axon (Eccles, 1964). This has been demonstrated in the neocortical pyramidal neuron (Stuart and Sakmann, 1994) in which the action potential is initiated in the axon, leading to a somatic action potential that actively invades the apical dendritic tree via activation of dendritic TTX-sensitive Na+ channels. (2) The small size of the narrow spikes in MG cells may be a simple consequence of geometry and ionic channel density, as predicted in the impulse propagation model of Moore et al. (1983). The MG cell has quite a thin axon, which terminates within the ELL (Meek et al., 1996). Because of the small diameter of the axon, its action potential may not generate enough action current to overcome the abrupt increase in load at the soma and will propagate only electrotonically to the soma. The depolarization caused by a small spike is not sufficient to generate a broad spike, which requires additional excitatory synaptic input. The broad spike always arises from a small spike because of its higher threshold. (3) The finding that broad spikes can be recorded extracellularly quite high in the molecular layer in vivo (Bell et al., 1997a,b), distant from the cell bodies in the ganglionic cell layer, also supports the hypothesis that large broad spikes are propagated into the apical dendrites. The pairing of postsynaptic broad spikes with EOCD-driven EPSPs was necessary, in vivo, for the induction of plastic changes in the response to this synaptic input. Similarly, the occurrence of the broad spike is necessary for the anti-Hebbian plasticity at the parallel fiber synapse that has been demonstrated in vitro (Bell et al., 1997b). Physiology of the mormyrid ELL 1307 Comparison of responses to ascending input in vivo and in vitro Primary afferent activation in vitro is achieved by electrical stimulation in the deep layers of the ELL (SD in Fig. 1), whereas in vivo it is achieved by natural electrosensory stimuli. The following description is based on both in vitro (Grant et al., 1998) and in vivo (Bell et al., 1997a) results. Comparison of the two reveals interesting features of the neural network in the ELL. Deep fiber layer stimulation in vitro evoked predominantly IPSPs in efferent neurons. In LF cells, IPSPs were observed in 88 % of the cells tested, and EPSPs were present in 18 % of the cells. In LG cells, IPSPs were evoked in 100 % of cells and EPSPs in 18 % of cells. In the in vivo recordings, however, the response with the lowest threshold for local electrosensory stimuli in LF cells was an EPSP, which evoked a brief burst of spikes at higher intensity. The minimum latency was 3–4 ms. The LF cell in vivo receives predominantly excitatory input via granule cells from the sensory stimulation of the center of the receptive field, and has been classified as an E-cell (excited cell) (Bell et al., 1997a). The response with the lowest threshold in LG cells in vivo was generally an IPSP with a minimum latency of 3–7 ms and a duration of more than 100 ms. The LG cells in vivo receive predominantly inhibitory input via granule cells from the sensory stimulation to the center of the receptive field and correspond to the I-cells (inhibited cell) of the classification of Bell et al. (1997a). Deep fiber layer stimulation in vitro also evoked predominantly IPSPs in MG cells. Of 32 MG cells tested, IPSPs only were recorded in 47 %, EPSPs only were recorded in 9 % and an IPSP–EPSP sequence was recorded in 6 %. Strong deep fiber stimulation in vitro evoked small narrow spikes and also large broad spikes, similar to those evoked by parallel fiber stimulation (Grant et al., 1998). In vivo, electrosensory-evoked IPSPs in most MG cells occurred with minimal latencies of 4–7 ms. Thus, there are several differences between the afferent responses recorded in vivo and in vitro. In particular, electrosensory stimulation in vivo evoked EPSPs in the LF cells, whereas electrosensory stimulation in vitro evoked predominantly IPSPs in these neurons. Although the connection between granule cells and the efferent or MG cells is still not completely understood, these differences may arise because the LF cell receives inputs from both the excitatory and inhibitory granule cells and from inhibitory interneurons; the total number of inputs from inhibitory neurons may be larger than the population of excitatory granule cell inputs. In vitro particularly, it is possible that field stimulation in the deep fiber layer does not reproduce exactly the structure of the projection of peripheral electroreceptive fields and might stimulate a larger population of inhibitory afferent neurons, masking the contribution of the granule cells responsible for the excitatory ‘center’ of the receptive field. The functional network of the ELL The cellular mechanisms underlying the integration in the ELL are not yet completely understood, but some of the roles and interactions of a number of different neurons have been identified. In the following paragraphs, we present a schematic summary sufficient to explain some of the interactions between neurons in the ELL. The hypothetical neuronal circuit presented in Fig. 5 is based on known morphology (Meek et al., 1996; Grant et al., 1996; Bell et al., 1997a,b; Han et al., 1999) and electrophysiological properties studied in vivo (Bell et al., 1997a,b) and in vitro (Grant et al., 1998). However, it also contains several features that are still hypothetical. The circuit includes seven types of neuron: two types of efferent neuron (large ganglion and large fusiform cells); two types of inhibitory Purkinje-like medium ganglion cells (MG1 and MG2); two types of granule cell (one of which inhibits MG1 and LG cells and one of which excites large fusiform and MG2 cells); and stellate cells of the molecular layer. Parallel fibers E-module I-module (EOCD) SC MG1 MG2 OFF image Inhibitory bias LG Inhibitory bias LF ON image Juxtalobar afferents (EOCD) Efferent axons GCs GCs Primary afferents Inhibitory cell and synapse Excitatory cell and synapse Fig. 5. Schematic representation summarizing sensory integration in the electrosensory lateral line lobe (ELL). The two types of efferent neuron (large fusiform and large ganglion cells) are shown within two separate submodules, the E-module and the I-module. Each module also has a medium ganglion cell and granule cells. The medium ganglion cells, which are γ-aminobutyric acid (GABA)ergic neurons, control the activity of the efferent neurons (shown as ‘inhibitory bias’ in the diagram). The descending inputs include EOCD-conveying parallel fibers in the molecular layer and EOCDconveying fibers from the juxtalobar nucleus in the deep part of the ELL. The excitatory module transfers a positive ‘ON’ image of electrosensory inputs to higher levels, whereas the inhibitory module transfers a negative ‘OFF’ image. LF, large fusiform cell; LG, large ganglion cell; MG1 and MG2, medium ganglion cells; GCs, granule cells; SC, stellate cell; EOCD, electric organ corollary discharge (modified from Han et al., 1999). 1308 Y. SUGAWARA AND OTHERS Morphological reconstructions suggest that MG1 cells preferentially inhibit large fusiform cells and that MG2 cells preferentially inhibit large ganglion cells. MG cells are known to terminate on other MG cells, but whether the connections are preferentially reciprocal, with MG1 cells inhibiting MG2 cells and vice versa, is still speculative. The circuit can be understood as a small module that processes electrosensory information from a local region of the skin and that is reproduced repeatedly across the electrosensory maps of the two mormyromast zones. EOCD signals are shown entering the ELL via parallel fibers and via fibers from the juxtalobar nucleus. The diagram in Fig. 5 greatly simplifies the effects of EOCD signals in the ELL. The juxtalobar input probably does more than simply gate the electrosensory input to LF, LG and MG cells at the early stage of integration. It also probably elicits EPSPs and IPSPs in these cells via direct or indirect connections. EOCD signals conveyed via the parallel fibers also integrate proprioceptive information and past electrosensory history in the form of a feedback pathway from the preeminential nucleus. In addition, fibers of the direct preeminential tract originating from the preeminential nucleus terminate in the lower molecular layer of the ELL, also bringing feedback from higher levels of the electrosensory pathway and EOCD-related information. Thus, the late stage of integration in the molecular layer involves EOCD signals, but also concerns feedback from other modalities of sensory information as well as the higher stages of the electrosensory pathway itself. Following the scheme in Fig. 5, electrosensory stimuli in the center of receptive fields will evoke EPSPs in LF and MG2 cells, but will evoke IPSPs in LG and MG1 cells. Cells that are inhibited in the center of their receptive field will receive excitation from other sources, such as the EOCD, or from the periphery of their receptive field. Each efferent cell receives inhibition from a type of MG cell that responds in an opposite manner to the same sensory stimulus. Efferent cell responses will be enhanced by such inhibition. LF cells, for example, will be excited by a stimulus increase in the center of their receptive field, and the same stimulus will inhibit neighboring MG1 cells. Inhibition of the MG1 cell will disinhibit the LF cell, enhancing its excitatory response to the stimulus. The inhibitory control of the efferent neurons exerted by the MG cells is shown as ‘inhibitory bias’ in Fig. 5. The two types of efferent cells and their associated MG cells and granule cells may be viewed as excitatory and inhibitory submodules, as indicated by the outlined shaded areas in Fig. 5. The excitatory submodule (E-module), centered on LF cells, will convey a positive ‘ON’ image to higher levels of the electrosensory system, i.e. an image in which increases in discharge rate signal increases in peripheral stimulus intensity. In this E-module, additive integration in the LF cell will occur between EOCD-driven EPSPs and sensory input. The inhibitory submodule (I-module), centered on large ganglion cells, will convey a negative ‘OFF’ image to higher levels, i.e. an image in which increases in discharge rate signal decreases in peripheral stimulus intensity. In this I-module, subtractive integration in the LG cell will occur between EOCD-driven EPSPs and sensory input. The two types of submodule may thus be compared with the ‘on’ and ‘off’ components of the visual system. Conclusion The mormyromast region of the ELL receives afferent input from mormyromast electroreceptors and relays it to the next higher level: the preeminential nucleus and the lateral nucleus of the torus semicircularis. Electric organ corollary discharges (EOCD) enter by two pathways: via the juxtalobar nucleus projection to the granule cell layer of the ELL and via the EGp and the parallel fiber projection to the molecular layer of the ELL. The early stage of integration in the ELL may occur in the granule cells, which receive the afferent input and EOCDdriven input from the juxtalobar nucleus. The granule cells must have two types of activities, excitatory and inhibitory, depending on their connection with the primary afferents. In response to stimuli at the center of the receptive field, excitatory granule cells will induce EPSPs in the E-module, centered on the efferent LF cell, and will induce IPSPs in the I-module. Stimuli to the periphery of the receptive field will evoke an opposite response for each module. EOCD-driven descending inputs evoke EPSPs in the apical dendrites just before the arrival of the reafferent sensory input. Consequently, in the LF cell of the E-module, EOCD-driven EPSPs and reafferentdriven EPSPs will be summated, leading to additive depolarization which induces a propagating large narrow spike. In the LG cell of the I-module, EOCD-driven input in the apical dendrite evokes an EPSP, but reafferent input will induce subtractive IPSPs which may block initiation of the spike. Thus, the E-module will transfer the positive-going ‘ON’ image, and the I-module will transfer the negative-going ‘OFF’ image to the next higher level. The MG cells, which receive sensory input opposite to that of the efferent cell in each module and also EOCD-driven EPSPs via their apical dendrites, make inhibitory synapses with the efferent neuron in their module and thus control its activity. If plastic changes occur at the EOCDdriven synapses, their effects within the network will be visible as modulation of the output of efferent neurons. This research was supported by grants from the del Duca Foundation and Yamada Science Foundation to Y.S., by grants from the National Science Foundation and the National Institute of Mental Health to C.B. and by contract C11*CT920085 from the European Economic Community, a grant from NATO and funds from the Centre National de Recherche Scientifique to K.G. References Bell, C. C. (1981). An efference copy which is modified by reafferent input. Science 214, 450–453. Bell, C. C. (1982). Properties of a modifiable efference copy in an electric fish. J. Neurophysiol. 47, 1043–1056. Physiology of the mormyrid ELL 1309 Bell, C. C. (1990). Mormyromast electroreceptor organs and their afferent fibers in mormyrid fish. III. Physiological differences between two morphological types of fibers. J. Neurophysiol. 63, 319–332. Bell, C. C., Caputi, A. and Grant, K. (1997a). Physiology and plasticity of morphologically identified cells in the mormyrid electrosensory lobe. J. Neurosci. 17, 6409–6423. Bell, C. C., Caputi, A., Grant, K. and Serrier, J. (1993). Storage of a sensory pattern by anti-Hebbian synaptic plasticity in an electric fish. Proc. Natl. Acad. Sci. USA 90, 4650–4654. Bell, C., Dunn, K., Hall, C. and Caputi, A. (1995). Electric organ corollary discharge pathways in mormyrid fish. I. The mesencephalic command associated nucleus. J. Comp. Physiol. A 177, 449–462. Bell, C. C., Finger, T. E. and Russell, C. J. (1981). Central connections of the posterior lateral line lobe in mormyrid fish. Exp. Brain Res. 42, 9–22. Bell, C. C. and Grant, K. (1989). Corollary discharge inhibition and preservation of temporal information in a sensory nucleus of mormyrid electric fish. J. Neurosci. 9, 1029–1044. Bell, C. C. and Grant, K. (1992). Sensory processing and corollary discharge effects in mormyromast regions of mormyrid electrosensory lobe. II. Cell types and corollary discharge plasticity. J. Neurophysiol. 68, 859–875. Bell, C. C., Grant, K. and Serrier, J. (1992). Sensory processing and corollary discharge effects in the mormyromast regions of the mormyrid electrosensory lobe. I. Field potentials, cellular activity in associated structures. J. Neurophysiol. 68, 843–858. Bell, C. C., Han, V. Z., Sugawara, Y. and Grant, K. (1997b). Synaptic plasticity in a cerebellum-like structure depends on temporal order. Nature 387, 278–281. Bell, C. C., Libouban, S. and Szabo, T. (1983). Pathways of the electric organ discharge command and its corollary discharges in mormyrid fish. J. Comp. Neurol. 216, 327–338. Bell, C. and Russell, C. J. (1978a). Termination of electroreceptor and mechanical lateral line afferents in the mormyrid acousticolateral area. J. Comp. Neurol. 182, 367–382. Bell, C. C. and Russell, C. J. (1978b). Effect of electric organ discharge on ampullary receptors in a mormyrid. Brain Res. 145, 85–96. Bell, C. C. and Szabo, T. (1986). Electroreception in mormyrid fish. Central anatomy. In Electroreception (ed. T. H. Bullock and W. Heiligenberg), pp. 375–421. New York: Wiley-Interscience. Bell, C. and von der Emde, G. (1995). Electric organ corollary discharge pathways in mormyrid fish. II. The medial juxtalobar nucleus. J. Comp. Physiol. A 177, 463–479. Bell, C. C., Zakon, H. and Finger, T. E. (1989). Mormyromast electroreceptor organs and their afferent fibers in mormyrid fish. I. Morphology. J. Comp. Neurol. 286, 391–407. Eccles, J. C. (1964). The Physiology of Synapses. Berlin: SpringerVerlag. 316pp. Grant, K., Meek, J., Sugawara, Y., Veron, M., Denizot, J. P., Hafmans, T. G. M., Serrier, J. and Szabo, T. (1996). Projection neurons of the mormyrid electrosensory lateral line lobe: morphology, immunohistochemistry and synaptology. J. Comp. Neurol. 375, 18–42. Grant, K., Sugawara, Y., Gomez, L., Han, V. and Bell, C. C. (1998). The mormyrid electrosensory lobe in vitro: Physiology and pharmacology of cells and circuits. J. Neurosci. 18, 6009–6025. Hagiwara, S. and Barley, L. (1981). Calcium channel. Annu. Rev. Neurosci. 4, 69–125. Han, V., Bell, C. C., Grant, K. and Sugawara, Y. (1999). The mormyrid electrosensory lobe in vitro. I. Morphology of cells and circuits. J. Comp. Neurol. 404, 359–374. Hebb, D. O. (1949). The Organization of Behavior. New York: Wiley. Jaffe, D. B., Johnston, D., Lasser-Ross, N., Lisman, J. E., Miyakawa, H. and Ross, W. N. (1992). The spread of Na+ spikes determines the pattern of dendritic Ca2+ entry into hippocampal neurons. Nature 357, 244–246. Kalmijn, A. J. (1974). The detection of electric field from inanimate and animate sources other than electric organs. In Handbook of Sensory Physiology, vol. III/3, Electroreceptors and Other Specialized Receptors in Lower Vertebrates (ed. A. Fessard), pp. 148–200. New York: Springer-Verlag, Maler, L. (1973). The posterior lateral line lobe of a mormyrid fish – A Golgi study. J. Comp. Neurol. 152, 281–299. Maler, L., Sas, E., Carr, C. E. and Matsubara, J. (1982). Efferent projections of the posterior lateral line lobe in gymnotiform fish. J. Comp. Neurol. 211, 154–164. Meek, J., Grant, K., Sugawara, Y., Hafmans, T. G. M., Veron, M. and Denizot, J. P. (1996). Interneurons of the ganglionic layer in the mormyrid electrosensory lateral line lobe: morphology, immunohistochemistry and synaptology. J. Comp. Neurol. 375, 43–65. Moore, J. W., Stockbridge, N. and Westerfield, M. (1983). On the site of impulse initiation in a neuron. J. Physiol., Lond. 336, 301–311. Regehr, W., Kehoe, J., Ascger, P. and Armstrong, C. (1993). Synaptically triggered action potentials in dendrites. Neuron 11, 145–151. Russell, C. J. and Bell, C. C. (1978). Neural responses to electrosensory input in mormyrid valvula cerebelli. J. Neurophysiol. 41, 1495–1510. Stuart, G. J. and Sakmann, B. (1994). Active propagation of somatic action potentials into neocortical pyramidal cell dendrites. Nature 367, 69–72. Szabo, T., Enger, P. S. and Libouban, S. (1979). Electrosensory systems in the mormyrid fish, Gnathonemus petersii: special emphasis on the fast conducting pathway. J. Physiol., Paris 75, 409–420. Szabo, T. and Hagiwara, S. (1967). A latency-change mechanism involved in sensory coding of electric fish (mormyrids). Physiol. Behav. 2, 331–335. Turner, R. W., Maler, L., Deerinck, T., Levinson, R. and Ellisman, M. H. (1994). TTX-sensitive dendritic sodium channels underlie oscillatory discharge in a vertebrate sensory neuron. J. Neurosci. 14, 6453–6471. Turner, W. R., Meyers, D. E. R., Richardson, T. L. and Barker, J. L. (1991). The site for initiation of action potential discharge over the somato dendritic axis of rat hippocampal CA1 pyramidal neurons. J. Neurosci. 11, 2270–2280. von Holst, E. and Mittelstaedt, H. (1950). Das Reafferenzprinzip. Wechselwirkungen zwischen Zentralnervensystem und Peripherie. Naturwissenschaften 37, 464–476. Zipser, B. and Bennett, M. V. L. (1976). Interaction of electrosensory and electromotor signals in lateral line lobe of a mormyrid fish. J. Neurophysiol. 39, 713–721.