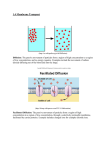
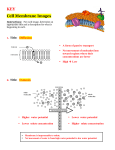
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell growth wikipedia , lookup
Cell culture wikipedia , lookup
Model lipid bilayer wikipedia , lookup
Lipid bilayer wikipedia , lookup
Chemical synapse wikipedia , lookup
Membrane potential wikipedia , lookup
Cell encapsulation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Signal transduction wikipedia , lookup
Cytokinesis wikipedia , lookup
SNARE (protein) wikipedia , lookup
List of types of proteins wikipedia , lookup
Journal of Experimental Botany, Vol. 47, No. 299, pp. 717-728, June 1996 Journal of Experimental Botany REVIEW ARTICLE The measurement of exocytosis in plant cells Nicholas Battey1'3, Andrew Carroll 2 , Pirn van Kesteren 1 , Alison Taylor2 and Colin Brownlee2 1 2 Plant Science Laboratories, The University of Reading, Whiteknights, Reading RG6 6AS, UK Marine Biological Association, Citadel Hill, Plymouth PL12PB, UK Received 3 October 1995; Accepted 12 February 1996 Abstract Exocytosis is of vital importance to the growth and development of plant cells. It is a dynamic process in which vesicles bearing polysaccharide precursors and proteins fuse with the plasma membrane and release their contents. Equally important, new plasma membrane is delivered by exocytosis as secretory vesicle membrane becomes incorporated. The requirements for polysaccharides, proteins and plasma membrane are very different in different cell types, so there must be sophisticated mechanisms for ensuring delivery of these materials to the correct cellular locations at the appropriate time and, particularly in the case of membrane, their recovery and recycling. Currently, little is known of these mechanisms in plants, but new methods for measuring exocytosis are under development, and existing techniques have already contributed data of considerable relevance. Here the methods for measuring exocytosis are described and evaluated, with emphasis on the electrophysiological measurement of capacitance as a relatively non-invasive method, and on cell-free assays because of their potential importance in the identification of proteins and other factors that control exocytosis in plant cells. Key words: Exocytosis, vesicle traffic, vesicle fusion, polysaccharides, cell wall, cell plate, root cap, secretion, patch-clamping. Introduction Exocytosis is the process by which plant cells secrete polysaccharide precursors for cell wall elaboration and hence cell growth. Extracellular proteins also cross the plasma membrane by exocytosis, following synthesis on the endoplasmic reticulum and transport through the 3 To whom correspondence should be addressed. Fax: +44 1734 750630. © Oxford University Press 1996 Golgi apparatus. Further, membrane necessary for plasma membrane growth and for its differentiation during the life of the cell is incorporated from the secretory vesicle membrane during exocytosis. For each of these processes (polysaccharide, protein and phospholipid delivery) there may be alternative routes, but the available evidence suggests that the exocytotic pathway is of major importance. The processes of cell development that depend on exocytosis, and the molecular mechanisms that may be involved in its control have been reviewed elsewhere (Battey and Blackbourn, 1993). The emphasis of this review will, therefore, be on methods for measuring exocytosis in plants. This stress on techniques is justifiable because although the study of exocytosis is in its infancy in plants, a wide variety of methods is available from work on animals and yeast. The presence of a cell wall and high turgor pressure creates problems specific to plant cells so that careful application of these methods is required. Yet it is this wall and the importance of cell growth in plant development that makes quantification of exocytosis of such especial interest in plants. Exocytosis releases secretory product and increases plasma membrane area. Methods that measure either of these processes are, therefore, potentially able to report exocytosis, but because of limitations in both approaches, the ideal solution is to try and estimate secretion and membrane area simultaneously. A good deal of effort is currently focused on the estimation of membrane area by capacitance, so this technique has been given some prominence here. Additionally, cell-free assays in which secretory vesicles and plasma membrane are isolated and then mixed in a fusion assay, are useful as a means to isolate and purify regulators of exocytosis. They may also contribute valuable information on mechanisms, if interpreted with caution and in parallel with measurements of 718 Battey et al. exocytosis in vivo. The opportunities for development of these assays in plant cells have, therefore, been emphasized. Measurement of secretory product release Polysaccharides Changes in secretory product release are likely to be accompanied by changes in the rate of exocytosis. However, there can clearly be many causes of such changes in flux, including alterations in rates of product synthesis, packaging, and vesicular transport through and from the Golgi, as well as in the exocytotic step itself. In the root cap of many plants hypersecretory cells exocytose large quantities of polysaccharide slime (Plate 1). Many measurements of this secretion have been made and there is much evidence for major control at the level of synthesis (as reflected in Golgi activity; Rougier, 1981). However, the analysis carried out by Bowles and Northcote (1974) shows that it is possible to calculate flux specifically for the post-Golgi (presumably exocytotic) step. Radioactivity from 14C-glucose saturates in the membrane compartments after 30 min, but continues to increase in the exported polysaccharides. These workers were, therefore, able to calculate the rate of export of polysaccharide across the plasma membrane and to deduce that slime components would need to be displaced from the Golgi apparatus every 20 s, whereas polysaccharides for cell wall formation would only be displaced every 150 s. Presumably, application of this method would allow measurement of the effects of putative agonists and antagonists on exocytosis of root cap slime. A similar approach, labelling secreted polysaccharide with 3Hfucose or 14C-arabinose, has been employed to study the factors regulating exocytosis from sycamore suspension cells (Morris and Northcote, 1977). This work showed that addition of various cations caused a very rapid increase in polysaccharide secretion, and it was speculated that this was an effect on vesicle fusion mediated by a rise in intracellular Ca 2+ . In pea stem segments, rate zonal centrifugation has been used to separate Golgi stacks and putative secretory vesicles and, in conjunction with pulse-chase labelling (with 3H-glucose), transit of polysaccharide from Golgi to secretory vesicles to cell wall has been followed (Robinson et al, 1976; Griffing and Ray, 1985). One conclusion is that nigericin may inhibit exocytosis by inhibiting zlpH-driven secretory vesicle swelling (Griffing and Ray, 1985). This technique seems to have some potential, for instance for study of the effects of auxin on the exocytotic step (Napier and Venis, 1995), but it has not been very widely employed. A variety of probes can give more qualitative information on polysaccharide secretion. The fucose-specific lectin UeA I (Ulex europaeus agglutinin I; Rougier et al, 1979; Roy and Vian, 1991) shows the polar nature of fucoserich slime secretion from root cap cells. Calcofluor White is useful for detection of /3-glucans (Galbraith, 1981; Griffing et al, 1986). In the special case of cell plate formation, fusion of exocytotic vesicles leads to the formation of the new cross-wall at the end of telophase (see Battey and Blackbourn, 1993, for review). Aniline blue staining shows that /91-3 glucan (callose) is an early product of the newly formed cell plate, and that callose synthesis continues in cell plates after their isolation from BY2 cell protoplasts (Kakimoto and Shibaoka, 1992). This provides an excellent means of identifying isolated cell plates: aniline blue is reasonably specific for callose (Wood and Fulcher, 1984), and this is reflected in specific staining of the cell plate (Plate 2). Proteins Plate 1. Exocytosis from the maize root cap. Polysaccharide 'slime' is the visible product of vigorous exocytosis from the hypersecretory cells of the maize root cap. Hypersecretory activity is confined to only a few cell layers, is polarized, and is very sensitive to external factors (x 35). (Photograph: courtesy of Dave Maw, Jealott's Hill Research Station, Bracknell.) Secretion of a-amylase is perhaps the best characterized protein secretion in plants (see Jones and Jacobsen, 1991, for review). As a measure of exocytosis it is very useful because secretion can be measured from protoplasts (Bush et al, 1986). It occurs over a well-defined period (36-72 h), and is Ca2+-dependent. Further, secretion of a-amylase from protoplasts is controlled by both GA and ABA, which seem to exert their effects by changing intracellular Measurement of exocytosis 719 Plate 2. Cell plates can be detected by aniline blue staining. The procedure is based on that of Kakimoto and Shibaoka (1992); the cells are pretreated for 1 h with 3 mM UDP-glucose (a callose precursor) and then stained with aniline blue. Left: Nomarski optics. Right: UV fluorescence image of the same field. Upper panel: intact tobacco BY-2 cells showing synchronized cell plate formation after treatment with aphidicolin and propyzamide using a method based on that of Yasuhara et at. (1993). A minor component in commercial preparations of aniline blue shows yellow-green fluorescence when bound to callose in the cell plate. An example of a cell plate is arrowed. Lower panel: Cell plates isolated from tobacco BY-2 cell protoplasts according to the method of Kakimoto and Shibaoka (1992), DNase and cold treated, and further purified by sucrose density gradient centrifugation. Aniline blue fluorescence indicates that the cell plates are typically intact after this procedure, but the Nomarski image suggests that some cytoplasm still remains associated with some of the cell plates (x 145). 720 Battey et al. Ca2+-levels (Gilroy and Jones, 1992). Zorec and Tester (1992, 1993) have measured exocytosis as a change in membrane capacitance in aleurone protoplasts (see below). A parallel study of a-amylase secretion and exocytosis would be very informative, particularly in view of the predicted effects of GA and ABA on (Ca 2+ dependent) exocytosis in this system. There are other cell types where further study of protein secretion would be rewarding. Peroxidase secretion from spinach cell suspensions is stimulated rapidly by the addition of 1 mM Ca 2+ to the medium (Sticher et al, 1981). Although the possibility of release of cell wall peroxidases could not be ruled out, the evidence was in favour of an effect on release from the intracellular compartment and, hence, presumably exocytosis. It would be very interesting to study this process in protoplasts. plasma membrane must take place in hypersecretory root cap cells, to allow the vesicle fusion required for delivery of polysaccharide slime in the absence of cell growth (for discussion see Morre and Mollenhauer, 1983). One estimate puts this at 10% of the plasma membrane recycled per minute (Shannon and Steer, 1984). Even in the rapidly elongating cells of the coleoptile more than sufficient membrane can be delivered by secretory vesicles (Quaite et al, 1983; Phillips et al, 1988). Clearly estimates of exocytosis that depend on measurement of plasma membrane area (such as membrane capacitance-see below) must take account of this active recycling. This presents some difficulty because the mechanism of recovery is still unclear. In the root cap, coated vesicles appear to retrieve membrane not only from the plasma membrane, but also from secretory vesicles in transit (Mollenhauer et al, 1991). However, it is not clear that endocytosis is quantitatively sufficient to account for the required rate of membrane recovery Measurement of plasma membrane area (Shannon and Steer, 1984; Emons and Traas, 1986; Steer, The maximum elastic stretching of the plasma membrane 1988). The close association of cortical ER with the is about 2% of its surface area (Wolfe and Steponkus, plasma membrane may facilitate molecular recycling of 1981, 1983; Wolfe et al., 1985; Dowgerte/ al., 1987). The membrane lipids (Craig and Staehelin, 1988), a mechanauthors concluded that further increases in plasma memism that has yet to be discounted and may be favoured brane area can only be accounted for by the addition of by the high turgor pressure acting on the plant plasma new membrane material from intracellular stores. The membrane (Cram, 1980; Staehelin and Chapman, 1987). problem is, therefore, to estimate accurately the rate of Grabski et al. (1993) obtained evidence for rapid transfer this addition. Micropipette aspiration techniques, in of fluorescently-labelled lipids from the plasma membrane which negative pressures are applied to patches of memto the ER, and have interpreted this as consistent with brane through a micropipette, reveal an equilibrium mem1 such a recycling mechanism. It is also possible that brane tension (mean value 120 /xN m" ) above which exocytosis leads to formation of a pore, release of vesicle new material is added to the plasma membrane until the contents, and then vesicle reformation, so that vesicle tension is released (Wolfe et al., 1985). Such exocytotic membrane is only temporarily incorporated into the addition of membrane can accommodate surface area plasma membrane. This has been proposed to explain changes resulting in tensions of up to 4 mN m" 1 ; above the current 'flicker' detected in nerve terminals (Spruce this protoplast lysis occurs. This approach illustrates the et al, 1990). exocytotic delivery of new membrane. However, it does The most complete descriptive model for the exocytotic not allow quantitative determination of the kinetics of event in plant cells has been presented by Staehelin, based membrane flux. on observation of disc- and horseshoe-shaped infoldings of the plasma membrane in suspension culture (Staehelin Estimation of vesicle supply and recycling using microscopy and Chapman, 1987) and root cap cells (Craig and This topic has been comprehensively reviewed by Steer Staehelin, 1988) (Plate 3). These are interpreted as steps (1988). Specific aspects of immediate relevance to the in the fusion of exocytotic vesicles, which after fusion are measurement of exocytosis are focused on here. The most flattened due to the high turgor pressure of plant cells, detailed transmission electron microscope (TEM) anaand then tip over sideways, presumably as a result of lysis is of pollen tube growth in relation to secretory movement of the cytoplasm relative to the plasma memvesicle formation/delivery rates (Picton and Steer, 1983; brane (Fig. 1). This model has been confirmed in work Steer and Picton, 1984). The principal conclusion is that on carrot suspension cells (Emons et al, 1992). A decline vesicle production rates are very similar in fast- and slowin the number of exocytotic configurations in elongated growing tubes, suggesting that vesicles are produced in cells between 1 and 14 d after subculturing may reflect a excess to allow maximum growth under favourable condidecrease in transport of hemicelluloses and pectins due to tions (Steer and Picton, 1984). This means that an excess exhaustion of sugar in the medium (Emons et al, 1992). of membrane is delivered to the pollen tube tip, so that It is interesting that these important observations of some form of recycling must normally occur. Similar Staehelin and Emons are not necessarily inconsistent with considerations indicate that very active removal of the the idea of transient vesicle fusion discussed by Spruce Measurement of exocytosis 721 Plate 3. Exocytotic configurations in the plasma membrane of a sycamore suspension cell. Freeze-fracture electron microscopy was used on the plasma membrane of cells ultra-rapidly frozen by liquid propane. This view of the exoplasmic face of the membrane shows horseshoe-shaped membrane infoldings that are believed to reflect exocytotic vesicle fusion. For the mechanism proposed to account for these shapes, see Fig. 1. Reproduced from Staehelin and Chapman (1987) by permission of Springer-Verlag, and courtesy of the authors (x40 000). Fig. 1. The Staehelin/Chapman model for exocytosis from plant cells. The diagrams illustrate the stages in vesicle fusion that Staehelin and Chapman (1987) suggest lead to the horseshoe-shaped configurations shown in Plate 3 Top row diagrams: plasma membrane viewed from the inside (plasmatic face, PF) as the vesicle approaches (I), and docks and forms a fusion pore (II) At stage III the fusion pore is deformed to a slit because the vesicle is flattened under the forces of turgor following content release, and at stage IV the vesicle topples over and the fusion slit expands along the margins of the vesicle to give the horseshoe-shape. Bottom row diagrams: side view of the same series of events. Stages III and IV appear to be unique to plants and probably reflect the effects of high turgor. The model implies that the plasma membrane can not immediately expand to accommodate the new vesicle membrane after fusion, presumably because it is constrained by the cell wall. Redrawn from Staehelin and Chapman (1987). et al. (1990): it is not clear what happens after flattening of the vesicle (Stage IV in Fig. 1). Recovery of the spent vesicle could be the next step (Craig and Staehelin, 1988; Staehelin and Chapman, 1987). However, TEM pictures of hypersecretory root cap cells certainly give the impression of complete vesicle incorporation into the plasma membrane (Plate 4), so it is possible that both mechanisms operate. 722 Battey et al. Plate 4. Transmission electron micrograph of a hypersecretory cell from the maize root cap. The section was double stained with uranyl acetate/lead citrate according to standard procedures. The polysacchande slime is unstained and is massed along the upper edge of the cell. The profile of a recently fused secretory vesicle is visible (large arrow), and vesicles apparently about to fuse can also be seen (small arrows). Note the cytoplasm contains both stained and unstained vesicles (x9200). Estimation of plasma membrane area by electrophysiological measurements of membrane capacitance: theory Biological membranes function as a barrier between two electrolyte solutions, and so they have a number of electrical properties. One of these is capacitance (Cm), which in all membranes has a constant value per unit surface area xl ^Fcm~ 2 . The current flowing through a capacitor is proportional to the rate of change of the applied voltage, i.e. T=CxdV/dt where C=capacitance. Exocytosis involves the addition of new membrane, which if added at a greater rate than it is removed will result in an increase in membrane surface area. Thus measurement of the changes in Cm provides a valuable, dynamic and geometry independent probe in the study of exocytosis. The measurement of capacitance in plant cell membranes depends upon the application of a changing voltage across the membrane and detection of the currents induced. This can be done using the whole cell patch clamp technique (Hammill et al., 1981). To measure Cm from whole cell recordings, a sinusoidal command voltage (Kcom) is applied to a cell inducing a sinusoidal current to flow across the plasma membrane. This current has both resistive and capacitative components, each with a distinctive phase angle (0) relative to the command signal. The phase of the capacitative current is orthogonal to the phase of the resistive current under normal circumstances. Thus, determining the phase of the resistive current (0 — 90°) allows the phase angle of the capacitative current (d) to be calculated; the current in this phase is measured as an output signal proportional to the membrane capacitance, but independent of membrane resistance (Fig. 2). Techniques used to monitor the changes in membrane capacitance following exocytotic events were developed principally to study stimulus-secretion coupling in excitable animal cells and have been used subsequently in a wide range of animal cells (Neher and Marty, 1982, Fernandez et al., 1984; Mason et al, 1989; Schweizer et al, 1989; Zorec et al., 1991). Two general approaches have been followed in the measurement of cell membrane capacitance, identical in theory but different in methodology. These are the phase detection technique using a lock-in amplifier (Neher and Marty, 1982), and the phasetracking technique (Fidler and Fernandez, 1989). The principles involved are illustrated in Fig. 2A and B. There are inherent problems with the phase detection technique, because the impedance of the cell may change during the experiment, as plasma membrane ion channels open and close; or the series resistance (Rs—see Fig. 2) may increase as the membrane begins to reform over the ruptured patch between the walls of the pipette. This causes the current from the cell (7C) to drift out of phase, so that the amplifier is not necessarily reading current at a phase directly proportional to membrane capacitance (Fig. 2C). In response to these problems, Fidler and Fernandez (1989) described a phase detection technique whereby the phase angle (9) is automatically tracked and continuously adjusted. This means that changes in the electrical properties of the cell during the course of the experiment can be accommodated. This approach is currently being used in studies of exocytosis from root cap protoplasts. The electrophysiological measurement of exocytosis in plant cells offers a number of opportunities. It allows exocytosis and membrane retrieval to be monitored directly; further, because the whole cell patch clamp configuration results in direct communication between the cytosol and the microelectrode filling solution, cytosolic Ca2+ can be buffered and putative regulators of exocytosis perfused into the cytosol and their effects recorded. There is also the potential to distinguish effects on exocytosis from those on membrane retrieval. In animals this has been achieved where there is clear temporal separation of the two processes (e.g. in neurons; von Gersdorff and Matthews, 1994; Heidelberger et al., 1994), or where secretory vesicles are sufficiently large to allow individual fusion events to be monitored (e.g. 0.5-1 /nm in mast cells; Breckenridge and Aimers, 1987). In plants, even where secretory vesicles are relatively large (e.g. about 500 nm in root cap hypersecretory cells; Plate 4) , this size is at the lower end of the detectable range; and, in Measurement of exocytosis general, exocytosis- might be expected to be accompanied by continuous membrane retrieval (although more information is needed here). These are important considerations because they imply that as it stands the patch clamp method will be useful for measuring the net result of exocytosis and membrane retrieval, but not for distinguishing between the two processes. Estimation of plasma membrane area by membrane capacitance: practical expectations and observations Considerations of membrane recycling (see above) suggest that secretory vesicle fusion should be followed by membrane recovery. In protoplasts, unless swelling is occurring, Cm might, therefore, be expected to increase following vesicle fusion and then return to its original value. However, what is actually observed will depend critically on the number of vesicles arriving at the plasma membrane per second, vesicle size in relation to plasma membrane area, and the actual rate of membrane recovery in the specific cell type. Typically, secretory vesicles are about 150nm in diameter (Picton and Steer, 1981) and a protoplast about 30 /nm in diameter. If each is assumed to be spherical then a vesicle will contribute only 0.0025% of the plasma membrane area. Capacitance changes of 1 pF can be detected readily (equivalent to about 3000 secretory vesicle fusions, but an increase of only 2% in plasma membrane area), and so quite a large amount of vesicle fusion can occur without a visible increase in protoplast volume. On the other hand, in hypersecretory root cap cells it has been estimated that an area equivalent to the whole plasma membrane is delivered every 10 min (Shannon and Steer, 1984); this gives an idea of the rates of membrane delivery that are possible. However, even in this case, if membrane recovery occurs as an immediate consequence of vesicle fusion, capacitance will increase only very transiently. It should also be noted that capacitance would be expected to increase if the membrane thins significantly due to elastic stretching. This exocytosis-independent increase in capacitance is generally thought to be small, but this is an assumption that remains to be verified. Cm has now been measured in protoplasts from the barley aleurone layer (Zorec and Tester, 1992, 1993), and maize coleoptile (Thiel et al., 1994). The results are consistent in showing an increase in Cm in response to approximately 1 yM free Ca 2 + in the patch-pipette, and a decrease in response to < 3 0 n M free Ca 2 + . This suggests that elevated Ca 2 + causes an increase in the rate of exocytosis (equivalent to about 10 vesicles s" 1 in the aleurone protoplast; Zorec and Tester, 1992), relative to the rate of membrane recovery. Further, the effect of reduced Ca 2 + concentrations suggests that even basal exocytosis (that needed to compensate for membrane recovery) is Ca 2+ -dependent. Application of pressure 723 through the patch-pipette causes protoplast swelling, followed by a (5-fold slower) contraction to the original diameter (Zorec and Tester, 1993). This is interesting, because it suggests that exocytosis can be triggered (or membrane recovery inhibited) directly by pressure, and that plant cells have greatly enhanced sensitivity to turgor pressure compared with animal cells. It is also significant that pressure appeared to stimulate fusion of a different population of vesicles than that involved in the Ca 2 + dependent response (Zorec and Tester, 1993). The response of protoplasts to pressure is likely to be different from the response of walled cells. It remains an intriguing possibility that membrane recovery could be activated under circumstances in which the cell wall forbids further expansion in response to exocytosis. This is in a sense the corollary of the proposal that tension generated in the plane of the plasma membrane by cell wall yielding provides the driving force for increase in plasma membrane surface area (Kell and Glaser, 1993). Accurate measurements of membrane capacitance, turgor, and wall extension would indicate how the regulatory hierarchy between these processes is maintained, and might, in time, make the mechanism of action of regulators such as auxin more intelligible. Cell-free assays for exocytosis Two cell-free approaches have been used previously to assay for exocytosis in plants. Baydoun and Northcote (1980) isolated Golgi-rich and plasma membrane-rich fractions by density gradient centrifugation of 14C-choline labelled and unlabelled root tips of maize. After mixing, the fractions were re-purified and the transfer of 14C-lipid was measured. Evidence was obtained for fusion between the Golgi and plasma membrane fractions; Ca 2 + and a membrane protein of 36 kDa were demonstrated to be key factors for the fusion process (Baydoun and Northcote, 1981). This pioneering work depended solely on marker enzymes for identification of Golgi and plasma membranes, and there was no indication of the intactness or otherwise of the secretory vesicles and the Golgi. Nevertheless, the results provided the first indication that cell-free assays for exocytosis were possible in plants, and that Ca 2 + might have a significant role. Morre et al. (1991) immobilized spinach plasma membrane on nitrocellulose filters, and then measured the transfer of 14 Clipid from a Golgi fraction to the plasma membrane acceptor. Transfer was temperature-sensitive and there was a suggestion that it might be stimulated by ATP. However, it is not clear that aggregation would be distinguished from fusion using this approach. These assays point the way for future work, and highlight the following areas for improvement. 724 Battey et al. A. B R R I Computer control c. v commV I c pA Ill IcpA Fig. 2. Schematic diagrams of a patch-clampcd cell illustrating the two methods of determining the phase angle d. In both systems the patch clamp amplifier measures the current flowing in response to a sinusoidally varying command voltage ( l ^ ) and the equivalent circuit model of a patchclamped cell consists of series resistance R,, the cell membrane resistance Rm and the membrane capacitance Cm (A). The lock-in or stationary phase detection technique, in which current from the cell / c is balanced by the current from the amplifier's transient cancellation circuitary / a at a summing junction ( + ) . R^ and Ca are, respectively, variable series resistance and whole cell capacitance settings. R, and C2 are fixed resistance and capacitance elements in parallel with Ra and C a . After making adjustments of Ra and C, so that / a = / c , Ri or C 2 is switched in using switches 1 or 2, respectively, to create controlled changes in the amplifier current. By altering the phase angle detector the amplifier is able to measure / c at a phase where switching in /?[ or C 2 produces the largest current response; this phase angle is 9-90° if /?, is switched in, and 9 if C2 is switched in. The proportionality of the induced current to the membrane capacitance is strongly dependent upon the accuracy with which the phase angle is determined. Increases in cell capacitance are calibrated by making incremental increases of the whole cell capacitance compensation on the amplifier (simulating increases in membrane capacitance) and noting the current response, in mammalian cells the phase detection technique has been used to monitor accurately capacitance changes in the range 0.4-80 fF, sufficient to observe the fusion of a single secretory granule. (B). The phase- Measurement of exocytosis Purification of intact secretory vesicles, and distinguishing these from vesiculated Golgi A marker is needed that distinguishes post-Golgi vesicles from Golgi. The absence of the Golgi enzyme xyloglucan xylosyl transferase from secretory vesicles, while the fucosyl transferase remains active, may provide one such marker (Brummel et al, 1990). The polysaccharide content of secretory vesicles changes as they mature, at least in hypersecretory root cap cells (Battey and Blackbourn, 1993). This might provide a marker for sectioned membrane pellets under the TEM. Purification of cytoplasmic side-out plasma membrane vesicles Aqueous two-phase partitioning produces a population of plasma membrane vesicles which are mainly 'outsideout' in orientation (Larsson et al, 1984). The vesicles can be turned into the required cytoplasmic side-out orientation by snap freezing and thawing (Brightman and Morre, 1992; Larsson et al, 1994). Separation of vesicles remaining in the outside-out orientation can then be achieved using the countercurrent distribution procedure (Widell et al, 1982; Akerlund and Albertsson, 1994). An alternative, and potentially much more rapid method involves incubating phase-partitioned plasma membrane vesicles with the detergent Brij 58, which turns the vesicles inside-out (Johansson et al, 1995). The only potential problem with this very convenient method is if the detergent alters the fusigenic properties of the plasma membrane vesicles. Distinguishing secretory vesicle docking (membrane aggregation) from fusion with the plasma membrane The assays developed by Baydoun and Northcote (1980, 1981) and Morre et al (1991) do not necessarily distinguish between docking and fusion, although that described by Baydoun and Northcote would eliminate Ca 2+ -dependent docking. Assays that measure membrane or content mixing should reflect only fusion, although even these methods need to be very carefully controlled. It is reasonably well established that vesicle fusion is accompanied by mixing of the lipids from the two membranes. This can be measured using lipid-soluble fluorescent probes that can be added to native membrane vesicles after purification. In the first approach one population of vesicles is saturated with a probe such as octadecylrhodamine B chloride ( R i 8 ) . On mixing with 725 the target membrane (unlabelled), the probe is diluted and fluorescence dequenching occurs; this can be quantified and used as a measure of fusion (Hoekstra et al, 1984; Mac Lean and Edwardson, 1992; Hoekstra and Klappe, 1993; Lee et al, 1994). This type of assay has been the subject of some criticism and debate, still currently unresolved (Stegman et al, 1995; Edwardson, 1995). In the second approach, resonance energy transfer between two lipid-soluble fluorophores, with the emission spectrum of one fluorophore partly overlapping the excitation spectrum of the other, is used to monitor membrane mixing (Comerford and Dawson, 1988; Keller et al, 1977). Energy transfer depends on close proximity between the lipid probes. Mixing a population of donor vesicles with both probes loaded in their membranes, with unlabelled acceptor vesicles, will therefore result in a decrease in resonance energy transfer efficiency. Alternatively, each probe can be loaded into a different membrane; fusion is then indicated by an increase in transfer efficiency. The advantages of energy transfer assays for monitoring fusion have been described (Hoekstra and Duzgiines, 1993), and include high sensitivity at low probe labelling rates, continuous monitoring, and wide applicability involving both artificial liposomes and native membranes. The relevance of this assay for measuring fusion in biological membranes is indicated by the good agreement between measurements of cell fusion (induced by high PEG concentrations) made by syncitia counting and by the resonance energy transfer assay (Partearroyo et al., 1994). A particularly attractive feature of the energy transfer assay is its potential to make visible the process of membrane fusion (Uster, 1993). The resonance energy transfer assay depends on probe mixing and factors that affect diffusion of lipids in the membrane bilayer are, therefore, likely to affect the assay. Further, because native membranes vary in composition, more variation in labelling efficiency can be expected than with artificial liposomes. However, assays can be standardized by diluting labelled donor or target membranes with unlabelled membranes to a set ratio of fluorescence intensities, or, if the fluorophores are in separate membranes, to a set ratio of membrane amounts. As with other assays, resonance energy transfer experiments should include proper positive (e.g. PEG-induced fusion) and negative controls (e.g. the use of EGTA to exclude Ca 2 + -induced aggregation) in order to assess real fusion events. Figure 3 shows an emission spectrum from a tracking detection technique. Here a 1 Mfi resistor is in series with the cell. In parallel to this resistor is a computer-controlled reed relay switch. During the course of an experiment this series resistance is switched in and the phase detector finds the phase of lc at which a maximal change occurs, this phase angle being 8. (C). Illustration of the change in phase of lc with respect to the phase of l^,,,,, when either /? t (lock-in method) or the 1 MQ resistor (phase-tracking method) is switched in. Traces (i) and (ii) show the phase of Vcom and /„ respectively; temporarily increasing a resistive element (trace iii) causes Ic to shift relative to the phase angle (8-90°) of the lock-in amplifier (broken line). This causes an error in the /,. reading and requires correction using the phase detector. 726 Battey et al. 125 o u a v u o 500 S2S 550 S7S 600 Emission wavelength (nm) Fig. 3. Resonance energy transfer assay for vesicle fusion Emission spectrum of fluorescein-labelled plasma membrane vesicles mixed with rhodamine-labelled Golgi membrane vesicles, after irradiation at the excitation wavelength of fluorescein (460 nm). The increased emission of rhodamine at 592 nm indicates energy transfer from fluorescein to rhodamine and thus fusion between the differently labelled membranes. Polyethylene glycol (PEG 3350) promotes vesicle fusion by removing water from the vicinity of the membranes. Triton X-100 dissolves the membranes and is a negative control. resonance energy transfer assay of fusion between maize Golgi-enriched vesicles and plasma membrane vesicles. Content mixing assays provide an ideal method of measuring vesicle fusion. However, even in this case problems can arise due to leakage of contents during the assay (Hoekstra and Diizgunes, 1993). Some content mixing methods depend on incorporation of the probes during the preparation of artificial liposomes used in the assay; this is the case with the terbium/dipicolinic acid technique developed by Diizgunes and Wilschut (1993). These assays are therefore not so useful for studies of exocytosis, in which native vesicles are essential. Conclusions In vivo quantification of exocytosis can be achieved by capacitance measurements, and this approach has the great advantage that potential regulators of exocytosis can be tested after dialysis through the patch pipette. However, the current limitation of patch-clamping for plant cells is that it is necessary to use protoplasts. The intact cell wall certainly has regulatory influences on plasma membrane expansion and hence on exocytosis and membrane recovery; it is the interplay between these biophysical effects and their translation into the biochemical domain that is really crucial to the understanding of exocytosis in plant cells. The potential importance of turgor pressure for exocytosis is illustrated by the apparently unique character of the fusion process in plants (Staehelin and Chapman, 1987; Emons et al, 1992; see above). These observations emphasize the difficulties con- fronting the attempt to quantify exocytosis, particularly in a cell-free context. Nevertheless, a cell-free approach is probably essential, to provide biochemical evidence of the nature and function of regulatory molecules. This needs to be complemented by further work towards the cloning and characterization of homologues of known regulators such as syntaxin (Bassham et al, 1995). The identification of exocytotic mutants in Arabidopsis, the use of epitopetagged secretory products, and advances in our knowledge of polysaccharide and protein processing en route to the plasma membrane will provide major opportunities for improvements in the currently available techniques for measuring exocytosis in plant cellls. Acknowledgements We are grateful to the BBSRC, Leverhulme Trust, Royal Society, and The University of Reading Research Endowment Trust Fund for financial support. Thanks to Paul Le Miere for help with Figure preparation, and to Richard Napier (HRI, Wellesbourne) and two anonymous referees for their invaluable comments on the manuscript. References Akerlund H-E, Albertsson P-E. 1994. Thin-layer counter current distribution and centrifugal counter current distribution apparatus. Methods in Enzymology 228, 87-99. Battey NH, Blackbourn HD. 1993. The control of exocytosis in plant cells. Tansley Review No. 57. New Phvtologist 125, 307-38. Bassham DC, Gal S, Da Silva Conceicao A, Raikhel NV. 1995. An Arabidopsis syntaxin homologue isolated by functional complementation of a yeast pepl2 mutant. Proceedings of the National Academy of Sciences USA 92, 7262-6. Baydoun EA-H, Northcote DH. 1980. Measurement and characteristics of fusion of isolated membrane fractions from maize root tips. Journal of Cell Science 45, 169-86. Baydoun EA-H, Northcote DH. 1981. The extraction from maize (Zea mays) root cells of membrane-bound protein with Ca 2+ -dependent ATPase activity and its possible role in membrane fusion in vitro. Biochemical Journal 193, 781-92. Bowles DJ, Northcote DH. 1974. The amounts and rates of export of polysaccharides found within the membrane system of maize root cells. Biochemical Journal 142, 139-44. Breckenridge LJ, Aimers W. 1987. Final steps in exocytosis observed in a cell with giant secretory granules. Proceedings of the National Academy of Sciences USA 84, 1945-9. Brightman AO, Morre DJ. 1992. Sidedness of plant plasma membrane vesicles altered by conditions of preparation. Plant Physiology 98, 183-90. Brummell DA, Camlrand A, MacLachlan GA. 1990. Differential distribution of xyloglucan glycosyl transferases in pea Golgi dictyosomes and secretory vesicles. Journal of Cell Science 96, 705-10. Bush DS, Cornejo M-J, Huang C-N, Jones RL. 1986. Ca 2 + stimulated secretion of a-amylase during development in barley aleurone protoplasts. Plant Physiology 82, 566-74. Comerford JG, Dawson AP. 1988. The mechanism of action of Measurement of exocytosis 2+ GTP on Ca efflux from rat liver microsomal vesicles. Biochemical Journal 249, 89-93. Craig S, Staehelin LA. 1988. High pressure freezing of intact plant tissues. Evaluation and characterization of novel features of the endoplasmic reticulum and associated membrane systems. European Journal of Cell Biology 46, 80-93. Cram WJ. 1980. Pinocytosis in plants. New Phytologist 84, 1-17. Dowgert MF, Wolfe J, Steponkus PL. 1987. The mechanics of injury to isolated protoplasts following osmotic contraction and expansion. Plant Physiology 83, 1001-7. Duzgiines N, Wilschut J. 1993. Fusion assays monitoring intermixing of aqueous contents. Methods in Enzymology 220, 3-14. Edwardson JH. 1995. Measurement of fusion between pancreatic zymogen granules and plasma membrane in vitro. Biochemical Journal 307, 876-8. Emons A-MC, Traas JA. 1986. Coated pits and coated vesicles on the plasma membrane of plant cells. European Journal of Cell Biology 41, 57-64. Emons A-MC, Vos JW, Kieft H. 1992. A freeze-fracture analysis of the surface of embryogenic and non-embryogenic suspension cells of Daucus carota. Plant Science 87, 85-97. Fernandez JM, Neher E, Gomperts BD. 1984. Capacitance measurements reveal stepwise fusion events in degranulating mast cells. Nature 312, 453-55. Fidler N, Fernandez JM. 1989. Phase tracking: an improved phase detection technique for cell membrane capacitance measurements. Biophysical Journal 56, 1153-62. Galbraith DW. 1981. Microfluorimetric quantitation of cellulose biosynthesis by plant protoplasts using Calcofluor White. Physiologia Plantarum 53, 111-16. Gilroy S, Jones RL. 1992. Gibberellic acid and abscisic acid co-ordinately regulate cytoplasmic calcium and secretory activity in barley aleurone protoplasts. Proceedings of the National Academy of Sciences USA 89, 3591-5. Grabski S, de Feijter AW, Schindler M. 1993. Endoplasmic reticulum forms a dynamic continuum for lipid diffusion between contiguous soybean root cells. The Plant Cell 5, 25-38. Grifflng LR, Ray PM. 1985. Involvement of monovalent cations in Golgi secretion by plant cells. European Journal of Cell Biology 36, 24-31. Grifflng LR, Mersey BG, Fowke LC. 1986. Cell-fractionation analysis of glucan synthase I and II distribution and polysaccharide secretion in soybean protoplasts. Planta 167, 175-12. Hammill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. 1985. Improved patch-clamp techniques for high resolution current recording from cells and cell-free membrane patches. Pfluegers Archiv. European Journal of Physiology 391, 85-100. Heidelberger R, Heinemann C, Neher E, Matthews G. 1994. Calcium dependence of the rate of exocytosis in a synaptic terminal. Nature 371, 513-15. Hoekstra D, Diizgunes N. 1993. Lipid mixing assays to determine fusion in liposome systems. Methods in Enzymology 220, 15-32. Hoekstra D, Klappe K. 1993. Fluorescence assays to monitor fusion of enveloped viruses. Methods in Enzymology 220, 261-76. Hoekstra D, de Boer T, Klappe K, Wilschut J. 1984. Fluorescence method for measuring the kinetics of fusion between biological membranes. Biochemistry 23, 5675-81. Johansson F, Olbe M, Sommarin M, Larsson C. 1995. Brij 58, a polyoxyethylene acyl ether, creates membrane vesicles of uniform sidedness. A new tool to obtain inside-out (cyto- 727 plasmic side-out) plasma membrane vesicles. The Plant Journal!, 165-73. Jones RL, Jacobsen JV. 1991. Regulation of synthesis and transport of secreted proteins in cereal aleurone. International Review of Cytology 126, 49-88. Kakimoto T, Shibaoka H. 1992. Synthesis of polysaccharides in phragmoplasts isolated from tobacco BY-2 cells. Plant Cell Physiology 33, 353-61. Kell A, Glaser RW. 1993. On the mechanical and dynamic properties of plant cell membranes: their role in growth, direct gene transfer and protoplast fusion. Journal of Theoretical Biology 160, 41-62. Keller PM, Person S, Snipes W. 1977. A fluorescence enhancement assay of cell fusion. Journal of Cell Science 28, 167-77. Larsson C, KjeUborn P, Widell S, Lundberg T. 1984. Sidedness of plant plasma membrane vesicles purified by partitioning in aqueous two-phase systems. FEBS Letters 171, 271-6. Larsson C, Sommarin M, Widell S. 1994. Isolation of highly purified plant plasma membranes and separation of insideout and right-side-out vesicles. Methods in Enzymology 228, 451-69. Lee EG, Marciniak SJ, Maclean CM, Edwardson JM. 1994. Pancreatic plasma membranes: promiscuous partners in membrane fusion. Biochemical Journal 298, 599-604. MacLean CM, Edwardson JM. 1992. Fusion between rat pancreatic zymogen granules and plasma membranes: modulation by a GTP-binding protein. Biochemical Journal 286, 747-53. Mason WT, Sikdar SK, Zorec R, Akerman S, Rawlings SR, Cheek T, Moreton R, Berridge M. 1989. Ion channels, intracellular calcium and exocytosis: control of hormone secretion in cultured bovine pituitary lactotrophs. In: Oxford GS, Armstrong CM, eds. Secretion and its control. New York: Rockefeller University Press, 225-38. Mollenhauer HH, Morre DJ, Griffing LR. 1991. Post-Golgi apparatus structures and membrane removal in plants. Protoplasma 162, 55-60. Morre DJ, Mollenhauer HH. 1983. Dictyosome polarity and membrane differentiation in outer cap cells of the maize root tip. European Journal of Cell Biology 29, 126-32. Morre DJ, Penel C, Morre DM, Sandelius A-S, Moreau P, Andersson B. 1991. Cell-free transfer and sorting of membrane lipids in spinach: donor and acceptor specificity. Protoplasma 160, 49-64. Morris MR, Northcote DH. 1977. Influence of cations at the plasma membrane in controlling polysaccharide secretion from sycamore suspension cells. Biochemical Journal 166, 603-18. Napier RM, Venis MA. 1995. Auxin action and auxin-binding proteins. Tansley Review No. 79. New Phytologist 129, 167-201. Neher E, Marty A. 1982. Discrete changes in cell membrane capacitance observed under conditions of enhanced secretion in bovine adrenal chromaffin cells. Proceedings of the National Academy of Sciences USA 79, 6712-16. Partearroyo MA, Cabezon E, Nieva J-L, Alonso A, Goiii FM. 1994. Real-time measurements of chemically-induced membrane fusion in cell monolayers, using a resonance energy transfer method. Biochimica el Biophysica Ada 1189, 175-80. Phillips GD, Preshaw C, Steer MW. 1988. Dictyosome vesicle production and plasma membrane turnover in auxinstimulated outer epidermal cells of coleoptile segments from Avena sativa (L.). Protoplasma 145, 59-65. Picton JM, Steer MW. 1981. Determination of secretory vesicle production rates by dictyosomes in pollen tubes of 728 Battey et al. Tradescantia using cytochalasin D. Journal of Cell Science 49, 261-72. Picton JM, Steer MW. 1983. Evidence for the role of Ca 2 + ions in tip extension in pollen tubes. Protoplasma 115, 11-17. Quaite E, Parker RE, Steer MW. 1983. Plant cell extension: structural implications for the origin of the plasma membrane. Plant, Cell and Environment 6, 429-32. Robinson DG, Eisinger WR, Ray PM. 1976. Dynamics of the Golgi system in wall matrix polysaccharide synthesis and secretion by pea cells. Berichte der Deutschen Botanischen Gesellschaft 89, 147-61. Rougier M. 1981. Secretory activity of the root cap. In: Tanner W, Loewus FA, eds. Encyclopaedia of plant physiology, Vol. 13B, Plant carbohydrates II. Extracellular carbohydrates. Berlin: Springer-Verlag, 542-74. Rougier M, Kieda C, Monsigny M. 1979. Use of lectin to detect the sugar components of maize root cap slime. Journal of Histochemistry and Cytochemistry 27, 878-81. Roy S, Vian B. 1991. Transmural exocytosis in maize root cap: visualization by simultaneous use of a cellulose-probe and a fucose-probe. Protoplasma 161, 181-91. Schweizer FE, Shafer T, Tapparelli C, Grob M, Karli UO, Heumann R, Thoenen H, Bookman RJ, Burger MM. 1989. Inhibition of exocytosis by intracellularly applied antibodies against a chromaffin granule-binding protein. Nature 339, 709-12. Shannon TM, Steer MW. 1984. The root cap as a test system for the evaluation of Golgi inhibitors. I. The structure and dynamics of the secretory system and response to solvents. Journal of Experimental Botany 35, 1697-1707. Spruce AE, Breckenridge LJ, Lee AK, Aimers W. 1990. Properties of the fusion pore that forms during exocytosis of a mast cell secretory vesicle. Neuron 4, 643-54. Staehelin LA, Chapman RL. 1987. Secretion and membrane recycling in plant cells: novel intermediary structures visualized in ultra-rapidly frozen sycamore and carrot suspensionculture cells. Planta 171, 43-57. Steer MW. 1988. Plasma membrane turnover in plant cells. Journal of Experimental Botany 39, 987-96. Steer MW, Picton JM. 1984. Control of cell wall formation in pollen tubes: the interaction of dictyosome activity with the rate of tip extension. In: Dugger WM, Bartnicki-Garcia S, eds. Structure, function and biosynthesis of plant cell walls. Maryland, USA: American Society of Plant Physiologists, 483-94. Stegman T, Orsel JG, Jamieson JD, Padfield PJ. 1995. Limitations of the octadecylrhodamine dequenching assay for membrane fusion. Biochemical Journal 307, 875-8. Sticher L, Penel C, Greppin H. 1981. Calcium requirement for the secretion of peroxidases by plant cell suspensions. Journal of Cell Science 48, 345-53. Thiel G, Ruprik M, Zorec R. 1994. Raising the cytosolic Ca2 + concentration increases the membrane capacitance of maize coleoptile protoplasts: evidence for Ca 2+ -stimulated exocytosis. Planta 195, 305-8. Uster PS. 1993. In situ resonance energy transfer microscopy: monitoring membrane fusion in living cells. Methods in Enzymology 221, 239-46. Von Gersdorff H, Matthews G. 1994. Inhibition of endocytosis by elevated internal calcium in a synaptic terminal. Nature 370, 652-5. Widell S, Lundborg T, Larsson C. 1982. Plasma membranes from oats prepared by partition in an aqueous polymer twophase system. Plant Physiology 70, 1429-35. Wolfe J, Dowgert MF, Steponkus PL. 1985. Dynamics of membrane exchange of the plasma membrane and the lysis of isolated protoplasts during rapid expansion in area. Journal of Membrane Biology 86, 127-38. Wolfe J, Steponkus PL. 1981. The stress-strain relation of the plasma membrane of isolated plant protoplasts. Biochimica el Biophysica Ada 643, 663-8. Wolfe J, Steponkus PL. 1983. Mechanical properties of the plasma membrane of isolated plant protoplasts. Plant Physiology 71, 276-85. Wood PJ, Fulcher RG. 1984. Specific interaction of aniline blue with (1 ->3)-/3-D-glucan. Carbohydrate Polymers 4, 49-72. Yasuhara H, Sonobe S, Shibaoka H. 1993. Effects of taxol on the development of the cell plate and of the phragmoplast in tobacco BY-2 cells. Plant and Cell Physiology 34, 21-9. Zorec R, Sikdar SK, Mason WT. 1991. Increased cytosolic calcium stimulates exocytosis in bovine lactotrophs. Direct evidence from changes in membrane capacitance. Journal of General Physiology 97, 473-97. Zorec R, Tester M. 1992. Cytoplasmic calcium stimulates exocytosis in a plant secretory cell. Biophysical Journal 63, 864-7. Zorec R, Tester M. 1993. Rapid pressure driven exocytosisendocytosis cycle in a single plant cell: capacitance measurements in aleurone protoplasts. FEBS Letters 333, 283-6.