Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
List of types of proteins wikipedia , lookup
Histone acetylation and deacetylation wikipedia , lookup
Community fingerprinting wikipedia , lookup
Transcription factor wikipedia , lookup
Deoxyribozyme wikipedia , lookup
Alternative splicing wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Polyadenylation wikipedia , lookup
Messenger RNA wikipedia , lookup
RNA interference wikipedia , lookup
Genomic imprinting wikipedia , lookup
RNA silencing wikipedia , lookup
Non-coding DNA wikipedia , lookup
Genome evolution wikipedia , lookup
Molecular evolution wikipedia , lookup
Ridge (biology) wikipedia , lookup
Gene regulatory network wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Gene expression profiling wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Eukaryotic transcription wikipedia , lookup
Promoter (genetics) wikipedia , lookup
RNA polymerase II holoenzyme wikipedia , lookup
Non-coding RNA wikipedia , lookup
Epitranscriptome wikipedia , lookup
Gene expression wikipedia , lookup
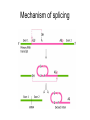
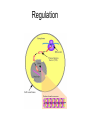
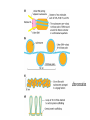
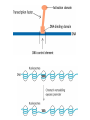
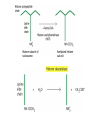
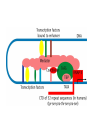
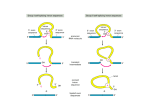
Transcription in eucaryotes The basic chemistry of RNA synthesis in eukaryotes is the same as in prokaryotes. Genes coding for proteins are coded for by RNA polymerase II, which is a complex enzyme. Capping the RNA transcribed by RNA polymerase II • • • During the initial synthesis of an mRNA the first NTP retains its triphosphate group. Capping consists of removing the phosphate and adding a GMP 5´-5´ (Fig. 24.11). The G which is added is then methylated at N-7 and the ribose of the original G is methylated on the 2´-OH. A further methyl may be added to the next ribose. Capping protects mRNA from exonucleases and is involved in translation initiation. Split genes Split genes • • • In eukaryotes genes contain non-protein coding regions called introns that interrupt the coding regions. A gene can contain as many as 500 introns that vary from 50-20,000 base pairs in length. The primary transcript must be edited to remove the introns before translation can occur. Mechanism of splicing (1) • • • • The key reaction in splicing is transesterification, where a phosphodiester bond is transferred to another OH group, breaking the RNA chain without any loss of energy. Introns always begin with GU and end with AG. The actual splicing reaction is catalysed by spliceosomes, which contain small nuclear RNA. SnRNAs are associated with proteins to form small ribonucleoprotein particles (snRNPs). A spiceosome contains about 300 different proteins and small nuclear RNAs U1, 2, 4, 5 and 6. Mechanism of splicing Ribozymes and the self-splicing of RNA • This was first shown to occur in rRNA from Tetrahymena and to require a divalent metal ion and guanosine but no protein (Fig. 24.14). • Initially the action was cis but subsequently ribonuclease P was shown to be a trans acting ribozyme. What is the biological status of introns? • • • In evolution introns may have accelerated the rate by allowing exon shuffling. Since exons often correspond to protein domains this is akin to developing a new piece of electronic equipment by rearranging whole boards rather than individual transistors. Introns facilitate shuffling because the exons must be retained intact during chromosome rearrangement whereas rearrangements which change introns are not likely to be harmful since they have no function. What is the origin of split genes? (1) • • One view is that prokaryote, intronless genes are the original form and that introns arose later by some unknown mechanism. Organisms which gained introns so that domain shuffling could occur would be advantaged in the long term. What is the origin of split genes? (2) • • • A second view is that primitive genes had external, non-coding regions. Fusion of genes resulted in intervening regions which were non-coding. The evolutionary pressure for rapid division then resulted in loss of introns in prokaryotes. Alternative splicing of two (or more) proteins for the price of one gene • A number of cases are known where different splicing patterns occur with the same initial transcript. These then lead to different mRNAs and different proteins. PolyA tail A general overview of the differences in the control of initiation and control of gene transcription in prokaryotes and eukaryotes • Unlike in prokaryotes RNA polymerase does not recognize sites on the DNA itself but binds because a large number of other proteins bind and recruit the polymerase. • A bacterium has about 4000 genes but a mammal about 30,000, with extensive differentiation into defined tissues. Nevertheless in nearly all cell types the DNA is the same. • Some genes are present in nearly all cells – these are housekeeping genes. Types of eukaryotic genes and their controlling regions • In eukaryotes there are three different RNA polymerases, I, II and III. • Type II transcribes nuclear protein coding genes. Type I codes for rRNA and type III for tRNA molecules. Type II eukaryotic gene promoters • Fig 24.16 shows the components of a type II promoter. • The basal elements include a short sequence of pyrimidines (initiator) and the TATA box, which is at about –25 base pairs. • Upstream control elements occur between –50 and –200 base pairs. These include the CAAT box, and the GC box. All genes require at least one of these but there is no standard pattern. See Fig. 24.17 for typical examples. Most transcription factors are regulated (1) • Many factors can exist in active and inactive states. Often activation is via phosphorylation or dephosphorylation. (See Figs. 24.18 a, b and c for examples). • Activation can also be associated with the movement of the transcription factor to the nucleus. • Factors have both DNA binding domains and a binding domains for other proteins (Fig. 23.8). Most transcription factors are regulated (2) • Commonly protein kinase A activates a transcription factor by phosphorylation. This occurs in response to a rise in cAMP. Regulation chromatin