Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Signal transduction wikipedia , lookup
Enzyme inhibitor wikipedia , lookup
Ultrasensitivity wikipedia , lookup
Metabolic network modelling wikipedia , lookup
Gene regulatory network wikipedia , lookup
Lipid signaling wikipedia , lookup
Biochemical cascade wikipedia , lookup
Lactate dehydrogenase wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Biosynthesis wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Citric acid cycle wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
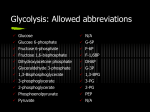
Carbohydrate metabolism Intermediary Metabolism Elizabeth F. Neufeld Suggested reference: Champe, Harvey and Ferrier, Lippincott’s Illustrated Reviews – Biochemistry, 3rd Edition 1 Kinetic properties of glucose transporters GLUT-2 Uptake in liver and pancreas b-cells is proportional to plasma concentration GLUT-1 GLUT-3 Uptake in brain is independent of plasma concentration over physiological range Km = concentration at which half maximum rate of transport occurs (1/2 Vmax) 2 GLUT4 activity is regulated by insulindependent translocation Intracellular pool of GLUT4 in membranous vesicles translocate to the cell membrane when insulin binds to its receptor. The presence of more receptors increases the Vmax for glucose uptake (does not affect Km). When insulin signal is withdrawn, GLUT4 proteins return to their 3 intracellular pool. GLUT4 is present in muscle and adipose tissue. Fate of glucose in the liver Glucose GLUT2 Glucose Glucokinase Glucose-6-P Glycogen synthesis Pentose phosphate Glycolysis 4 Glucokinase vs. Hexokinase Glucokinase: Km = 10 mM, not inhibited by glucose 6-phosphate. Present in liver and in pancreas b cells. Hexokinase: Km= 0.2 mM, inhibited by glucose 6phosphate. Present in most cells. 5 Glucokinase vs. Hexokinase • Hexokinase has low Km and therefore can efficiently use low levels of glucose. But is quickly saturated. • Glucokinase is found in liver and b-cells of pancreas • Glucokinase allows liver to respond to blood glucose levels • It has a high Km, so it does not become saturated till very high levels of glucose are reached • At low glucose levels, very little taken up by liver, so is spared for other tissues. • Not inhibited by glucose 6-phosphate, allowing accumulation in liver for storage as glycogen 6 Glucose action in the b-cell Glucose enters the b-cell as blood glucose concentration rises. Glycolysis to generate ATP closes K+ channels in the cell membrane, stopping outward transport, and opening Ca+ channels. Inward flux of Ca+ causes exocytosis of insulincontaining secretory vesicles. Glucose also stimulates synthesis of new insulin. 7 Fate of glucose in muscle Insulin + Glucose GLUT4 Glucose Hexokinase Glucose-6-P Glycogen synthesis Glycolysis 8 Glycogen accumulation in muscle 9 Fate of glucose in adipocytes Lipoproteins + Insulin + Glucose GLUT4 Insulin LPL Fatty Glucose Hexokinase acids Glucose-6-P Glycerol-3-P Insulin Triglycerides 10 How is metabolism regulated? Two broad classes of pathways • Catabolic – break down molecules to generate energy • Anabolic - require energy for synthesis of molecules The two pathways are kept distinct by regulatory mechanisms and/or sequestration in different cell compartments. Pathways contain recurring enzymatic mechanisms • Oxidation-reduction reactions • Isomerization reactions • Group transfer reactions • Hydrolytic reactions • Addition or removal of functional groups 11 How is metabolism regulated? Movement Active transport Signal amplification Biosynthesis Oxidation of fuel molecules High ATP concentrations inhibit catabolic pathways and stimulate anabolic pathways 12 How is metabolism regulated? Fast mechanisms, for immediate changes Substrate concentration Allosteric regulation (feedback, feed forward) Phosphorylation-dephosphorylation Signals emanating from hormone action Slow mechanisms, for long-term changes Genetic regulation Response to diet and other environmental variables 13 How is metabolism regulated? long term effects Rapid effect Rapid effects 14 Overview of glucose metabolic pathways • Glycolysis: from G6P to pyruvate • Gluconeogenesis: from oxaloacetate to G6P • Glycogen synthesis: from G6P to glycogen • Glycogenolysis: from glycogen to G6P • TCA cycle The pathways must be carefully regulated to keep pathways going in opposite directions from proceeding simultaneously. 15 Regulation of glycolysis • Glycolytic flux is controlled by need for ATP and/or for intermediates formed by the pathway (e.g., for fatty acid synthesis). • Control occurs at sites of irreversible reactions • Phosphofructokinase- major control point; first enzyme “unique” to glycolysis • Hexokinase or glucokinase • Pyruvate kinase •Phosphofructokinase responds to changes in: • Energy state of the cell (high ATP levels inhibit) • H+ concentration (high lactate levels inhibit) • Availability of alternate fuels such as fatty acids, ketone bodies (high citrate levels inhibit) • Insulin/glucagon ratio in blood (high fructose 2,6bisphosphate levels activate) 16 Control points in glycolysis Glucose-6-P - hexokinase * 17 Why is phosphofructokinase, rather than hexokinase, the key control point of glycolysis? Because glucose-6-phosphate is not only an intermediate in glycolysis. It is also involved in glycogen synthesis and the pentose phosphate pathway. PFK catalyzes the first unique and irreversible reaction in glycolysis. 18 Phosphofructokinase (PFK-1) as a regulator of glycolysis PFK-1 fructose-6-phosphate fructose-1,6-bisphosphate PFK allosterically inhibited by: • High ATP lower affinity for fructose-6-phosphate by binding to a regulatory site distinct from catalytic site. • High H+ reduced activity to prevent excessive lactic acid formation and drop in blood pH (acidosis). • Citrate prevents glycolysis by accumulation of this citric acid cycle intermediate to signal ample biosynthetic precursors and availability of fatty acids or ketone bodies for oxidation. 19 Phosphofructokinase (PFK-1) as a regulator of glycolysis PFK-1 activated by: Fructose-2,6-bisphosphate (F-2,6-P2) PFK-2 F-6-P PFK-1 + F-1,6-P2 glycolysis F-2,6-P2 F-2,6-P2 Activates PFK-1 by increasing its affinity for fructose-6-phosphate and diminishing the inhibitory effect of ATP. 20 Phosphofructokinase-2 (PFK-2) is also a phosphatase (bifunctional enzyme) Bifunctional enzyme has two activities: • 6-phosphofructo-2-kinase activity, decreased by phosphorylation • Fructose-2,6-bisphosphatase activity, increased by phosphorylation kinase ATP ADP fructose-2,6-bisphosphate fructose-6-phosphate Pi phosphatase 21 Hormonal control of F-2,6-P2 levels and glycolysis Hormonal regulation of bifunctional enzyme • Glucagon (liver) or epinephrine (muscle) increase cAMP levels, activate cAMPdependent protein kinase. In liver, this leads to decreased F-2,6-P and inhibits glycolysis. The effect is opposite in muscle; epinephrine stimulates glycolysis. Phosphorylation of PFK2 by • Insulin decreases cAMP, protein kinase activates increases F-2,6-P stimulates its phosphatase activity on glycolysis. F2,6P in liver. 22 GLUCOSE GK G-6-Pase G-6-P F-6-P Gluconeogenesis FBPase 1 Glycolysis PFK 1 F-1,6-P2 P-ENOLPYRUVATE PEPCK OXALOACETATE PK PYRUVATE 23 GLUCOSE GK G-6-Pase G-6-P F-6-P Gluconeogenesis FBPase 1 Glycolysis PFK 1 F-1,6-P2 Decrease Hepatic Glucose Output Increase Hepatic Glucose Utilization P-ENOLPYRUVATE PEPCK OXALOACETATE PK PYRUVATE 24 GLUCOSE GK G-6-Pase G-6-P F-6-P Gluconeogenesis FBPase 1 Glycolysis PFK 1 F-1,6-P2 Increase Hepatic Glucose Output Decrease Hepatic Glucose Utilization P-ENOLPYRUVATE PEPCK OXALOACETATE PK PYRUVATE 25 F-6-P / F-1,6-P 2 SUBCYCLE G-6-P F-6-P FBP ase 2 PFK 2 F-2,6-P2 - + PFK 1 FBPase 1 F-1,6-P 2 + PK 26 The bifunctional enzyme Fructose-6-P Fructose-2,6-bis-P FBP ase 2 Fructose-6-P PFK 2 P Fructose-2,6-bis-P 27 The bifunctional enzyme Fructose-6-P Fructose-2,6-bis-P FBP ase 2 Fructose-6-P PFK 2 P Fructose-2,6-bis-P Phosphorylation of PFK2 by PKA promotes gluconeogenesis 28 The bifunctional enzyme Fructose-6-P Fructose-2,6-bis-P FBP ase 2 Fructose-6-P PFK 2 Fructose-2,6-bis-P Double mutant, blocks phosphorylation of PFK2 and phosphatase activity of FBPase2 29 The bifunctional enzyme Fructose-2,6-bis-P Fructose-6-P FBP ase 2 PFK 2 Fructose-6-P Hepatic overexpression of the double mutant results in a gene expression profile consistent with the fed state, and protection from Type I and II diabetes Fructose-2,6-bis-P Increased PFK1, Increased glycolysis, Fed State 30 Gluconeogenesis • Mechanism to maintain adequate glucose levels in tissues, especially in brain (brain uses 120 g of the 160g of glucose needed daily). Erythrocytes also require glucose. • Occurs exclusively in liver (90%) and kidney (10%) • Glucose is synthesized from non-carbohydrate precursors derived from muscle, adipose tissue: pyruvate and lactate (60%), amino acids (20%), glycerol (20%) 31 Gluconeogenesis takes energy and is regulated Converts pyruvate to glucose Glucose-6-P - hexokinase Glucose 6-phosphatase Gluconeogenesis is not simply the reverse of glycolysis; it utilizes unique enzymes (pyruvate carboxylase, PEPCK, fructose-1,6bisphosphatase, and glucose-6-phosphatase) for irreversible reactions. 6 ATP equivalents are consumed in synthesizing 1 glucose from pyruvate in this pathway 32 Irreversible steps in gluconeogenesis • First step by a gluconeogenic-specific enzyme occurs in the mitochondria Pyruvate pyruvatecarboxylase oxaloacetate • Once oxaloacetate is produced, it is reduced to malate so that it can be transported to the cytosol. In the cytosol, oxaloacetate is subsequently dexcarboxylated/phosphorylated by PEPCK (phosphoenolpyruvate carboxykinase), a second enzyme unique to gluconeogenesis. The resulting phosphoenol pyruvate is metabolized by glycolysis enzymes in reverse, until the next irreversible step 33 Irreversible steps in gluconeogenesis (continued) • Fructose 1,6-bisphosphate + H2O Fructose 1,6Bisphosphatase (FBPase) fructose-6-phosphate + Pi • In liver, glucose-6-phosphate can be dephosphorylated to glucose, which is released and transported to other tissues. This reaction occurs in the lumen of the endoplasmic reticulum. Requires 5 proteins! 1) G-6-P transporter 2) Ca-binding stabilizing protein (SP) 3) G-6-Pase 4) Glucose transporter 5) Pi transporter 34 Gluconeogenesis and Glycolysis are reciprocally regulated • Fructose 1,6-bisphosphatase is main regulatory step in gluconeogenesis. • Corresponding step in glycolysis is 6-phosphofructo-1-kinase (PFK-1). • These two enzymes are regulated in a reciprocal manner by several metabolites. Fructose-6-phosphate + Citrate Citrate AMP + F 2,6-BP + 6-phosphofructo -1-kinase Fructose 1,6-bisphosphatase - AMP - F 2,6-BP Fructose 1,6-bisphosphate Reciprocal control—prevents simultaneous reactions in same cell. 35