Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Apical dendrite wikipedia , lookup
Synaptic gating wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Holonomic brain theory wikipedia , lookup
Optogenetics wikipedia , lookup
Development of the nervous system wikipedia , lookup
Nervous system network models wikipedia , lookup
Neuroregeneration wikipedia , lookup
Biological neuron model wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Neuroanatomy wikipedia , lookup
Synaptogenesis wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
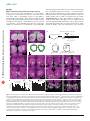
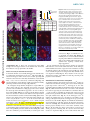
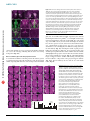
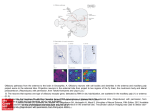
ARTICLES © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience Antagonistic roles of Wnt5 and the Drl receptor in patterning the Drosophila antennal lobe Ying Yao1,5, Yuping Wu2,5, Chong Yin2,5, Rie Ozawa1, Toshiro Aigaki3, Rene R Wouda4, Jasprina N Noordermeer4, Lee G Fradkin4 & Huey Hing1,2 Numerous studies have shown that ingrowing olfactory axons exert powerful inductive influences on olfactory map development. From an overexpression screen, we have identified wnt5 as a potent organizer of the olfactory map in Drosophila melanogaster. Loss of wnt5 resulted in severe derangement of the glomerular pattern, whereas overexpression of wnt5 resulted in the formation of ectopic midline glomeruli. Cell type–specific cDNA rescue and mosaic experiments showed that wnt5 functions in olfactory neurons. Mutation of the derailed (drl) gene, encoding a receptor for Wnt5, resulted in derangement of the glomerular map, ectopic midline glomeruli and the accumulation of Wnt5 at the midline. We show here that drl functions in glial cells, where it acts upstream of wnt5 to modulate its function in glomerular patterning. Our findings establish wnt5 as an anterograde signal that is expressed by olfactory axons and demonstrate a previously unappreciated, yet powerful, role for glia in patterning the Drosophila olfactory map. The perception of odors requires that stimulus information be systematically organized in a ‘map’ in the olfactory bulb. Indeed, axons of olfactory receptor neurons (ORNs) expressing a given odorant receptor sort out from other axons and terminate specifically in one of a field of discrete synaptic structures, termed glomeruli, in the olfactory bulb1. Previous studies have shown that the pattern of glomeruli, or olfactory map, is a direct result of precise axon pathfinding and synaptogenesis2,3. Recent work in several species has identified a number of molecules that are necessary for the proper development of the olfactory map. These molecules include transmembrane proteins such as Ncam-180, Neuropilin-1, Plexin A, Robo, the odorant receptors, Dscam and Cadherin, as well as cytoplasmic signaling molecules such as Src, Fyn, Dock and Pak4–11. An attempt to integrate these diverse mechanisms has led to a hierarchical model of ORN axon targeting, wherein ORN axons are sequentially guided toward their postsynaptic targets with an increasing degree of precision5,8. A wealth of evidence shows that ingrowing ORN axons are important for directing the development of the glomeruli in Drosophila and other species. First, genetic or surgical disruption of the ORN axons blocks the formation of the glomeruli, indicating that ORN axons are necessary for glomerular development9,12,13. Second, when the LIM kinase 1 (Limk1) gene is overexpressed in the ORNs, or when the ORNs are transplanted to ectopic sites, the axons direct the formation of glomeruli in ectopic positions, indicating that the presence of ORN axons is also sufficient for glomerular development13–16. It has been suggested that ORN axons have an intrinsic ability to specify glomerular development in their target tissues17, but the underlying mechanisms for this are unknown. The Drosophila olfactory system is highly suited for use in unraveling the mechanisms of olfactory map development. The anatomy and development of the fly antennal lobe closely resembles that of the olfactory bulb of mammals, but is vastly simpler, containing only 43 glomeruli compared with B2,000 in the mouse. We conducted an overexpression screen for genes that disrupted the stereotyped anatomy of the Drosophila antennal lobe. We identified wnt5 as a candidate regulator of antennal lobe development. The wnt5 gene encodes a member of the large, evolutionarily conserved Wnt family of secreted proteins, which have well-established roles in embryonic patterning, cell proliferation and cell differentiation18,19. Recent studies have shown that Wnt5 functions as a repulsive cue that routes axons through the correct commissures of the embryonic ventral midline20 and regulate their fasciculation21, and is required for stabilizing a subset of axons in the adult brain22. We show here that wnt5 functions in ORNs to regulate the patterning of the olfactory glomeruli. In the absence of wnt5, ORN axons do not target properly, and the glomerular map is disorganized. We also provide evidence that Drl, a Ryk-family receptor tyrosine kinase previously demonstrated to act as a repulsive neuronal Wnt5 receptor in the embryonic CNS20, functions in glia to inhibit Wnt5 signaling via its extracellular Wnt inhibitory factor (WIF) domain. Our results demonstrate that Wnt5 is an anterograde signal by which ORN axons specify the organization of the olfactory map and that Drl, presented by glial cells, is a powerful modulator of this signal. 1Neuroscience Program and 2Cell and Developmental Biology, University of Illinois at Urbana-Champaign, 601 South Goodwin Avenue, Urbana, Illinois 61801, USA. Sciences, Tokyo Metropolitan University, 1-1 Minami-osawa, Hachioji-shi, Tokyo 192-0397, Japan. 4Laboratory of Developmental Neurobiology, Department of Molecular Cell Biology, Leiden University Medical Center, Einthovenweg 20, 2300 RC Leiden, The Netherlands. 5These authors contributed equally to this work. Correspondence should be addressed to H.H. ([email protected]). 3Biological Received 10 July; accepted 17 September; published online 14 October 2007; doi:10.1038/nn1993 NATURE NEUROSCIENCE ADVANCE ONLINE PUBLICATION 1 ARTICLES © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience RESULTS ORNs overexpressing wnt5 disrupt antennal lobe structure From a screen of 3,996 P{GS} Drosophila lines23, we found that the overexpression of wnt5 (P{GS1}1192) under the control of the SG18.1Gal4 driver, which is preferentially expressed in the ORNs9,24, profoundly altered the anatomy of the antennal lobe (Fig. 1). In the wild-type adult expressing GFP under the control of SG18.1-Gal4, the antennal lobes were spherical (B70 mm diameters) and were connected by a commissure B20 mm thick (Fig. 1a). In the wnt5-overexpressing animals, the antennal lobes were malformed and glomerulus-like a b structures appeared in the commissure in 79% (42/53) of the antennal lobes, substantially enlarging the structure to B60 mm in thickness (Fig. 1b). We observed that a large percentage of the Or22a, Or43b and Or47b glomeruli in the wnt5-overexpressing animals were split into smaller subunits, many of which were displaced into the midline (Supplementary Fig. 1 online). The termination of ORN axons in ectopic positions indicates that ORN axons were misguided in the wnt5-overexpressing animals. Examination of the projection neurons showed that their dendritic arbors were disrupted, and a subset of them invaded the midline, where they innervated the ectopic glomeruli c P{GS1}1024, 1039 and 1192 insertion -glutamyltranserase gene wnt5 gene wnt5 deletion line WT SG18.1 > P{GS1} 1192 e d f wnt5 WT Or43b Or47a & Or47b D wnt5 g h WT Or71a l Or43b o wnt5 60 50 40 30 20 10 0 Between two glomeruli * *** ** Or71a p 120 WT wnt5 From glomerulus to the top of AL *** *** *** * Splitting in wnt5 Crossing defect in wnt5 100 Or47a & Or47b n 720K wnt5 q r WT wnt5 Or47a & Or47b 60 40 20 a b 59 c 42 a 72 0K 47 b 67 d 67 b 71 43 d b wnt5 WT 47 67 a c a- 47 42 b 59 a 67 b 71 720K 80 0 43 WT Or47b in WT Or47b in wnt5 j m Percentage (%) wnt5 Or47a in WT Or47a in wnt5 i Or43b k Distance (µm) Or43b in WT Or43b in wnt5 M Figure 1 The olfactory map is disrupted in the wnt5 mutant. (a–r) Preparations of wild-type (WT) flies and wnt5 mutants were stained with nc82 (magenta) and antibody to GFP (green). Antennal lobes of wild-type (a), wnt5-overexpressing (P{GS1}1192) (b) and wnt5400 (d) animals were visualized using GFP driven by SG18.1-Gal4. Commissures are indicated with arrows. (c) Schematic diagram of the wnt5 locus showing the P{GS} lines recovered from our screen and the wnt5400 deletion breakpoints. (e) Superimposed outlines of antennal lobes from wild-type flies and the wnt5400 mutants (n ¼ 12). The wnt5400 mutant antennal lobes were flattened dorsomedially (D, dorsal; M, medial), producing a characteristic heart-shape appearance. (f) Plots of the positions of Or43b, Or47a and Or47b glomeruli in the wild type and wnt5400 mutant. Preparations of wild-type (g–j) and wnt5400 mutant (k–n) antennal lobes in which specific glomeruli are highlighted with GFP. In the wnt5400 mutant, the glomeruli were closer to the antennal lobe dorsal edge or were split into smaller structures (arrowheads), compared with wild type. ORN axons frequently took meandering paths (arrow) to their targets and failed to cross the midline (double arrowheads). (o) Histograms showing the distances between various glomeruli and the antennal lobe dorsal edge or between two glomeruli in the same antennal lobes (ALs). Values are mean ± s.e.m. (p) Quantification of the glomerular-splitting and midline-crossing defects in the wnt5400 mutant. Single confocal sections showing the projection neuron dendritic arrangement in the wild type (q) and wnt5400 mutant (r), labeled by GH164-Gal4 driving UAS-mGFP. For each group, n ¼ 25–40. Scale bar, 20 mm. 2 ADVANCE ONLINE PUBLICATION NATURE NEUROSCIENCE a WT b 26 h wnt5 c © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience WT e Percentage (%) ARTICLES 26 h 120 100 80 60 40 20 0 wnt5 f Crossing midline Project dorsally Absent 40–50 hAPF 50 h g h j k 18 4 50 18 0 22 0 28 0 0 0 9 18 h i Adult Adult 40–50 hAPF (n = 18) (n = 26) (n = 50) (n = 55) d 50 h WT wnt5 18 h (Supplementary Fig. 1). Thus, wnt5 overexpression in the ORNs disrupted the stereotyped structure of the antennal lobe and elicited the formation of ectopic glomeruli at the midline. wnt5 is necessary for antennal lobe structure To determine whether wnt5 normally functions in the antennal lobes, we examined the antennal lobes of the wnt5400 null mutant (Fig. 1c), which is homozygous viable21. In the wnt5400 mutant expressing GFP under the control of SG18.1-Gal4, many antennal lobes (37/55 antennal lobes, B67%) were not connected by commissures (Fig. 1d,p) and appeared misshapen; being flattened dorsally and elongated ventrally, producing a characteristic heart shape (Fig. 1d,e,k–n). We investigated the glomerular arrangement of eight ORN subclasses. In the wild type, these glomeruli are located at stereotyped positions in the antennal lobe25,26 (Fig. 1g–j). In the wnt5400 mutant, the dorsal edge of the antennal lobe appeared to be closer to many of these glomeruli (Fig. 1f,k–n,o). To test the possibility that the antennal lobes were distorted, we measured the distance between glomeruli in the same antennal lobe (Or47a to Or47b, and Or67d; Fig. 1j,n,o). In the wild type, the distance between the Or47a and Or47b glomeruli was 22.51 ± 1.60 mm (mean ± s.e.m., n ¼ 32). In the wnt5400 mutant, this distance decreased to 8.70 ± 2.73 mm (n ¼ 32, P ¼ 0.0001 compared with wild type). Thus, the wnt5400 antennal lobes showed a characteristic ‘collapse’ of the dorsomedial region of the antennal lobe (Fig. 1e). Examination of the ORN fibers also showed that ORN axons frequently took meandering paths toward their targets (arrows, Fig. 1k,l), looped back on the ipsilateral glomeruli (double arrowheads, Fig. 1n), stalled before the commissure, projected aberrantly to dorsal regions of the NATURE NEUROSCIENCE ADVANCE ONLINE PUBLICATION Figure 2 Wnt5 functions during antennal lobe development and localizes to projection neuron dendrites. (a–d) Preparations of wild type (a,c) and wnt5400/Y hemizygote (b,d) in which GFP was driven by SG18.1-Gal were stained with nc82 (magenta) and GFP (green) antibodies. Figures show projected confocal sections. (a,c) In the wild type, contralateral axons crossed the midline (arrow) and the antennal lobes had spherical shapes. (b,d) In the mutant, contralateral axons projected aberrantly toward dorsal regions of the brain (arrows). (e) Quantification of the commissural defects seen in the wnt5400 mutant at 40–50 hAPF or adulthood. (f–h) Preparations of 18-hAPF pupal brains expressing GFP (green) under the control of the pan-olfactory driver pblGal4 were stained with antibodies to Wnt5 (red) and Elav (blue). Figures show single confocal sections. Asterisks indicate three (of the four) spheroidal structures that accumulated Wnt5 and are surrounded by masses of ORN axons. (i–k) Preparations of 18-hAPF pupal brains expressing GFP (green) under the control of GH146-Gal4 were stained with antibodies to Wnt5 (red) and Elav (blue). Projection neuron dendrites overlapped largely with the niduses of Wnt5 staining (red). Dorsal is up and lateral is to the right in f–k. Scale bars, 20 mm in a–d and 10 mm in f–k. brain (arrows, Fig. 2) or terminated on ectopic glomeruli (arrowheads, Fig. 1k,l,n,p). In summary, the loss of wnt5 resulted in aberrant targeting of the ORN axons and characteristic shifts of dorsomedial glomeruli leading to the antennal lobes acquiring a distinct heartshaped appearance. We also examined the projection neuron dendrites by expressing GFP under the control of the GH146-Gal4 driver. In the wild type, GH146-Gal4 labeled a subset of projection neuron dendrites, revealing an invariant pattern in the antennal lobe neuropil (Fig. 1q). In the wnt5400 mutant, this pattern was disrupted and many dendritic arbors were displaced ventrally (Fig. 1r). Thus, mutation of the wnt5 gene disrupted the targeting of the ORN axons and the positioning of the projection neuron dendrites. wnt5 functions in antennal lobe development To determine when the observed defects arise in the wnt5400 mutant, we evaluated the antennal lobe structure of the wnt5400 mutant during the pupal stages. In the control wnt5400/+ heterozygotes at 26 h after puparium formation (hAPF), the antennal lobe had a smooth neuropil (Fig. 2a). ORN axons had arrived at the outer surface of the antennal lobes and were projecting across the midline in a commissure that was B5 mm in width. At 50 hAPF, the antennal lobe neuropil was well partitioned into glomeruli and a thick commissure connected the left and right antennal lobes (Fig. 2c). In the hemizygous wnt5400/Y mutant, the antennal lobes were oval shaped at 26 hAPF, as in the wild type, although in many brains, axons appeared to project dorsally instead of across the midline (Fig. 2b). At 50 hAPF, the antennal lobes appeared misshapen, with many showing the distinctive heart shape seen in the adult (Fig. 2d). Contralateral axons failed to decussate and instead projected dorsally in 85% of the brains examined (22/26; Fig. 2e). By the adult stage, many of the brains no longer showed dorsal projections, indicating that the wayward axons had likely retracted or degenerated (Fig. 2e). These results indicate that wnt5 3 ARTICLES a WT WT wnt5/Y; SG > wnt5 wnt5 wnt5/Y;SG > wnt5 f wnt5/Y;720K > wnt5 h g Or47b in WT Or47b in wnt5/Y mutant Or47b in rescue of wnt5/Y mutant D M *** *** * WT wnt5/Y wnt5/Y; wnt5/Y; SG > wnt5 72OK > wnt5 j wnt5 wnt5/Y; SG > wnt5 WT and wnt5/Y wnt5/Y; 72OK > wnt5 wnt5/Y;GH146 > wnt5 Splitting in WT k l WT wnt5 WT d WT wnt5 50 48 46 44 42 40 ** Percentage (%) i 30 25 20 15 10 5 0 wnt5/Y wnt5/Y; 720K > wnt5 c Distance between 720K and the top of AL (µm) Distance between Or47b and the top of AL (µm) © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience e b 80 Splitting in wnt5 Crossing defect in WT Crossing defect in wnt5 60 40 20 0 Figure 3 wnt5 functions in the ORNs. (a–h) Antennal lobes from wild-type flies (a), wnt5400 mutants (b) and wnt5400 mutants expressing UAS-wnt5 under the control of various drivers (c,d,h) were stained with antibody to GFP (green), to reveal the Or47b axons, and nc82 (magenta). Expression of UAS-wnt5 with SG18.1-Gal4 (c) and 72OK-Gal4 (d) rescued the wnt5400 mutant phenotype, but expression with GH146-Gal4 did not (h). (e) Histograms show the distances between the Or47b glomerulus and the top edge of the antennal lobe in the various genotypes. (f) Superimposed antennal lobe outlines from various genotypes (n ¼ 12). (g) Plots showing the Or47b glomerulus positions in the various genotypes. (i,j) Representative antennal lobes with MARCM clones of wild-type and wnt5400 mutant 72OK neurons. In antennal lobes with wnt5400 mutant 72OK axons, the 72OK glomerulus was split into smaller structures (arrowheads) and contralateral axons (arrows) looped back on the ipsilateral targets instead of crossing the midline. (k) A plot of the positions of the 72OK glomerulus in the wild-type and wnt5400 mosaic animals. (l) Quantification of the distance between the 72OK glomerulus and the top edge of the antennal lobe, the splitting of glomerulus and the absence of the contralateral tract in wild-type and wnt5400 mosaic animals. Values are mean ± s.e.m. All figures show projected confocal sections. SG ¼ SG18.1-Gal4; 72OK ¼ 72OK-Gal4. Scale bar, 20 mm. functions during development to regulate ORN axon projection and antennal lobe development. Wnt5 protein is localized in the developing antennal lobes To ascertain the distribution of Wnt5 protein, we stained pupal brains with an antibody to Wnt5 (ref. 21) that did not stain wnt5400 mutant brains (Supplementary Fig. 2 online), which demonstrates its specificity. We stained pupae expressing GFP under the control of pbl-Gal4, a pan–olfactory neuron driver11. We observed ORN axons penetrating into the antennal lobe as early as 18 hAPF (arrows, Fig. 2f,g). At this time, Wnt5 immunolabeling was seen in four spheroidal structures (B7 mm in diameter, asterisks, Fig. 2h), each surrounded by profuse axonal terminals (Fig. 2f). This pattern of Wnt5 expression remained relatively unchanged up to 42 hAPF (Supplementary Fig. 2). To identify the spheroidal structures, we stained 18-hAPF animals expressing GFP under the control of GH146-Gal4, which labels the projection neuron dendrites, with the Wnt5 antibody. Wnt5 immunoreactivity largely coincided with the projection neuron dendrites, indicating that Wnt5 was localized to projection neuron dendrites during glomerular development (Fig. 2i–k). Besides the dendritic localization, we show below that Wnt5 also accumulated in the region of the antennal commissure in the drl2 mutant. Wnt5 staining was greatly reduced by 70 hAPF, when glomerular development was largely complete (data not shown). Our immunolocalization results indicate that Wnt5 is localized to incipient glomeruli during the period of ORN axon targeting and glomerular development. 4 wnt5 is required in the ORNs To delineate the cell type in which wnt5 functions, we first restored wnt5 function to specific cell types in the wnt5400 mutant background. We employed a UAS-wnt5 transgene21, which expresses wnt5 at a low level and therefore does not disrupt antennal lobe development (Supplementary Fig. 1). Three criteria were specifically assessed: the shape of the antennal lobe, the position of the Or47b glomerulus and the presence or absence of the commissure. In the wnt5400 mutant, the antennal lobe was heart shaped (compare Fig. 3a,b), the Or47b glomerulus was located 14.44 ± 0.8 mm (n ¼ 14) from the dorsal edge of the antennal lobe compared with the wild-type glomerulus, which was located 24.66 ± 1.3 mm (n ¼ 20) from the dorsal edge (P o 0.0001; Fig. 3), and the commissure was present in only 30% of the brains. When we expressed UAS-wnt5 using the GH146-Gal4 driver, which is expressed in projection neurons, the mutant phenotype was not rescued (Fig. 3h). No rescue was observed when UAS-wnt5 was expressed under the control of MZ317-Gal4, which is expressed in glia (data not shown). When UAS-wnt5 was expressed under the control of SG18.1-Gal4, the wnt5400 mutant phenotype was rescued to a significant degree. The rescued antennal lobes were spherical, instead of heart-shaped (Fig. 3c,f), the Or47b glomerulus was located 20.12 ± 1.4 mm from the dorsal edge of the antennal lobe (n ¼ 24, P ¼ 0.0102 compared with mutant; Fig. 3e,g), and the antennal lobes were connected by a commissure in 55% (11/20) of the brains. Expression of UAS-wnt5 under the control of the 72OK-Gal4 driver, which is expressed in a subset of ORNs7, also significantly rescued the mutant ADVANCE ONLINE PUBLICATION NATURE NEUROSCIENCE ARTICLES Or43b Or67b Or42a b Or71a c d g h l WT a e f drl * * j k m n o q r WT p * drl s Percentage (%) © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience drl i 120 drl- mutant phenotype 100 Targeting Unbalance D t D 80 L 60 M L M 71a 43b 40 Figure 4 The olfactory map is disrupted in the drl mutant. (a–r) Antennal lobes of wild type (a–d, m–o) and drl2 mutants (e–l,p–r) were stained with antibody to GFP (green) and nc82 (magenta). (a–d) Wild-type antennal lobes expressing GFP in the Or43b, Or67b, Or42a and Or71a glomeruli. (e–l) drl2 mutant antennal lobes showing the corresponding glomeruli. Arrowheads indicate the split glomeruli, arrows indicate the wandering ORN axons and asterisks denote the ectopic midline glomeruli. Antennal lobes from wild-type flies (m) and drl2 mutants (p) expressing GFP (green) under the control of SG18.1-Gal4 were stained with nc82 (magenta). Ectopic protrusions were visible at the midline of the drl2 mutant (asterisk). In the wild-type adult, projection neuron dendrites, visualized with GH146-Gal4 driving GFP (green), were arranged in a stereotyped pattern (n) and never terminated at the midline (o). In the drl2 mutant, the dendritic pattern was severely deranged (q) and dendrites innervated ectopic glomeruli in the midline (asterisk, r). (s) Quantification of the targeting (dark bars) or the unbalanced (light bars) defects in the various ORN subtypes. (t) Plots of the positions of the ectopic Or43b and Or71a glomeruli (magenta dots) in the drl2 mutant. All panels, except for n, o, q and r, show projected confocal sections. Scale bars, 20 mm in a–d and 10 mm in e–h. 20 0 22a 43b 59c 42a 92a 47a 33c 67b 46a 71a 43a 47b Medial Lateral V phenotype. The rescued antennal lobes had a wild-type shape (Fig. 3d,f), the Or47b glomerulus was located 19.68 ± 1.2 mm from the dorsal edge of the antennal lobe (n ¼ 16, P ¼ 0.001 compared with mutant; Fig. 3e,g) and the antennal lobes were connected by a commissure in 77% (10/13) of the brains. Next, we investigated whether wnt5 is required in the ORNs for proper antennal lobe development. We employed the mosaic analysis with a repressible cell marker (MARCM) system27 to induce and examine clones of wild-type or wnt5400 homozygous 72OK cells. Large clones were examined, as small clones were likely to be rescued by Wnt5 secreted from wild-type cells adjacent to the clones. In control animals with wild-type clones, the antennal lobes had a spherical shape (Fig. 3i). The wild-type 72OK ORNs projected to their expected positions (49.14 ± 3.5 mm from dorsal surface of antennal lobe, n ¼ 14; Fig. 3k,l) and formed glomeruli with stereotyped shapes. Their contralateral axons formed a distinct fascicle that projected normally across the midline. In animals bearing wnt5400 clones, the antennal lobes were misshapen, with a heart-shaped appearance (Fig. 3j). The mutant 72OK ORNs projected to a slightly more dorsal position than that of the control (44.16 ± 4.5 mm, n ¼ 18, P ¼ 0.002; Fig. 3k,l) and formed distorted and split glomeruli (Fig. 3j,l). Their contralateral axons frequently appeared defasciculated and (72.2%, 13/18 brains) failed to project across the midline (Fig. 3j,l). Our cell type–specific cDNA rescue and mosaic experiments therefore indicated that wnt5 functioned in the ORNs for appropriate antennal lobe patterning. drl is necessary for patterning the glomerular map The capacity of Wnt5 to regulate glomerular patterning focused our attention on its potential downstream signaling pathway. Drl has been shown to act as a receptor for the repulsive Wnt5 cue in axons that cross NATURE NEUROSCIENCE ADVANCE ONLINE PUBLICATION V the embryonic midline20. To determine whether drl also functions in the antennal lobe, we examined the antennal lobes of the drl2 null mutant28 (Fig. 4). In 39% (22/56) of the mutant brains, ectopic glomeruli developed at the midline (asterisks, Fig. 4a–f,m,p,r), whereas abnormal protrusions extended from the dorsal-medial corner of the antennal lobes of the remainder (61%). In each of the 12 subclasses of ORNs that we analyzed, ectopic midline glomeruli were seen at low frequencies, suggesting that all subclasses have an equal tendency to terminate at the midline. Plotting the positions of the Or43b and Or71a glomeruli in different antennal lobes revealed no consistent pattern, indicating that the glomeruli were randomly situated in the mutant (Fig. 4t). In addition to the glomerular defects, ORN axons frequently took circuitous routes to their targets (arrows, Fig. 4g,i–k). Strikingly, a number of axons terminated unilaterally (confirmed by unilateral antennal ablations, Supplementary Fig. 3 online), resulting in the loss of presynaptic structures from a single antennal lobe (Fig. 4i–k). To better assess the data, we grouped the glomerular positioning and splitting defects as ‘targeting defects’ (Fig. 4e–h,l) and the unilateral loss (or reduction) of glomeruli as ‘unbalanced defects’ (Fig. 4i–k). Both types of defects were present in each of the 11 ORN subclasses (Fig. 4s). We also inspected the organization of the projection neurons in the drl2 mutant using the GH146-Gal4 marker. Unlike in the wild-type adult, where GH146-Gal4-expressing cells arborized in specific glomeruli, the organization of the dendritic arbors appeared chaotic in the drl2 mutant adult (Fig. 4n,q), and a subset of arbors even invaded the midline, where they targeted the ectopic midline glomeruli (Fig. 4o,r). Loss of drl therefore leads to disruption of the glomerular arrangement and midline invasion by projection neuron dendrites. This midline invasion of projection neuron dendrites is highly reminiscent of the wnt5 overexpression phenotype (Supplementary Fig. 1). When we examined the antennal lobes of the drl2 mutant at 40 hAPF (Fig. 5), we 5 ARTICLES a WT Figure 5 Drl functions during antennal lobe development and localizes to TIFR glia. (a) In the wild type at 40 hAPF, projection neuron dendrites, visualized with GH146-Gal4 driving GFP (green), were confined to the antennal lobes. (b) An equivalent section of drl2 mutant showed projection neuron dendrites extending toward the midline (arrows). (c–g) Frontal optical sections of a wild-type 30-hAPF pupal brain expressing GFP (green) under the control of Repo-Gal4 stained with an antibody to Drl (magenta). Antennal lobes are indicated by the dotted outlines. (c–e) The Drl antibody intensely stained the TIFR (brackets). (f,g) A more posterior section, at the level of the antennal commissure, shows a bundle of glial processes that connects the two antennal lobes and also expressed the Drl protein (arrows). (h) No Drl staining was found in the drl2 mutant brain. (i–k) Antennal lobe from a 30-hAPF wild-type animal expressing GFP (green) under the control of GH146-Gal4 stained with antibodies to Drl (red) and Elav (blue). Drl distribution colocalized largely with the projection neuron dendrites. Dorsal is up and lateral is to the right in i–k. All the panels show single confocal sections. Scale bars, 10 mm. b 40 h drl 40 h c d e f g h i j k glial cells. At 30 hAPF, GFP was highly expressed by the transient interhemispheric fibrous ring (TIFR)29, a toroidal glial structure lying in the mid-sagittal plane of the brain (brackets, Fig. 5c,e). The ventral edge of the TIFR is closely associated with and bridges the antennal lobes. In more posterior sections, at the level of the antennal commissure, a thick bundle of glial processes was seen to pass from one antennal lobe to the other (Fig. 5f). The TIFR was strongly stained by the Drl antibody (Fig. 5d), including the bundle of glial processes linking the antennal lobes (Fig. 5g). We next stained brains from animals expressing GFP under the control of GH146-Gal4. At 30 hAPF, GFP highlighted the projection neuron cell bodies and their dendritic arbors in the antennal lobe neuropil (Fig. 5j). Drl immunolabeling largely overlapped with the dendritic GFP staining, indicating that Drl was localized to a large subset of the projection neuron dendrites (Fig. 5i–k). The Drl antibody did not stain the drl2 null mutant brains (Fig. 5h). 30 h observed fine dendritic processes growing into the midline, which did not occur in the wild type, indicating that the defects arose during development (Fig. 5a,b). Drl is localized to glial cells and projection neurons To ascertain the localization of the Drl protein in the olfactory system, we stained developing brains with an antibody to Drl. We first stained animals expressing GFP under the control of Repo-Gal4, which labels Or46a Or43b Or22a Or47b b c d e f g h i j k l drl drl WT a Repo > drl n Repo > drl Repo > drl o p drl m Repo > drl Repo > drl ∆cyto Repo > drl ∆cyto r Repo > drl ∆cyto s Repo > drl ∆cyto Or43b drl q Repo > drl ∆wif v u 40 WT 6 drl FL GH146 FL K371A Repo ∆INTRA 43b 46a 47b 22a 43b 46a 43b 47b 22a 43b 46a 43b 46a 47b 22a 43b 46a 0 47b 20 22a GH146 > drl 60 43b GH146 > drl Targeting Unbalance 80 46a t Repo > drl K371A 100 drl-mutant phenotype Percentage (%) Repo > drl ∆wif drl © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience 30 h ∆WIF Figure 6 Drl functions in glial cells and its WIF domain is essential. (a–u) Preparations of wildtype flies (a–d) and drl2 mutants (e–u) expressing Or46a-mGFP, Or43b-mGFP, Or22a-mGFP and Or47b-mGFP were stained with antibody to GFP (green) and nc82 (magenta). (a–d) Glomeruli had predictable shapes and were located in invariant topographic positions in the wild type. (e–h) In the drl2 mutant, glomeruli were ectopically positioned, split or missing. (i–l) Expression of UAS-drl under the control of the Repo-Gal4 driver strongly rescued the mutant phenotype. Glomeruli were located in their stereotyped positions. (m–p) Expression of a truncated Drl protein lacking the cytoplasmic domain using the Repo-Gal4 driver rescued the mutant phenotype. (q,r) Expression of a truncated Drl protein lacking the extracellular WIF domain failed to rescue the mutant phenotype. Glomeruli were split or missing. (s) Expression of a ‘kinase-dead’ Drl protein (K371A) also rescued the mutant phenotype. (t,u) Expression of UAS-drl under the control of GH146-Gal4 partially rescued the mutant phenotype. Although the glomerular defects remained, protrusions from the antennal lobes were absent. (v) Quantification of the mutant defects found in the various genotypes. Dark bars represent targeting defects, whereas light bars represent unbalanced defects. FL, full-length Drl protein. All images are projections of confocal z-series. Scale bars, 20 mm. n Z 15. ADVANCE ONLINE PUBLICATION NATURE NEUROSCIENCE a * Drl’s extracellular domain regulates glomerular patterning Our ability to rescue the mutant phenotype allowed us to probe the functions of the different domains of the Drl protein in glomerular patterning. To determine whether the Drl kinase activity is required for antennal lobe development, we mutated the conserved K371 residue, which is essential for kinase activity32, to an alanine. Expression of the UAS-drlK371A transgene under the control of Repo-Gal4 strongly rescued the drl mutant phenotype (26% midline defects and 11% Or43b axon targeting defects compared with 100% and 35%, respectively, in the drl2 mutant; Fig. 6s), indicating that kinase activity is not NATURE NEUROSCIENCE ADVANCE ONLINE PUBLICATION * c * drl –/– drl –/–; SG18.1 > wnt5 SG18.1 > wnt5 d e f * * * g h i WT wnt5/Y +/Y;drl j k l –/– m wnt5/+;drl p –/– 100 50 44% 38% 12 18 8 18 13 0 –/– n WT * /– wnt5/Y;drl drl functions in glial cells To delineate the cell types in which drl functions, we sought to restore drl function to specific cell types in the drl2 mutant background. Because Drl localized, in part, to projection neuron dendrites, we employed the GH146-Gal4 driver, which is active in two thirds of all projection neurons30,31, to drive the expression of UAS-drl. We examined several criteria to assess genetic rescue (Fig. 6): the presence or absence of ectopic midline structures and the position and integrity of the Or22a, Or43b, Or46a and Or47b glomeruli (Fig. 6a–d). In the drl2 mutant, 100% of the brains showed aberrant midline structures (either midline glomeruli or protrusions) and 38%–94% of the brains showed defects in the positioning or integrity of the glomeruli (Fig. 6e–h,v). Expression of UAS-drl under the control of GH146-Gal4 partially rescued the mutant phenotype (58% targeting defects; Fig. 6t,u,v). To determine whether drl is required in the projection neurons, we generated and examined drl-mutant projection neuron MARCM single-cell clones. Loss of drl from single projection neurons, however, did not disrupt the projection neuron dendrite morphology or the stereotyped arrangement of the glomeruli in the antennal lobes (Supplementary Fig. 4 online). To further evaluate whether drl functions in neurons during antennal lobe development, we expressed UAS-drl under the control of elav-Gal4, a pan-neuronal driver, but this did not rescue the drl2 mutant phenotype (data not shown). Next, we tested whether expression of drl in glia would rescue the drl2 phenotype. Expression of UAS-drl under the control of the pan-glial driver RepoGal4 completely eliminated the midline defects (0%) and strongly reduced the occurrence of targeting defects to 11–24% for different glomeruli (Fig. 6i–l,v). We conclude that drl functions in the glia for the proper development of the antennal lobes. b W w T n +/ t5/Y Y; dr w l –/– nt dr 5/Y l– ; w /– n dr t5/+ l– ; Figure 7 drl functions upstream of wnt5 to inhibit wnt5 function. (a–f) Preparations of various genotypes expressing GFP under the control of SG18.1-Gal4 were stained for GFP (green) and nc82 (magenta). (a) Antennal lobes of the drl2 mutant. (b) Antennal lobes of the drl2 mutant expressing UAS-wnt5 under the control of SG18.1-Gal4. (c) Antennal lobes of the wild type expressing UAS-wnt5 under the control of SG18.1-Gal4. (d–f) Corresponding deep sections of a–c, revealing the large ectopic glomeruli (asterisks) of b and e compared with a and d. (g) Wild-type antennal lobes with invariant shape and position of Or47b glomeruli. (h) The wnt5400 antennal lobes had a characteristic heart shape and the Or47b axons failed to cross the midline (arrow). (i) In the drl2 antennal lobes, the Or47b glomerulus was split and ectopic glomeruli appeared at the midline (asterisk). (j) In the wnt5;drl double mutant, the antennal lobes had a heart-shaped appearance and the Or47b contralateral fibers failed to cross the midline (arrow). (k) Removal of a copy of wnt5 suppressed the drl mutant phenotype. The antennal lobes had wild-type shapes and ectopic midline structures were absent. (l) Quantification of the percentage of antennal lobes showing the failure of Or47b axons to cross the midline. (m–q) Superimpositions of the antennal lobe outlines from the various genotypes (n ¼ 6 for each genotype). Midline protrusions are indicated with an asterisk. a–c and g–k are projected confocal sections and d–f are single confocal sections. Scale bar, 20 mm. Midline crossing (%) © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience ARTICLES o wnt5/Y +/Y; drl –/– q wnt5/Y; drl –/– wnt5/+; drl –/– essential for Drl function in antennal lobe development. To ascertain whether the cytoplasmic domain is necessary for antennal lobe development, we used the UAS-drlDcyto transgene33, which encodes a truncated Drl protein bearing only the extracellular and transmembrane domains. Notably, UAS-drlDcyto retained substantial ability to rescue the drl mutant phenotype (33% midline defects, 23% targeting defects, Fig. 6m–p,v). To determine whether the Drl WIF domain is needed for antennal lobe development, we generated a UAS-drlDWIF transgene, which encodes a Drl protein that only lacks the WIF domain. The loss of the WIF domain abolished the ability of drl to rescue the mutant phenotype (89% midline defects and 56% targeting defects; Fig. 6q,r,v). Taken together, these results indicate that Drl regulates antennal lobe development largely through its Wnt5-binding WIF domain. wnt5 acts downstream of drl in antennal lobe patterning We next investigated the genetic relationship between wnt5 and drl. The similarity of the midline defects seen in the drl loss-of-function and wnt5 gain-of-function mutants suggested that the two genes might act antagonistically in antennal lobe development. To test this idea, we examined the effect of expressing a low level of wnt5 in the drl2 mutant background (Fig. 7). In the drl2 mutant, midline glomeruli were found in 39% of the animals, with most showing only protrusions from the dorsomedial corner of the antennal lobes (Fig. 7a,d). Expression of a single copy of UAS-wnt5 under the control of SG18.1-Gal4 in the wildtype background did not alter the antennal lobe structure (Fig. 7c,f and Supplementary Fig. 1). In contrast, expression of a single UAS-wnt5 in the drl2 mutant background resulted in the induction of large midline glomeruli in 83% of the animals (Fig. 7b,e). Thus, wnt5 function is 7 ARTICLES Figure 8 Wnt5 is localized in the antennal lobe WT drl drl;Repo > drl neuropil and midline commissure in the drl mutant animals. (a–f) Preparations of 36-hAPF pupae of wild-type flies (a,d) and drl2 mutants (b,c,e,f) expressing UAS-GFP under the control WT drl AL of Repo-Gal4 were stained for Wnt5 (red), GFP AL AL AL AL AL drl; Repo > drl (green) and Elav (blue). (c,f) drl2 mutant 120 *** coexpressing UAS-drl under the control of 100 *** Repo-Gal4. (a–c) Anterior optical sections 80 showing Wnt5 staining in the antennal lobe 60 neuropil. (d–f) Posterior sections showing Wnt5 staining in the commissure (dashed lines). The 40 TIFR is indicated by a bracket in a. (g) QuantiAL 20 AL AL AL AL AL fication of the levels of Wnt5 in the antennal lobe 0 36 h 36 h 36 h Superficial Commissure neuropils and commissures of the various geno30 h 36 h types. (h–k) Preparations of wild-type (h,i) and drl2 mutant (j,k) pupae expressing UAS-GFP under the control of GH146-Gal4 were stained for Wnt5 (red), GFP (green) and Elav (blue). Micrographs were taken at equivalent section planes (at the level of the commissure) and the antennal lobes are outlined with dashed lines. (j) At 30 hAPF in the drl2 mutant, Wnt5 was located medial to the antennal lobes (arrowheads), whereas Wnt5 remained in the confines of the antennal lobes in the wild type (h). (k) At 36 hAPF in the drl2 mutant, Wnt5 accumulation increased the antennal commissure (arrowhead). Projection neuron dendrites (arrows) began to transgress the antennal lobe boundaries and projected toward the region of Wnt5 accumulation, which was not seen in the wild type (i). All figures are of single confocal optical sections. Scale bar, 10 mm. b c Superficial a e f i j k WT h drl © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience Commissure d Percentage intensity of Wnt5 mAb staining (%) g strongly increased in the drl2 mutant background, which is consistent with the notion that drl antagonizes wnt5 during glomerular development and antennal lobe patterning. To determine the hierarchical order of wnt5 and drl in the genetic pathway regulating antennal lobe development, we constructed animals bearing null mutations in both genes and expressing GFP in the Or47b axons. Heterozygosity for drl2 had no effect on the homozygous wnt5400 (wnt5/Y; drl/+) mutant phenotype (data not shown). In contrast, heterozygosity for wnt5400 markedly suppressed both the dorsomedial antennal lobe protrusions and the glomerular defects that were typical of the drl2 mutant (wnt5/+; drl/drl; Fig. 7i,k,o,q). Homozygosity for both the wnt5400 and drl2 null mutations resulted in antennal lobes having the characteristic wnt5 mutant phenotype (wnt5/Y; drl/drl; Fig. 7g–j). The dorsal-medial corners of the antennal lobes were collapsed, producing a heart-shaped appearance (Fig. 7m–p), and the Or47b contralateral axons failed to cross the midline in 56% of the animals (compared with 0% in drl2 mutants and 62% in wnt5400 mutants; Fig. 7j,l). wnt5400 is thus epistatic to drl2, suggesting that wnt5 functions downstream of and is repressed by drl in the signaling pathway that regulates antennal lobe development. Wnt5 protein accumulates at the midline in the drl mutant The antagonistic relationship between drl and wnt5 led us to ask how they might function together to regulate antennal lobe development. The strong midline defects of the wnt5 and drl mutants and the localization of Drl to the TIFR suggest a role for these proteins at the midline (Fig. 8). As mentioned above, we observed Wnt5 immunoreactivity in the wild-type antennal lobe neuropil at 36 hAPF (100 ± 5.5%, n ¼ 8; Fig. 8a,g), but no detectable staining in the antennal commissure (47.0 ± 1.7%, n ¼ 8; Fig. 8d,g). We then stained for the Wnt5 protein in the drl2 loss-of-function mutant. Of note, Wnt5 was found in the antennal lobe neuropil (96.0 ± 8.9%, n ¼ 16, P ¼ 0.5165 compared with wild-type neuropil; Fig. 8b,g) and the antennal 8 commissure, a structure enwrapped by processes of the TIFR glia (75.9 ± 8.5%, n ¼ 16, P o 0.0001 compared with wild-type commissure; dashed lines, Fig. 8e,g). This result indicates that drl downregulates Wnt5 protein in the antennal commissure. To determine whether the downregulation of Wnt5 at the midline is a result of drl acting in glia, we restored drl specifically to glia in the drl2 mutant. In drl2 mutants expressing UAS-drl under the control of the Repo-Gal4 driver, Wnt5 staining in the antennal commissure was no longer observed (43.7 ± 4.7%, n ¼ 16, P ¼ 0.25 compared with wild-type commissure; Fig. 8f,g). The midline accumulation of Wnt5 in the drl2 mutant prompted us to ask how it might affect glomerular development in the mutant. At 30 hAPF, as Wnt5 began to accumulate in the dorsomedial corners of the antennal lobes in the drl2 mutant (arrowheads in Fig. 8j, compare with Fig. 8h), projection neuron dendrites were observed lateral to the domains of Wnt5 staining, confined to the antennal lobes (Fig. 8j). At 36 hAPF, as high levels of Wnt5 accumulated in the commissure (arrowhead in Fig. 8k, compare with Fig. 8i), the projection neuron dendrites began to transgress the antennal lobe boundaries and projected medially toward the region of ectopic Wnt5 enrichment (arrows in Fig. 8k). Our results show that in the absence of drl, Wnt5 accumulates in the commissure, leading to the formation of ectopic midline glomeruli. DISCUSSION The mechanisms by which ingrowing axons sort into precise maps, such as those found in the olfactory glomeruli or the somatosensory barrels, are poorly understood. Deafferentation and transplantation experiments revealed that ingrowing axons are important for specifying the maps in the initially homogenous structures15,16,34. However, little is known about how the ingrowing axons carry out these feats. In this report, we show that ingrowing ORN axons express Wnt5, which contributes to organizing the glomerular pattern of the Drosophila ADVANCE ONLINE PUBLICATION NATURE NEUROSCIENCE © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience ARTICLES olfactory system. We also show that the Drl receptor tyrosine kinase acts in glial cells to modulate Wnt5 signaling. This previously unknown interaction between ORN axons and glia reveals an important function of ORN axon-glia interactions in regulating the precise neural circuitry of the Drosophila antennal lobes. The wnt5 mutant had characteristic disruptions of the olfactory map. Many dorsomedial glomeruli were displaced ventrally (resulting in heart-shaped antennal lobes) and the antennal commissure failed to form. In contrast to the loss-of-function defects, overexpression of wnt5 led to the displacement of glomeruli into the midline. Examination of the ORN axons in the wnt5 mutant showed that they take circuitous paths to their targets and frequently misprojected to dorsal regions of the brain. Consistent with a role for wnt5 in antennal lobe development, the antennal lobe defects appeared during the pupal stage, when ORN axon targeting and glomerular development occur. Our genetic mosaic and cell type–specific rescue experiments indicated that wnt5 is required in the ORNs. Antibody stainings indicated that the Wnt5 protein was enriched on the dendrites of the projection neurons, where it presumably accumulated subsequent to its secretion by ORNs. In addition to the projection neuron dendrites, Wnt5 also accumulated in the antennal commissure in the drl2 mutant. We propose that Wnt5 is a signal by which ingrowing ORN axons direct the development of their target field. Mutation of the drl gene also produced disruptions of the olfactory map. However, unlike the stereotyped shifts of glomeruli seen in the wnt5 mutant, the glomeruli were randomly positioned in or missing from one antennal lobe in the drl mutant. Furthermore, there was a strong tendency for glomeruli to form at the midline. As in the wnt5 mutant, ORN axons took indirect routes to their targets. That drl functions in development is supported by the observation that antennal lobe defects were visible at 40 hAPF, the time when ORN axon targeting and glomerular development take place. Antibody staining showed that the Drl protein was highly expressed by the projection neurons and TIFR glia, cells that are intimately associated with the ingrowing ORN axons. In the projection neurons, Drl was enriched in the dendrites of nascent glomeruli, four of which also appeared to accumulate Wnt5. The TIFR is a donut-shaped midsagittal structure located between the antennal lobes. Our histological studies showed that TIFR glial processes were closely associated with ORN axons that were projecting across the midline. Several observations indicated that drl functions in the TIFR to regulate wnt5 function. First, removal of drl from single projection neuron clones did not disrupt the development and morphology of the projection neurons. Second, neuronal expression of drl in the drl2 mutant background did not rescue the mutant phenotype. Third, expression of UAS-drl under the control of Repo-Gal4 strongly rescued the drl mutant phenotype, suggesting that drl functions in glial cells. Although we cannot rule out roles for Drl in the projection neurons, collectively, our observations suggest that drl functions predominantly in glial cells to regulate antennal lobe development. The phenotypic similarities between the drl loss-of-function and the wnt5-overexpressing mutants raise the intriguing possibility that the two genes act antagonistically in antennal lobe development. Indeed, expression of a weak wnt5 transgene in the ORNs, which has no effect in the wild type, triggers the formation of ectopic glomeruli in the drl2 mutant. Thus, wnt5 and drl function in opposition to each other in antennal lobe development. To ascertain the relative positions of wnt5 and drl in this signaling pathway, we generated animals carrying null mutations in both genes. We found that the wnt5400;drl2 double mutants had the characteristic wnt5 phenotype. The wnt5 gene is therefore epistatic to the drl gene, indicating that wnt5 functions NATURE NEUROSCIENCE ADVANCE ONLINE PUBLICATION downstream of drl in antennal lobe development. This conclusion is also supported by the observation that, although the removal of a copy of the wnt5 gene strongly suppressed the drl homozygous mutant phenotype, the removal of a copy of the drl gene had no effect on the wnt5 homozygous mutant phenotype. The genetic data that drl downregulates wnt5 function is further supported by our observation that the Wnt5 protein significantly accumulates in the commissure in the absence of Drl. Taken together, our genetic and histological data indicate that drl acts to inhibit the activity of wnt5 during antennal lobe development. To probe the molecular mechanisms by which Drl regulates antennal lobe development, we mutated the various domains of Drl. We observed that neither disruption of the kinase activity nor deletion of the intracellular domain significantly impaired rescue by the drl transgene. In contrast, deletion of the extracellular WIF domain completely abolished Drl’s ability to rescue the mutant phenotype. These results suggest that Drl regulates antennal lobe patterning predominantly through its extracellular WIF domain. How might Drl inhibit the function of Wnt5? One possibility is that Drl inhibits Wnt5 function simply by promoting Wnt5’s sequestration or endocytosis, thus limiting its interaction with another as yet unidentified receptor (Supplementary Fig. 5 online). This receptor might be one of the other Drosophila receptor tyrosine kinases or a member of the Frizzled family, one of which, frizzled 2 (fz2), interacts genetically with wnt5 to stabilize axons of the Drosophila visual system22. Alternatively, Drl may directly interact with another receptor and Wnt5, as has been observed previously for its mammalian ortholog Ryk and members of the Wnt and Frizzled families35. This interaction could inhibit or alter the signal transduced from the membrane. However, we did not observe a requirement for Drl’s cytoplasmic domain, suggesting that transduction of the Wnt5 signal by Drl alone is unlikely to have a major role in patterning the antennal lobes. How do glial cells interact with the ORN axons to specify the olfactory map? Our data suggest that the ingrowing ORN axons contribute to antennal lobe patterning through secretion of Wnt5 and that glial cells locally regulate Wnt5 actions through Drl. We propose the following working model for how Wnt5-Drl signaling might regulate glomerular patterning. Ingrowing ORN axons express Wnt5, which is important for the precise organization of the glomeruli and pathfinding of the ORN axons, such as those crossing the midline or projecting to the dorsomedial region of the antennal lobes. Normal antennal lobe development requires that the Wnt5 signal be locally attenuated by the TIFR glial cell–expressed Drl protein. In the wnt5 mutant, the lack of Wnt5 signaling results in the failure of ORN axons to cross the midline and the establishment of glomeruli in more ventral positions. In the drl mutant, Wnt5 accumulates at the midline and presumably inappropriately signals through another receptor, resulting in aberrant ORN axon targeting to the midline and the formation of ectopic glomeruli at the dorsomedial corner of the antennal lobe and at the midline. Further studies will hopefully help to unravel the precise mechanisms by which Wnt5 and Drl act together to specify the patterning of the Drosophila olfactory map. METHODS Experimental animals. The P{GS} Drosophila lines were generated by the Drosophila Gene Search Project (Metropolitan University, Tokyo)36. The Or-Gal4 and Or-MCD8-GFP (Or-mGFP) lines were from L. Vosshall (The Rockefeller University) and B. Dickson (IMBA, Austria), the UAS-drlDcyto line was from J. Dura (Institut de Genetique Humaine)33 and the 72OK-Gal4 line was kindly provided by K. Ito (The University of Tokyo). The construction of 9 ARTICLES the GH-mGFP line has been described14. Other stocks, referenced throughout the text, were obtained from Bloomington Drosophila Stock Center. © 2007 Nature Publishing Group http://www.nature.com/natureneuroscience Transgenes. Full-length drl cDNA (from Open Biosystems) was subcloned into the pUAST vector to generate UAS-drl. To create UAS-drlK371A, we generated a fragment of the drl coding region (base pairs 1079–1830) bearing the K371A mutation by PCR, fused it in frame with the remainder of the drl coding region and then cloned it into pUAST. To generate UAS-drlDWIF, we generated a DNA fragment (nucleotide 472–1830) that lacked the WIF-encoding sequences by PCR, fused it with the first 60 nucleotides of the drl coding region, and then subcloned it into pUAST. Transgenic animals were generated by standard procedures. Immunohistochemistry. Adult (1 to 2 d old) or pupal brains were quickly dissected in cold phosphate-buffered saline (PBS, 130 mM NaCl, and 10 mM Na2HPO4, pH 7.2) and fixed in PLP fixative (2% paraformaldehyde, 0.25% sodium periodate, 75 mM lysine-HCl and 37 mM sodium phosphate, pH 7.4) for 1 h. The fixed brains were washed with PBST (PBS with 0.5% Triton X-100) and stained with primary antibodies overnight. For Wnt5 staining, dissected brains were directly stained with antibody to Wnt5 in PBS (2.5 h at 4 1C), washed with PBS and goat serum, and fixed in PLP (1 h, 25 1C). Antibodies and dilutions are described here. mAb nc82 (1:20 dilution) was a gift from A. Hofbauer37, rabbit antibody to GFP (1:100 dilution) was obtained from Molecular Probes, and rat antibody to mCD8 (1:100 dilution) was from Caltag. Affinity-purified rabbit antibody to Wnt5 (ref. 21) and rabbit antibody to Drl were both used at 1:100 dilutions. The anti-Drl antiserum was raised against a GST fusion protein that included Drl amino acids 123–222 and was affinity purified against the same protein coupled to a column. The secondary antibodies, FITC-conjugated goat antibodies to rabbit, Cy3-conjugated goat antibodies to mouse and FITC-conjugated goat antibodies to rat, were obtained from Jackson Laboratories and were used at 1:100 dilution. Statistic analysis. Statistical comparisons were performed using Microsoft Excel and GraphPad Prism4. Tests on independent groups were two-tailed Student’s t-test. Values and error bars indicate mean ± s.e.m. *, ** and *** denote P o 0.05, 0.01 and 0.001, respectively. Note: Supplementary information is available on the Nature Neuroscience website. ACKNOWLEDGMENTS We thank the Bloomington Drosophila Stock Center for the fly lines, A. Hofbauer for the generous gift of the nc82 antibody and W. Zhou for construction of the UAS-drl transgenic fly line. This work was supported by grants from the US National Institutes of Health and the US National Institute on Deafness and other Communication Disorders (DC5408-01), the Roy J. Carver Charitable Trust (#03-27) (H.H.), ASPASIA (Netherlands Organization for Scientific Research) and Pionier grants (J.N.), and a Genomics grant (L.F. and J.N.) from the Nederlandse Organisatie voor Wetenschappelijk Onderzoek. AUTHOR CONTRIBUTIONS Y.Y., Y.W., C.Y. and R.O. conducted experiments in the laboratory of H.H. R.R.W. conducted experiments in the laboratory of J.N.N. L.G.F., Y.Y., Y.W. and H.H. analyzed the data. T.A. provided the P{GS} lines screened by the H.H. lab. H.H. and L.G.F. wrote the manuscript with contributions from the other authors. Published online at http://www.nature.com/natureneuroscience Reprints and permissions information is available online at http://npg.nature.com/ reprintsandpermissions 1. Mombaerts, P. Axonal wiring in the mouse olfactory system. Annu. Rev. Cell Dev. Biol. 22, 713–737 (2006). 2. Dynes, J.L. & Ngai, J. Pathfinding of olfactory neuron axons to stereotyped glomerular targets revealed by dynamic imaging in living zebrafish embryos. Neuron 20, 1081–1091 (1998). 3. Potter, S.M. et al. Structure and emergence of specific olfactory glomeruli in the mouse. J. Neurosci. 21, 9713–9723 (2001). 4. Miyasaka, N. et al. Robo2 is required for establishment of a precise glomerular map in the zebrafish olfactory system. Development 132, 1283–1293 (2005). 10 5. Lin, D.M. & Ngai, J. Development of the vertebrate main olfactory system. Curr. Opin. Neurobiol. 9, 74–78 (1999). 6. Hummel, T. et al. Axonal targeting of olfactory receptor neurons in Drosophila is controlled by Dscam. Neuron 37, 221–231 (2003). 7. Hummel, T. & Zipursky, S.L. Afferent induction of olfactory glomeruli requires N-cadherin. Neuron 42, 77–88 (2004). 8. Key, B. & St John, J. Axon navigation in the mammalian primary olfactory pathway: where to next? Chem. Senses 27, 245–260 (2002). 9. Ang, L.H., Kim, J., Stepensky, V. & Hing, H. Dock and Pak regulate olfactory axon pathfinding in Drosophila. Development 130, 1307–1316 (2003). 10. Lattemann, M. et al. Semaphorin-1a controls receptor neuron-specific axonal convergence in the primary olfactory center of Drosophila. Neuron. 53, 169–184 (2007). 11. Sweeney, L.B. et al. Temporal target restriction of olfactory receptor neurons by Semaphorin-1a/PlexinA-mediated axon-axon interactions. Neuron. 53, 185–200 (2007). 12. Oland, L.A., Orr, G. & Tolbert, L.P. Construction of a protoglomerular template by olfactory axons initiates the formation of olfactory glomeruli in the insect brain. J. Neurosci. 10, 2096–2112 (1990). 13. Stout, R.P. & Graziadei, P.P. Influence of the olfactory placode on the development of the brain in Xenopus laevis (Daudin). I. Axonal growth and connections of the transplanted olfactory placode. Neuroscience 5, 2175–2186 (1980). 14. Ang, L.H. et al. Lim kinase regulates the development of olfactory and neuromuscular synapses. Dev. Biol. 293, 178–190 (2006). 15. Graziadei, P.P. & Kaplan, M.S. Regrowth of olfactory sensory axons into transplanted neural tissue. 1. Development of connections with the occipital cortex. Brain Res. 201, 39–44 (1980). 16. Schneiderman, A.M., Matsumoto, S.G. & Hildebrand, J.G. Trans-sexually grafted antennae influence development of sexually dimorphic neurones in moth brain. Nature 298, 844–846 (1982). 17. Oland, L.A. & Tolbert, L.P. Multiple factors shape development of olfactory glomeruli: insights from an insect model system. J. Neurobiol. 30, 92–109 (1996). 18. Moon, R.T., Bowerman, B., Boutros, M. & Perrimon, N. The promise and perils of Wnt signaling through beta-catenin. Science 296, 1644–1646 (2002). 19. Cadigan, K.M. & Nusse, R. Wnt signaling: a common theme in animal development. Genes Dev. 11, 3286–3305 (1997). 20. Yoshikawa, S., McKinnon, R.D., Kokel, M. & Thomas, J.B. Wnt-mediated axon guidance via the Drosophila Derailed receptor. Nature 422, 583–588 (2003). 21. Fradkin, L.G. et al. The Drosophila Wnt5 protein mediates selective axon fasciculation in the embryonic central nervous system. Dev. Biol. 272, 362–375 (2004). 22. Srahna, M. et al. A signaling network for patterning of neuronal connectivity in the Drosophila brain. PLoS Biol. 4, e438 (2006). 23. Zhang, D. et al. Misexpression screen for genes altering the olfactory map in Drosophila. Genesis 44, 189–201 (2006). 24. Jhaveri, D., Sen, A. & Rodrigues, V. Mechanisms underlying olfactory neuronal connectivity in Drosophila-the atonal lineage organizes the periphery while sensory neurons and glia pattern the olfactory lobe. Dev. Biol. 226, 73–87 (2000). 25. Fishilevich, E. & Vosshall, L.B. Genetic and functional subdivision of the Drosophila antennal lobe. Curr. Biol. 15, 1548–1553 (2005). 26. Couto, A., Alenius, M. & Dickson, B.J. Molecular, anatomical, and functional organization of the Drosophila olfactory system. Curr. Biol. 15, 1535–1547 (2005). 27. Lee, T. & Luo, L. Mosaic analysis with a repressible neurotechnique cell marker for studies of gene function in neuronal morphogenesis. Neuron 22, 451–461 (1999). 28. Dura, J.M., Taillebourg, E. & Preat, T. The Drosophila learning and memory gene linotte encodes a putative receptor tyrosine kinase homologous to the human RYK gene product. FEBS Lett. 370, 250–254 (1995). 29. Simon, A.F., Boquet, I., Synguelakis, M. & Preat, T. The Drosophila putative kinase linotte (derailed) prevents central brain axons from converging on a newly described interhemispheric ring. Mech. Dev. 76, 45–55 (1998). 30. Jefferis, G.S., Marin, E.C., Stocker, R.F. & Luo, L. Target neuron prespecification in the olfactory map of Drosophila. Nature 414, 204–208 (2001). 31. Stocker, R.F., Heimbeck, G., Gendre, N. & de Belle, J.S. Neuroblast ablation in Drosophila P[Gal4] lines reveals origins of olfactory interneurons. J. Neurobiol. 32, 443–456 (1997). 32. Hanks, S.K., Quinn, A.M. & Hunter, T. The protein kinase family: conserved features and deduced phylogeny of the catalytic domains. Science 241, 42–52 (1988). 33. Taillebourg, E., Moreau-Fauvarque, C., Delaval, K. & Dura, J.M. In vivo evidence for a regulatory role of the kinase activity of the linotte/derailed receptor tyrosine kinase, a Drosophila Ryk ortholog. Dev. Genes Evol. 215, 158–163 (2005). 34. Schlaggar, B.L. & O’Leary, D.D. Potential of visual cortex to develop an array of functional units unique to somatosensory cortex. Science 252, 1556–1560 (1991). 35. Lu, W., Yamamoto, V., Ortega, B. & Baltimore, D. Mammalian Ryk is a Wnt coreceptor required for stimulation of neurite outgrowth. Cell 119, 97–108 (2004). 36. Toba, G. et al. The gene search system. A method for efficient detection and rapid molecular identification of genes in Drosophila melanogaster. Genetics 151, 725–737 (1999). 37. Hofbauer, A. Eine Bibliothek monoklonaler Antikorper gegen das Gehirn von Drosophila melanogaster. (University of Wurzburg, Wurzburg, 1991). ADVANCE ONLINE PUBLICATION NATURE NEUROSCIENCE