Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Environmental enrichment wikipedia , lookup
Sensory cue wikipedia , lookup
Activity-dependent plasticity wikipedia , lookup
Convolutional neural network wikipedia , lookup
Neuroethology wikipedia , lookup
Neural oscillation wikipedia , lookup
Optogenetics wikipedia , lookup
Recurrent neural network wikipedia , lookup
Time perception wikipedia , lookup
Cortical cooling wikipedia , lookup
Visual search wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Visual selective attention in dementia wikipedia , lookup
Microneurography wikipedia , lookup
Visual memory wikipedia , lookup
Metastability in the brain wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Development of the nervous system wikipedia , lookup
Visual servoing wikipedia , lookup
Neural engineering wikipedia , lookup
Visual extinction wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Neuroesthetics wikipedia , lookup
C1 and P1 (neuroscience) wikipedia , lookup
/. Embryol. exp. Morph. 91, 101-115 (1986)
101
Printed in Great Britain © The Company of Biologists Limited 1986
Visual deprivation and the maturation of the
retinotectal projection in Xenopus laevis
M. J. KEATING, S. GRANT*, E. A. DAWES
AND K. NANCHAHALt
National Institute for Medical Research, Mill Hill, London NW71AA, UK
SUMMARY
There has been a resurgence of interest, recently, in the possible role of neural activity in the
ordering of synaptic connections in the lower vertebrate retinotectal system. Blockade of all
neural activity, by chronic administration of tetrodotoxin (TTX), during the regeneration of the
optic nerve in goldfish has been found to prevent the re-emergence of a fully ordered retinotectal
projection. We sought to determine the effects of visual deprivation, a less radical perturbation
of neural activity than that produced by TTX, on the initial development of the retinotectal
projection. The contralateral visuotectal projection was studied in Xenopus laevis which had
been reared in darkness from before the onset of visual function. The projection mapped
electrophysiologically at metamorphic climax, or in postmetamorphic juveniles, showed a
normal retinotopic topography. The topographic precision of the projection, as revealed by the
multiunit receptive field sizes, was the same in light- and dark-reared animals. The laminar
distribution, in the superficial neuropil of the optic tectum, of terminals from different classes of
retinal ganglion cells was also normal. It is concluded that the specific retinotectal connections
underlying these features of the projection are generated by intrinsic developmental processes
which do not require visual experience. Among these intrinsic processes might be 'spontaneous'
neural activity.
INTRODUCTION
In all vertebrate classes, optic fibres project to the contralateral optic tectum
(superior colliculus). This structure plays a major role in the localization of stimuli
in space and in the orientation of the organism to such stimuli (Trevarthen, 1968;
Schneider, 1969; Wurtz & Albano, 1980). A prerequisite for this function is the
orderly arrangement of optic fibre terminals in the tectum so as to produce a map
of the retinal surface, and hence of the visual field, across this structure. The lower
vertebrate retinotectal projection has been studied extensively with a view to
understanding the developmental mechanisms responsible for the production of
* Present address: Department of Anatomy, Medical College of Pennsylvania,
3200 Henry Avenue, Philadelphia, Penn. 19129, USA.
t Present address: Department of Environmental & Preventive Medicine, St Bartholomew's
Hospital Medical College, Charterhouse Square, London EC1M 6BQ, UK.
Key words: amphibia., visual deprivation, Xenopus laevis, retinotectal projection, neural
activity.
102
M. J. KEATING, S. GRANT, E. A. DAWES AND K. NANCHAHAL
ordered neuronal connections. This system has long been viewed as an archetype
of those systems in which ordered connections are produced by intrinsic developmental processes (Sperry, 1951, 1963; Gaze, 1970, 1984; Keating, 1976, 1981;
Jacobson, 1978; Rager, 1980). It has been assumed that neural activity or visual
experience had little role to play in the elaboration of the retinotectal projection.
Harris (1980) furnished some support for this view. He demonstrated, in the newt,
that an eye growing in an environment in which neural activity was suppressed by
tetrodotoxin (TTX) was nevertheless able to elaborate a retinotectal projection.
Within the limits imposed by neuroanatomical assessment of the projection, it was
found to display at least general retinotopic order.
The search for the mechanisms ordering the retinotectal projection has involved
both detailed studies of the normal developmental sequence through which the
mature projection is established (Gaze, Keating & Chung, 1974; Gaze, Keating,
Ostberg & Chung, 1979) and observations of the patterns of connections following
a wide range of developmental perturbations (see Edds, Gaze, Schneider & Irwin,
1979). Both groups of investigations have revealed a considerable potential
plasticity in retinotectal synaptic relationships. These results are difficult to accommodate within any simple variant of the theory of Neuronal Specificity which
attributes selective synaptogenesis to selective recognition between appropriately
labelled pre- and post-synaptic neuronal processes. The response to these difficulties has been the proposal of alternative mechanisms emphasizing the role of
fibre ordering (Horder & Martin, 1978; Rager, 1980) or the elaboration of
increasingly complex models, involving multifactorial contributions to the process
of synaptogenesis (e.g. Fraser & Hunt, 1980; Meyer, 1982a).
This environment has led to a reconsideration of the possible role of neural
activity in the detailed ordering of the retinotectal projection (Chung, 1974;
Willshaw & von der Malsburg, 1976; Whitelaw & Cowan, 1981). These theoretical
proposals have received recent experimental support. During regeneration of the
optic nerve in goldfish, neural activity in the nerve was blocked by chronic
intraocular administration of TTX (Meyer, 1983; Schmidt & Edwards, 1983). The
resulting retinotectal projection was less well ordered than in untreated controls.
Furthermore, in optic tecta caused to receive direct innervation from both eyes,
the binocular input was found to segregate into 'stripes' (Levine & Jacobson, 1975;
Constantine-Paton & Law, 1978; Law & Constantine-Paton, 1981; Springer &
Cohen, 1981). This segregation did not occur if neural activity in the optic nerve
was blocked by TTX (Meyer, 1982fc; Constantine-Paton & Reh, 1983; Boss &
Schmidt, 1984).
Given this resurgence of interest it seemed appropriate to examine the effects of
visual deprivation on the development of the retinotectal projection. Visual
deprivation will affect the pattern of neural activity in the optic nerve but will not
produce the total block of neural activity associated with TTX treatment. Xenopus
laevis larvae were placed in darkness at stage 35/36 (Nieuwkoop & Faber, 1967)
which is before the onset of visually driven electrical activity in the retina
(Witkovsky et al. 1976). They were reared in darkness until a terminal experiment
Visual deprivation and the retinotectal projection
103
in which the retinotectal projection was mapped electrophysiologically. Three
aspects of the retinotectal projection were assessed:
(i) detailed order in the projection as it is normally mapped;
(ii) the size of the multiunit receptive fields (MURFs) recordable at one tectal
locus - this is a quantitative index of the precision with which optic fibres
terminating at a given tectal site arise only from ganglion cells at a restricted,
retinal locus; and
(iii) the differential depth distribution of terminals from different classes of
retinal ganglion cells (Maturana, Lettvin, McCulloch & Pitts, 1959; Keating &
Gaze, 1970). A brief account of some of these results has appeared (Dawes,
Grant, Keating & Nanchahal, 1984).
METHODS
Xenopus laevis embryos were produced in the laboratory by mating adults following injection
of human chorionic gonadotrophin. Such embyros were divided into two groups. The control
group was raised under normal diurnal lighting conditions. The experimental group was raised,
from hatching at stage 35/36 (Nieuwkoop & Faber, 1967), in darkness. Both groups were
maintained in tanks containing oxygenated Stearns (+ iodide) solution. Until metamorphosis
they were fed three times/week on strained baby food (Heinz). The water was changed
once/month. After metamorphosis Tubifex worms were supplied once/week, at which time also
the water was changed.
Dark-rearing
The animals were reared in tanks in a light-tight cabinet in a light-tight room. This latter was
fitted with a light-tight double-entry lock. As a check on the upper limit of ambient illumination
within the light-tight cabinets, on several occasions photographic film was exposed within the
cabinet for a period of one week. On development, this film (Kodak Plus X Pan Professional
Sheet Film 4147) showed no evidence of light exposure. The sensitivity of this film is such that
this evidence indicates an. upper limit light energy level of 10~15 J cm"2 s"1. The actual light level
was probably considerably less than this. No visually driven activity is detectable electrophysiologically from an animal under comparable lighting conditions. Moreover, this regime was
sufficient to produce marked effects on another system of visual neuronal connections in
Xenopus, the intertectal system, the maturation of which does seem to require normal visual
experience (Keating & Feldman, 1975; Grant & Keating, 1981).
The visually deprived animals do experience a minimal degree of visual exposure during
feeding and water changing. Food and water were changed under conditions of very dim red
light and occupied about 30 s per week. The cumulative time of visual experience under such
limited conditions would, therefore, be about 5min for a stage-66 animal and 30min for a oneyear-old animal. Precautions to minimize visual experience were also taken during the preparation for terminal electrophysiological experiments. The animals were transported from the
animal accommodation to the laboratory in light-tight tanks, in which they were anaesthetized.
They were immediately fitted with thick light-occluding eye covers prior to dissection. One eye
remained covered during the acquisition of data on the retinotectal projection through the other
eye. No differences were noted between response properties early in the experiment and those
late in the experiment, that is after the eye had received several hours of visual experience.
Similarly, initial recordings through the eye that had been covered throughout the experiment,
showed response properties identical to those obtained through the eye that had been exposed
for several hours.
104
M. J. KEATING, S. GRANT, E. A. DAWES AND K. NANCHAHAL
Electrophysiological recordings
(i) Mapping the contralateral visuotectal projection
Contralateral visuotectal projections were mapped by standard electrophysiological techniques. In metamorphosing or three-month postmetamorphic juveniles the visuotectal projections were mapped with the animals immersed in Niu-Twitty solution in a transparent
perspex dome (Gaze et al. 1974). One-year postmetamorphic juveniles were mapped in air
(Gaze, Keating, Szekely & Beazley, 1970).
(ii) The depth distribution of unit types
Units recorded from optic fibre terminals in the superficial neuropil of the optic tectum were
classified according to their responses to moving black stimuli, to spots of light and to changes in
background illumination. 96 % of units were easily classifiable into one of three categories
(Keating & Gaze, 1970; Chung, Gaze & Stirling, 1973). 'Sustained' units show little response to
changes of background illumination, respond better to small stimuli than to larger ones, and
their characteristic feature is a sustained response to an object moved into their receptive field
and held stationary there. 'Event' units respond to both 'on' and 'off' of background illumination or light spots within the excitatory receptive field. They respond briskly to stimuli moving
across their receptive fields but give no sustained response to stationary stimuli. 'Dimmer' units
respond preferentially to large rather than small stimuli and respond to the switching 'off of a
light spot situated within their receptive field. The characteristic feature of these units is their
response to background illumination which consists of a burst of activity at each decrement in
background illumination, the response becoming more vigorous as darkness is approached.
Unclassifiable units were omitted from further analysis.
In normal anurans these different unit types assume a laminar distribution in the superficial
neuropil (layers 8 and 9) of the tectum (Maturana et al. 1959; Keating & Gaze, 1970; Chung et al.
1973; Chung, Bliss & Keating, 1974). One of our primary concerns in this paper was whether
visual experience was necessary to effect this laminar distribution. To determine this we would
ideally have liked to make absolute measures of the depths from the surface of the tectum at
which the different unit types are recorded. While this is feasible in Rana it is not in Xenopus.
The superficial neuropil of a one-year postmetamorphic Xenopus is only some 200-250 /an thick,
which is approximately half that of adult Rana. Moreover the pia mater is thicker in Xenopus
than in Rana, so problems of dimpling and drag during an electrode penetration are correspondingly greater in Xenopus. For these reasons the laminar distribution of unit types in normal
and in dark-reared Xenopus was assessed by noting the relative positions, within any penetration, of units of different type.
(iii) The plotting of multiunit receptive fields
A recording microelectrode at one locus in the superficial neuropil of the optic tectum may
record from only one or from several optic fibre terminals. In the latter case the recording is
referred to as a multiunit recording and the receptive field, stimulation of which produces
activity in the recording microelectrode, is a multiunit receptive field (MURF). The size of the
MURF is an index of the precision of spatial organization in the retinotectal projection. It
indicates the size of the retinal locus containing the ganglion cells whose fibre terminals project
to the vicinity of the microelectrode position.
The measured size of the MURF may be influenced by the selectivity and sensitivity of the
recording microelectrode, and by the level and signal-to-noise ratio of the amplification. We
attempted to minimize the possible effects of such variables. Microelectrodes were selected for
their capacity to record multiunits. When possible, and this was usually the case, the same
recording microelectrode was used for experiments on both light-reared and dark-reared
animals. The amplification levels of the signal were constant throughout the experimental series.
During multiunit recordings, data from the recording microelectrode were fed through a
window discriminator. The level of the window was set 100% above the noise level. For a
recording to be characterized as multiunit, we required that it be constituted of at least three
different spikes, above the window level, as revealed by differences in spike height and shape.
Visual deprivation and the retinotectal projection
105
Analysis in this paper is limited to multiunits of the 'event' type (or Class III of Maturana,
Lettvin, McCulloch & Pitts, 1960; Griisser, Griisser-Cornehls & Bullock, 1964). Our main
purpose was to compare the size of MURFs in normal and in dark-reared animals. The MURFs
of different unit types are themselves different in normal animals. Inclusion of different unit
types would, thus, have introduced an additional variable. 'Event' units are the most commonly
encountered.
The sizes of 'event' MURFs were plotted quantitatively using computer control of the visual
stimulus and analysis or the acquired spike data. Visual stimuli consisted of square dark stimuli
moved systematically, under computer control, across a large television screen. The approximate position of the MURF was first determined on an Aimark perimeter as in the procedure
by which the contralateral visuotectal projection is normally mapped. The perimeter was then
removed and the television screen positioned so that the MURF was approximately centred on
the screen. The screen was situated 38 cm from the animal's eye. At this distance the area of the
visual field occupied by the effective stimulus area of the screen was 64-5° in the nasotemporal
and 34-5° in the superior-inferior axis of the field. The stimulus routinely used was a dark square
of side 6° moved at a velocity of 44° s"1. The background luminance of the screen was 170 cd
m~2 and the contrast between the visual stimulus and this background was 0-98.
The screen was traversed systematically by the visual stimulus. Starting at one corner of the
screen the stimulus moved across the screen either vertically or horizontally. It was then stepped
1-5° on a direction orthogonal to the sweep, and the vertical or horizontal sweep repeated. This
process was repeated until the entire screen had been traversed by the moving stimulus. Spikes
occurring during the traverse were timed and recorded by the computer in a manner similar to
that described by Hodos, Dawes & Keating (1982). The computer produced a two-dimensional
matrix representation of the screen. This field was divided into l-5°xl-5° areas and the number
of spikes resulting from stimulation of that area was presented as an element in the 43x23
matrix. The measurement of a MURF size involved the pooling of matrices from traverses
involving stimulus movement in both horizontal and both vertical directions. The diameter of
the MURF was measured from this matrix output - the horizontal diameter from responses to
vertical stimulus movement and the vertical diameter from responses to horizontal movement.
The diameter recorded was the mean of these two diameters. To eliminate errors of spurious
readings due to occasional spontaneous or artefactual spikes, the excitatory receptive field size
of the MURF was considered to be that in which individual matrix elements were 10 % or more
of the maximum element size. It was the diameter of this field that was measured. The
computer, also, produced a graphical representation of this information. This involved a
pseudo-three-dimensional plot in which the x axis represented the nasotemporal dimension of
the field, the y axis the superoinferior dimension of the field and the z axis the number of spikes
evoked in response to stimulation of the corresponding l-5xl-5° of the visual field.
RESULTS
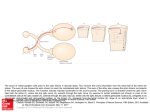
The contralateral visuotectal projection was mapped in 39 dark-reared animals.
9 of these animals were mapped at metamorphic climax, 7 at three months after
metamorphosis, 18 at one year postmetamorphosis, and 5 at three-five years
postmetamorphosis. The result from an animal mapped at metamorphic climax is
shown in Fig. 1, and that from a one-year-old juvenile in Fig. 2. Both projections
show detailed retinotopic ordering comparable to that seen in normal animals. No
disturbance of order in either the dorsoventral or nasotemporal retinal axis was
observed.
A more quantitative measure of the precision of connections in the contralateral
visuotectal projection is given by the size of the multiunit receptive field (MURF)
recordable from a single tectal locus. MURF sizes were measured for 29 event
multiunits from eight normal animals and for 35 event multiunits from six darkreared animals. All animals were aged one year postmetamorphosis. The mean
106
M. J. KEATING, S. GRANT, E. A. DAWES AND K. NANCHAHAL
/^
Left tectum
\
48
45 •
41 •
34 •
27 •
20 •
13 •
7
>
X
•"*•>
•
•
•
•
•
•
•
•
.3
46
42
•
•
•
•
•
• 4
• 1.
•
•
•
•
•
•
•
•
•
35
28
21
14
8
Right
visual
field
Fig. 1. Contralateral visuotectal projection through the right eye to the left tectum in a
stage-66 Xenopus raised in total darkness from stage 35/36. The outline of the dorsal
surface of the left tectum is shown above with the midline arrow pointing rostrally.
Below is shown the visual field of the animal's right eye. The animal may be imagined
as positioned behind the chart representation with the optic axis of its right eye centred
on the origin of the chart, looking out at the reader. The disc is the outline of a standard
perimetric chart representing the visualfieldextending out for 100° of visual angle in all
directions from the centre. The numbers on the tectal diagram represent microelectrode positions. The corresponding numbers in the visual field representation
indicate the localized visual field positions, stimulation of which evoked activity at the
corresponding microelectrode position. N, S, T, /-Nasal, superior, temporal, inferior
poles of the right visual field. Bar equals 200 jum.
Visual deprivation and the retinotectal projection
Left tectum
Right
visual
field
x
K i
\
\
Fig. 2. Contralateral visuotectal projection through the right eye to the left tectum in a
one-year postmetamorphic juvenile Xenopus raised in darkness from stage 35/36.
Conventions as in Fig. 1. Bar equals 200 fim.
107
108
M. J. K E A T I N G , S. G R A N T , E . A . D A W E S AND K.
NANCHAHAL
A
Fig. 3. Pseudo-three-dimensional plots of an 'event' multiunit receptive field in (A)
a normal one-year postmetamorphic Xenopus and in (B) a one-year postmetamorphic
Xenopus reared in darkness from stage 35/36. (Details in Materials and Methods.) The
horizontal axis represents 64-5° of the nasotemporal field axis and the vertical axis
represents 34-5° of the superoinferior field axis.
MURF diameters (±S.D.) for the two groups were normal animals 20-5° ± 5-8°,
dark-reared animals 21-1° ± 6-9°. The difference was not significant. These values
are different from those given in our preliminary communication (Dawes et al.
1984). For that communication we measured receptive field diameter using a
criterion of matrix elements containing 50 % or more of the maximum element
size, whereas here we present data using a 10% criterion (see Materials and
Methods). Illustrative plots of MURFs from a light-reared and a dark-reared
animal are shown in Fig. 3.
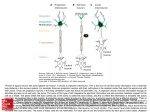
To study the relative depth distribution at which different unit types were
recorded, units were classified by standard criteria (see Materials and Methods)
into one of three classes - sustained, event or dimmer. The unit's depth position,
relative to that of other unit classes recorded in the same electrode penetration,
was noted. For this purpose a total of 89 penetrations were made infifteennormal
109
Visual deprivation and the retinotectal projection
animals and 69 penetrations in eight dark-reared animals. The animals were aged
one year postmetamorphosis. In 50 of these penetrations (28 in normal animals
and 22 in dark-reared animals) only one unit type was recorded during the
penetration. These were thus discarded from the analysis. The remaining 108
penetrations were categorized according to the number of unit types recorded and
the relative depths of those unit types. The data for normal and for dark-reared
animals are shown in Fig. 4. In 42 penetrations all three unit types were recorded.
30 of these penetrations were in normal and 12 in dark-reared animals. In all 42
penetrations the sustained units were always the most superficial, the dimmer units
the deepest, and event units were found at intermediate depths. In 66 penetrations
(31 in normal and 35 in dark-reared animals) only two of the three unit types were
recorded. In no case did the relative position, in depth, of these unit types deviate
from the normal. We conclude from these data that the normal laminar distribution of unit types is present in dark-reared animals.
DISCUSSION
The experiments reported in this paper sought to examine the hypothesis that
visually driven neural activity participates in the ordering of connections in the
30
11
15
16
19
5—
—A—,A—A-
D
—Q
-A—A—A-
_..
Fig. 4. Diagrammatic representation of the laminar distribution of different classes of
optic fibre terminals in the superficial neuropil of the optic tectum in (A) one-year
postmetamorphic normal Xenopus, and in (B) one-year postmetamorphic Xenopus
reared in darkness from stage 35/36. The diagram represents a transverse section
through the tectal neuropil, with the dorsal surface of the optic tectum uppermost.
Vertical lines represent electrode penetrations in which different combinations of two
or more of the three fibre classes were recorded. The number at the surface represents
the number of that particular penetration category that were recorded. 5, D, • sustained units; E, A, A - event units; D, O, • - dimmer units.
110
M. J. KEATING, S. GRANT, E. A. DAWES AND K. NANCHAHAL
developing contralateral retinotectal projection of Xenopus laevis. This is, we
believe, the first study involving mapping of the retinotectal projection in a lower
vertebrate visually deprived from before the onset of visual function. Our results
indicate that visually driven neural activity is not required for the maturational
elaboration of a highly ordered projection.
The effects of visual environment on the maturation of visual structures have
been investigated in some teleost fish (Grun, 1979; Jeserich & Rahmann, 1979;
Rahmann, Jeserich & Zeutsius, 1979; Zeutsius & Rahmann, 1984; Zeutsius,
Probst & Rahmann, 1984). Retinal development apparently proceeds normally
under conditions of visual deprivation but can be accelerated slightly by visual
experience (Grun, 1979). The initial development of optokinetic responses and of
tectal structure was not affected by visual deprivation. Prolonged dark rearing did
produce deficits in optokinetic behaviour and in tectal lamination and changes in
synaptic ultrastructure in the tectum. The topography of the retinotectal projection was not examined in these studies. Moreover, prolonged dark rearing
produced marked general effects on motility and growth. The fish did not mature
to adulthood and only a few survived 100 days of dark rearing. Given these
profound general effects, it is possible that the deficits in visuomotor behaviour
and tectal structure were not primary responses to visual deprivation. They may
have reflected secondary effects of the disruption of general mechanisms necessary
for growth and survival.
The effects of various forms of visual deprivation on the development of the
mammalian visual system have been the focus of many investigations (reviewed in
Wiesel, 1982; Sherman & Spear, 1982; Fregnac & Imbert, 1984; Boothe, Dobson
& Teller, 1985). Dark rearing or binocular lid suture produces some abnormalities
in the dorsal lateral geniculate nucleus and visual cortical areas. The topography of
the projections is not affected but receptive field properties of cortical cells are less
specific and less responsive than normal (Spear, Tong & Sawyer, 1983). Responses
in the superior colliculus, following developmental binocular deprivation, display
a marked reduction in directional selectivity and a reduced response to the
ipsilateral eye (Flandrin & Jeannerod, 1975; Hoffman & Sherman, 1975). It
appears, however, that these effects are secondary to defects in the visual cortex or
corticotectal pathway since similar effects are produced in normal cats following
visual cortical lesions (Sterling & Wickelgren, 1970; Mize & Murphy, 1976). The
actual retinotectal input itself does not appear to be affected by binocular visual
deprivation.
The maturation of the vertebrate retinotectal system thus seems to be less
dependent upon visual experience than does the mammalian geniculocortical
system. Visual deprivation alters, but does not eliminate, neural activity in visual
pathways. Total blockade of propagated sodium action potentials may be achieved
by administration of TTX, a sodium channel blocker. If neural activity has a role
to play in the maturation of visual pathways then such a role may be revealed by
observing the effects of TTX.
Visual deprivation and the retinotectalprojection
111
Harris (1980,1984) addressed this question in an ingenious series of experiments
on the developing amphibian retinotectal system. He transplanted an embryonic
eye cup from Ambystoma trigrinum to an embryonic newt of the species Taricha
torosa. This latter species manufactures TTX in sufficient quantities to suppress
action potentials in a TTX-sensitive species such as Ambystoma. The transplanted
eye from Ambystoma thus grew in an environment in which it was incapable of
generating action potentials. The transplant grew, apparently normally, and
elaborated a retinotectal projection. Of necessity the topography of this projection
had to be assessed neuroanatomically. Evidence for a reasonable degree of
retinotopicity was presented. The methodology forced by the paradigm would not
have revealed small differences in the organization of the projection.
On the other hand, in the mammalian visual system, Archer, Dubin & Stark
(1982) reported that the development of retinogeniculate connections was affected
much more profoundly by TTX blockade than by visual deprivation. The ipsilateral retinotectal projection in mammals during development displays an initial
coverage of the greater part of the superior colliculus, but then in early postnatal
life undergoes restriction to rostral colliculus (Land & Lund, 1979). This
restriction is associated with selective retinal ganglion cell death (Insausti,
Blakemore & Cowan, 1984). Fawcett, O'Leary & Cowan (1984) showed that TTX
blockade of neural activity prevented this cell death and the associated restriction
of the ipsilateral retinotectal projection.
A role for neural activity in the 'fine tuning' of connections in the lower
vertebrate retinotectal projection has been suggested by observations on the effect
of TTX blockade during the regeneration of optic fibres, following nerve section,
in adult goldfish. Meyer (1983) and Schmidt & Edwards (1983) reported that,
while the resulting retinotectal projection showed normal polarity and general
retinotopic order, the precision of retinotopicity was considerably reduced compared to that of normal or control regeneration maps. These findings suggest that
neural activity does play a role in the refinement of topographic order in the
retinotectal projection.
If one were to extrapolate these findings on the regenerating goldfish
retinotectal projection to the initial development of the system, and then compare
them with our findings that dark rearing does not affect the fine tuning in
the developing retinotectal projection, then it might be concluded that it is
'spontaneous', i.e. non-visually driven neural activity, that plays this refining role.
Willshaw & von der Malsburg (1976) presented a model of the organization of
specific connections between two neuronal arrays such as the retina and tectum.
This model assumed some initial polarity information in the arrays, but beyond
that the major organizing mechanism lay in a postulated coherence of spontaneous
activity in neighbouring ganglion cells. Such statistical coherence has been described in retinal ganglion cells of goldfish (Arnett, 1978), rabbit (Arnett &
Spraker, 1981) and cat (Mastronarde, 1983).
A test of this hypothesis might be provided by rearing under conditions of
stroboscopic illumination. Stroboscopic illumination would be expected to entrain
112
M. J. KEATING, S. GRANT, E. A. DAWES AND K. NANCHAHAL
the 'spontaneous' activity and render the firing of all ganglion cells coherent, thus
removing the neighbourhood connotation of such coherent firing. Schmidt &
Eisele (1985) report that, in goldfish, the retinotectal projection resulting from
regeneration of the optic nerve fails to show refined topography if the fish are
exposed only to stroboscopic illumination. Experiments are in progress to determine whether Xenopus, reared under stroboscopic conditions, also show disruption of their retinotectal projections.
A somewhat discordant note to the line of reasoning being developed here has,
however, been sounded by Schmidt & Eisele (1985). Three goldfish were maintained in darkness during regeneration of the optic nerve. Schmidt & Eisele found
that the resulting retinotectal projection showed reduced topographic precision in
that the MURF size was larger than normal. Schmidt (1985) takes the view that the
coherence of 'spontaneous' activity in neighbouring ganglion cells is alone insufficient to produce topographic precision in the projection. Refinement of topographic order requires additional coherence provided by local visual stimulation.
Further work will be required to determine whether the difference between our
results and those of Schmidt & Eisele (1985) reflects a species difference, or a
difference between development of the projection and regeneration. It is interesting that the visual discrimination behaviour of goldfish, following optic nerve
regeneration in darkness, is not different from that of fish maintained during
regeneration under normal diurnal lighting conditions (Leitner, Francis &
Gazzaniga, 1982). Moreover, Yoon (1975) reported that the retinotopically
ordered compression of the entire visual field onto rostral tectum, that follows
caudal tectal removal in goldfish, occurred even if the fish were maintained in
darkness.
Little analytical attention has been given to the mechanisms responsible for
what might be called a third dimension of the retinotectal projection. Different
retinal ganglion cell classes, revealed by different receptive field properties, send
their axons to different laminae in the superficial neuropil of the anuran optic
tectum (Maturana et al. 1959). This laminar distribution is restored following
regeneration of the optic nerve (Keating & Gaze, 1970). Chung et al. (1973) made
the intriguing observation that in adult Xenopus, maintained under prolonged
stroboscopic lighting conditions, this laminar distribution was disrupted. We find
no such disruption under conditions, of visual deprivation. The juxtaposition of
these two findings may indicate that 'spontaneous' activity plays a part in generating the laminar distribution. Tucker & Hollyfield (1977) describe quite marked
changes in the inner plexiform layer of dark-reared Xenopus. One might have
expected this to produce significant changes in the receptive field properties of
ganglion cells in dark-reared animals. In terms of the qualitative criteria used to
classify receptive field types we did not observe any changes but quantitative
studies may reveal functional correlates of the morphological changes described by
Tucker & Hollyfield.
The present study is intended as a contribution to a dissection of the mechanisms
generating ordered neuronal connections. Such mechanisms may be broadly
Visual deprivation and the retinotectal projection
113
classified into those independent of neural activity and those utilizing neural
activity. The latter class may be further subdivided into intrinsic mechanisms
utilizing 'spontaneous' neural activity and those requiring environmentally driven
neural activity. Our data seems to indicate that in the anuran retinotectal system
the normal highly ordered connections are generated without the participation of
visually driven neural activity.
REFERENCES
S. M., DUBIN, M. W. & STARK, L. A. (1982). Abnormal development of kitten retinogeniculate connectivity in the absence of action potentials. Science, N.Y. 217, 743-745.
ARNETT, D. W. (1975). Statistical dependence between neighbouring retinal ganglion cells in
goldfish. Expl Brain Res. 32, 49-53.
ARNETT, D. & SPRAKER, T. E. (1981). Cross-correlation analysis of the maintained discharge of
rabbit retinal ganglion cells. /. Physiol. 317, 29-47.
BOOTHE, R. G., DOBSON, V. & TELLER, D. Y. (1985). Postnatal development of vision in human
and non-human primates. A. Rev. Neurosci. 8, 495-545.
Boss, V. C. & SCHMIDT, J. T. (1984). Activity and the formation of ocular dominance patches in
dually innervated tectum of goldfish. /. Neurosci. 4, 2891-2905.
CHUNG, S.-H. (1974). In search of the rules for nerve connections. Cell 3, 201-205.
CHUNG, S.-H., BLISS, T. V. P. & KEATING, M. J. (1974). The synaptic organization of optic
afferents in the amphibian tectum. Proc. R. Soc. Lond. B 187, 449-459.
CHUNG, S.-H., GAZE, R. M. & STIRLING, R. V. (1973). Abnormal visual function in Xenopus
following stroboscopic illumination. Nature, New Biology 246, 186-189.
CONSTANTINE-PATON, M. &LAW, M.I. (1978). Eye-specific bands in the tecta of three-eyed frogs.
Science, N.Y. 202, 639-<541.
CONSTANTINE-PATON, M. & REH, T. (1983). Eye-specific stripes in the tectal lobes of three-eyed
frogs are dependent on neural activity. Soc. Neurosci. Abstr. 9, 760.
DAWES, E. A., GRANT, S., KEATING, M. J. & NANCHAHAL, K. (1984). The elaboration of the
contralateral visuotectal projection in the frog, Xenopus laevis, is unaffected by visual
deprivation. J. Physiol. 350, 62P.
EDDS, M. V., JR, GAZE, R. M., SCHNEIDER, G. E. & IRWIN, L. N. (1979). Specificity and plasticity
of retinotectal connections. Neurosci. Res. Prog. Bull. 17, 243-375.
FAWCETT, J. W., O'LEARY, D. M. & COWAN, W. M. (1984). Activity and the control of ganglion
cell death in the rat retina. Proc. natn. Acad. Sci. U.S.A. 81, 5589-5593.
FLANDRIN, J. M. & JEANNEROD, M. (1975). Superior colliculus, environmental influences on the
development of directional responses in the kitten. Brain Res. 89, 348-352.
FRASER, S. E. & HUNT, R. K. (1980). Retinotectal specificity: models and experiments in search
of a mapping function. A. Rev. Neurosci. 31, 319-352.
FREGNAC, Y. & IMBERT, M. (1984). Development of neuronal selectivity in primary visual cortex
of cat. Physiol. Revs 64, 325-434.
GAZE, R. M. (1970). The Formation of Nerve Connections. New York: Academic Press.
GAZE, R. M. (1984). Ordered nerve connections: pathways and maps. In Organizing Principles
of Neural Development (ed. S. C. Sharma), pp. 361-381. New York: Plenum Press.
GAZE, R. M., KEATING, M. J. & CHUNG, S.-H. (1974). The evolution of the retinotectal map
during development in Xenopus. Proc. R. Soc. Lond. B 185, 301-330.
GAZE, R. M., KEATING, M. J., OSTBERG, A. & CHUNG, S.-H. (1979). The relationship between
retinal and tectal growth in larval Xenopus: implications for the development of the retinotectal projection. /. Ernbryol. exp. Morph. 53, 103-143.
GAZE, R. M., KEATING, M. J., SZEKELY, G. & BEAZLEY, L. (1970). Binocular interaction in the
formation of specific intertectal neuronal connections. Proc. R. Soc. Lond. B 175, 107-147.
GRANT, S. & KEATING, M. J. (1981). The effect of visual deprivation on the maturation of
intertectal neuronal connections in Xenopus laevis. J. Physiol. 320, 19-20P.
GRUN, G. (1979). Light-induced acceleration of retina development in a mouth-brooding
teleost. /. exp. Zool. 208, 291-302.
ARCHER,
114
M. J. K E A T I N G , S. G R A N T , E . A. D A W E S AND K.
NANCHAHAL
O. J., GRUSSER-CORNEHLS, U. & BULLOCK, T. H. (1964). Functional organization of
receptive fields of movement sensitive neurones of the frog's retina. Pflug. Arch. 279, 88-94.
HARRIS, W. A. (1980). The effects of eliminating impulse activity on the development of the
retinotectal projection in salamanders. /. comp. Neur. 194, 303-317.
HARRIS, W. A. (1984). Axonal pathfinding in the absence of normal pathways and impulse
activity. /. Neurosci. 4, 1153-1162.
HODOS, W., DAWES, E. A. & KEATING, M. J. (1982). Properties of the receptive fields of frog
retinal ganglion cells as revealed by their response to moving stimuli. Neuroscience 7,
1533-1544.
HOFFMANN, K. P. & SHERMAN, S. M. (1975). Effects of early binocular visual deprivation on
visual input to cat superior colliculus. /. Neurophysiol. 38,1049-1059.
HORDER, T. J. & MARTIN, K. A. C. (1978). Morphogenetics as an alternative to chemospecificity
in the formation of nerve connections. Symp. Soc. exp. Biol. 32, 275-358.
INSAUSTI, R., BLAKEMORE, C. & COWAN, W. M. (1984). Ganglion cell death during development
of ipsilateral retino-collicular projection in golden hamster. Nature, Lond. 308, 362-365.
JACOBSON, M. (1978). Developmental Neurobiology, 2nd edn. New York: Plenum Press.
JESERICH, G. & RAHMANN, H. (1979). Effects of light deprivation onfinestructural changes in the
optic tectum of the rainbow trout (Salmo gaidneri, Rich.) during ontogenesis. Dev. Neurosci.
2, 19-24.
KEATING, M. J. (1976). The formation of visual neuronal connections: an appraisal of the present
status of the theory of 'Neuronal Specificity'. In Studies on the Development of Behaviour and
the Nervous System, vol. 3. Neural and Behavioural Specificity (ed. G. Gottlieb), pp. 59-110.
New York: Academic Press.
KEATING, M. J. (1981). The organization of the developing visual system. In Development in the
Nervous System (ed. D. Garrod & J. D. Feldman). Brit. Soc. devl Biol. Symposium 5,
pp. 35-60. Cambridge University Press.
KEATING, M. J. & FELDMAN, J. D. (1975). Visual deprivation and intertectal neuronal
connections in Xenopus laevis. Proc. R. Soc. Lond. B 191, 467-474.
KEATING, M. J. & GAZE, R. M. (1970). The depth distribution of visual units in the contralateral
optic tectum following regeneration of the optic nerve in the frog. Brain Res. 21, 197-206.
LAND, P. W. & LUND, R. D. (1979). Development of the rat's uncrossed retinotectal pathway
and its relation to plasticity studies. Science, NY. 205, 698-700.
LAW, M. & CONSTANTINE-PATON, M. (1981). Anatomy and physiology of experimentally
produced striped tecta. /. Neurosci. 1, 741-759.
LEITNER, D. S., FRANCIS, A. & GAZZANIGA, M. S. (1982). Optic nerve regeneration in goldfish
under light deprivation. Brain Res. Bull. 8, 105-107.
LEVINE, R. L. & JACOBSON, M. (1975). Discontinuous mapping of retina on to tectum innervated
by both eyes. Brain Res. 98, 172-176.
MATURANA, H. R., LETTVIN, J. Y., MCCULLOCH, W. S. & PITTS, W. H. (1959). Evidence that cut
optic nerve fibers in a frog regenerate to their proper places in the tectum. Science, N. Y. 130,
1709-1710.
MATURANA, H. R., LETTVIN, J. Y., MCCULLOCH, W. S. & PITTS, W. H. (1960). Anatomy and
physiology of vision in the frog, Rana pipiens. J. gen. Physiol. 43. Suppl. 129-175.
MASTRONARDE, D. N. (1983). Correlated firing of cat retinal ganglion cells. I. Spontaneously
active inputs to X and Y-cells. J. Neurophysiol. 49, 303-324.
MEYER, R. L. (1982O). Ordering of retinotectal connections: a multivariate operational analysis.
Curr. Top. devl Biol. 17, 101-145.
MEYER, R. L. (19826). Tetrodotoxin blocks the formation of ocular dominance columns in
goldfish. Science, N.Y. 218, 589-591.
MEYER, R. L. (1983). Tetrodotoxin inhibits the formation of refined retinotopography in
goldfish. Dev. Brain Res. 6, 293-298.
MIZE, R. R. & MURPHY, E. H. (1976). Alterations of receptive field properties of superior
colliculus cells produced by visual cortex ablation in infant and adult cats. /. comp. Neurol.
168, 393-424.
NIEUWKOOP, P. D. & FABER, J. (1967). A Normal Table of Xenopus laevis (Daudin), 2nd edn.
Amsterdam: North-Holland Publishing Co.
GRUSSER,
Visual deprivation and the retinotectal projection
115
G. H. (1980). Development of the retinotectal projection in the chicken. Adv. Anat.
Embryol. 93, 1-92.
RAHMANN, H., JESERICH, G. & ZEUTSIUS, I. (1979). Ontogeny of visual acuity of rainbow trout
under normal conditions and light deprivation. Behaviour 68, 315-322.
SCHMIDT, J. T. (1985). Selective stabilization of retinotectal synapses by an activity dependent
mechanism. Fed. Proc. 44, 2767-2772.
SCHMIDT, J. T. & EDWARDS, D. L. (1983). Activity sharpens the map during the regeneration of
the retinotectal projection in goldfish. Brain Res. 269, 29-39.
SCHMIDT, J. T. & EISELE, L. E. (1985). Stroboscopic illumination and dark-rearing block the
sharpening of the regenerated retinotectal map in goldfish. Neuroscience 14, 536-546.
SCHNEIDER, G. E. (1969). Two visual systems: brain mechanisms for localization and
discrimination are dissociated by tectal and cortical lesions. Science, N.Y. 163, 895-902.
SHERMAN, S. M. & SPEAR, P. D. (1982). Organization of visual pathways in normal and visually
deprived cats. Physiol. Revs 62, 738-855.
SPEAR, P. D., TONG, L. & SAWYER. C. (1983). Effects of binocular deprivation on responses of
cells in cat's lateral suprasylvian visual cortex. J. Neurophysiol. 49, 366-382.
SPERRY, R. W. (1951). Mechanisms of neural maturation. In Handbook of Experimental
Psychology (ed. S. S. Stevens), pp. 236-280. New York: J. Wiley & Son.
SPERRY, R. W. (1963). Chemoaffinity in the orderly growth of nerve fibre patterns and
connections. Proc. natn. Acad. Sci. U.S.A. 50, 703-710.
SPRINGER, A. D. & COHEN, S. M. (1981). Optic fiber segregation in goldfish with two eyes
innervating one tectal lobe. Brain Res. 255, 23-36.
STERLING, P. &WICKELGREN,B. G. (1970). Function of the projection from the visual cortex to the
superior colliculus. Brain Behav. Evol. 3, 210-218.
TREVARTHEN, C. B. (1968). Two mechanisms of vision in primates. Psychol. Forsch. 31,299-337.
TUCKER, G. S. & HOLLYFIELD, J. G. (1977). Modifications by light of synaptic density in the inner
plexiform layer of the toad, Xenopus laevis. Expl Neurol. 55,133-151.
WHITELAW, V. A. & COWAN, J. D. (1981). Specificity and plasticity of retinotectal connections:
a computational model. J. Neurosci. 1, 1369-1387.
WIESEL, T. N. (1982). Postnatal development of the visual cortex and the influence of
environment. Nature, Lond. 299, 583-591.
WILLSHAW, D. J. & VONDERMALSBURG, C. (1976). How patterned neural connections can be set up
by self organization. Proc. R. Soc. Lond. B 194, 431-445.
WITKOVSKY, P., GALLIN, E., HOLLYFIELD, J. G., RIPPS, H. & BRIDGES, C. D. B. (1976).
Photoreceptor thresholds and visual pigment levels in normal and vitamin-A deprived
Xenopus tadpoles. /. Neurophysiol. 39, 1272-1287.
WURTZ, R. H. & ALBANO, J. E. (1980). Visual motor function of the primate superior colliculus.
A. Rev. Neurosci. 3,189-226.
YOON, M. G. (1975). Effects of post-operative environments on reorganization of retinotectal
projection in goldfish. /. Physiol. 246, 673-694.
ZEUTSIUS, I. & RAHMANN, H. (1984). Influence or dark-rearing on the ontogenetic development
of Sarotherodon mossambicus (Cichlidae, Teleostei): I. Effects on body weight, body growth
pattern, swimming activity and visual acuity. ExplBiol. 43, 77-85.
ZEUTSIUS, I., PROBST, W. & RAHMANN, H. (1984). Influence of dark-rearing on the ontogenetic
development of Sarotherodon mossambicus (Cichlidae, Teleostei): II. Effects on allometrical
growth relations and differentiation of the optic tectum. Expl Biol. 43, 87-96.
RAGER,
{Accepted 9 September 1985)