Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Magnesium transporter wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cell nucleus wikipedia , lookup
Cytokinesis wikipedia , lookup
Endomembrane system wikipedia , lookup
Cellular differentiation wikipedia , lookup
Protein phosphorylation wikipedia , lookup
Signal transduction wikipedia , lookup
Protein moonlighting wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
Intrinsically disordered proteins wikipedia , lookup
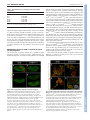
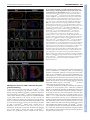
DEVELOPMENT AND STEM CELLS RESEARCH ARTICLE 2819 Development 137, 2819-2828 (2010) doi:10.1242/dev.052944 © 2010. Published by The Company of Biologists Ltd Arginine methylation of SmB is required for Drosophila germ cell development Joël Anne* SUMMARY Sm proteins constitute the common core of spliceosomal small nuclear ribonucleoproteins. Although Sm proteins are known to be methylated at specific arginine residues within the C-terminal arginine-glycine dipeptide (RG) repeats, the biological relevance of these modifications remains unknown. In this study, a tissue-specific function of arginine methylation of the SmB protein was identified in Drosophila. Analysis of the distribution of SmB during oogenesis revealed that this protein accumulates at the posterior pole of the oocyte, a cytoplasmic region containing the polar granules, which are necessary for the formation of primordial germ cells. The pole plasm localisation of SmB requires the methylation of arginine residues in its RG repeats by the Capsuléen-Valois methylosome complex. Functional studies showed that the methylation of these arginine residues is essential for distinct processes of the germline life cycle, including germ cell formation, migration and differentiation. In particular, the methylation of a subset of these arginine residues appears essential for the anchoring of the polar granules at the posterior cortex of the oocyte, whereas the methylation of another subset controls germ cell migration during embryogenesis. These results demonstrate a crucial role of arginine methylation in directing the subcellular localisation of SmB and that this modification contributes specifically to the establishment and development of germ cells. INTRODUCTION The small uridyl-rich nuclear ribonucleoproteins (U snRNPs) are the major constituents of the spliceosome, a multicomponent complex that catalyzes the splicing of pre-mRNAs in the nucleus. The majority of U snRNP particles consist of one U snRNA, several snRNA-specific proteins, and a common core of seven Sm proteins. The loading of the Sm proteins onto the snRNA is mediated by the survival of motor neurons (SMN) complex (Meister and Fischer 2002; Massenet et al., 2002). The 7methylguanosine cap of the snRNA is then hypermethylated to a 2,2,7-trimethylguanosine cap (Mattaj, 1986) and the snRNP is imported into the nucleus (Massenet et al., 2002; Narayanan et al., 2004; Ospina et al., 2005). Sm proteins are primarily small proteins that assemble to form characteristic homomeric or heteromeric rings that consist of six or seven subunits with a characteristic central pore, the presence of which is crucial for binding U-rich regions of single-stranded RNA (Scofield and Lynch, 2008). Members of the family are characterised by a conserved bipartite Sm domain or ‘Sm fold’, which functions, at least in part, in binding to neighbouring Sm proteins within such rings (Hermann et al., 1995; Séraphin, 1995; Khusial et al., 2005). SmB, SmD1 and SmD3 contain in their Cterminal tails glycine-arginine-glycine (GRG) and arginine-glycine (RG) motifs, the arginine residues of which are post-translationally methylated, a modification that takes place in the cytoplasm (Brahms et al., 2000). Department of Developmental Genetics, Deutsches Krebsforschungszentrum, Im Neuenheimer Feld 280, D-69120 Heidelberg, Germany. *Author for correspondence ([email protected]) Accepted 9 June 2010 In human cells, symmetrical dimethylarginine (sDMA) modification of Sm proteins is carried out by two distinct type II protein arginine methyltransferases (PRMTs): PRMT5 and PRMT7 (Gonsalvez et al., 2007). In agreement with inhibitor-based results showing that the recruitment of Sm proteins to the SMN complex is facilitated by sDMA modification (Brahms et al., 2001; Friesen et al., 2001a), depletion of either enzyme interferes with the ability of SMN to efficiently recognise the Sm proteins (Gonsalvez et al., 2007). Thus, sDMA modification of Sm proteins serves an important regulatory function in human cells. Loss of capsuléen (csul; the fly PRMT5 orthologue) activity results in hypomethylation of Sm proteins (Anne and Mechler, 2005; Gonsalvez et al., 2006; Anne et al., 2007). Despite a reduction in sDMA modification of Sm proteins, homozygous csul mutants are viable (Gonsalvez et al., 2006; Anne et al., 2007). In csul animals, the major Sm-class snRNPs are expressed normally (Gonsalvez et al., 2006), suggesting that csul is dispensable for snRNP assembly in Drosophila. In addition, flies express an orthologue of PRMT7, Dart7 (Art7, CG9882) (Boulanger et al., 2004), which is also required for sDMA modification of Sm proteins in vivo (Gonsalvez et al., 2008). Unlike csul, Dart7 is an essential gene (Gonsalvez et al., 2008). Attempts have been made to elucidate the function of the arginine methylation of Sm proteins. In human cultured cells the RG-rich tail of the SmD3 protein is required for snRNP assembly and optimal nuclear import (Friesen et al., 2001b; Girard et al., 2004; Narayanan et al., 2004). Moreover, a lack of arginine methylation at the C-terminus of SmD3 exerts no effect on snRNP assembly in the cytoplasm (Khusial et al., 2005). The requirement for methylation in snRNP assembly has been recently tested in Drosophila. The use of two distinct lines that synthesise modified isoforms of SmD1, in which either the entire RG-rich tail is deleted or the arginine residues are substituted by lysine, shows that in Drosophila the RG-rich tail of SmD1 is required for cell viability, whereas the symmetric dimethylation of C-terminal arginine DEVELOPMENT KEY WORDS: Drosophila, Arginine methylation, Sm proteins, Germ cells, Oogenesis residues in SmD1 is required neither for viability nor for snRNP assembly (Gonsalvez et al., 2008). These data suggest that the sDMA modification of SmD1 is dispensable for snRNP particle assembly. The role, if any, of the post-translational methylation of Sm proteins therefore remains unresolved. The goal of the present work was to determine the functional relevance of arginine methylation of SmB during Drosophila development. MATERIALS AND METHODS Molecular biology Plasmid constructs were generated by PCR amplification of the relevant DNA segments (High Fidelity PRC Master, Roche), which were subcloned into the p-{Pvas-GFP} (Sano et al., 2002) or p-{UASp} (Rorth, 1998) vectors. The SmD3 cDNA plasmid was kindly provided by H. Schenkel (DKFZ, Heidelberg, Germany). SmB and SmD1 cDNAs (BDGP EST Project clones LD14049 and RE39488, respectively) were provided by the Drosophila Genomics Resource Center (DGRC), Bloomington, IN, USA. SmB constructs and mutagenesis A combination of site-directed mutagenesis using the QuikChange Kit (Stratagene) and PCR amplification using mutagenic oligonucleotides was used to substitute arginine residues in the SmB coding sequence. Arginine residues R1-7 (at positions 112, 130, 1145, 174, 181, 189 and 196 of the SmB sequence) were modified to leucine (R1 and R3-6) or glycine (R2 and R7). Immunocytochemistry For whole-mount immunostaining, the following antibodies were used: antiOsk from rabbit and rat (gift of A. Ephrussi, EMBL, Heidelberg, Germany), anti-Vas from rat (gift of P. Lasko, McGill University, Montréal, Québec, Canada), anti-Tud from rabbit [TUD56; gift of S. Kobayashi (Amikura et al., 2001)], anti-symmetrical dimethylarginine (SYM10) from rabbit (Upstate), and anti-human SmB/B⬘ from mouse (ANA125, Calbiochem; gift of R. Bordonné, Université de Montpellier, France). DNA was counterstained with DAPI (Molecular Probes). Immunoreactivity was detected with appropriate Cy3-, Cy5- (Jackson ImmunoResearch) or Alexa Fluor 488- (Molecular Probes) conjugated secondary antibodies (1:200). Actin was detected using Alexa Fluor-phalloidin (Molecular Probes) at 1:300. Images were acquired on a Nikon Eclipse C1si laser-scanning confocal microscope and processed with Adobe Photoshop and ImageJ software. RESULTS Distribution of SmB during oogenesis Previous studies (Gonsalvez et al., 2006; Anne et al., 2007) have shown that the SmB protein is an in vivo substrate of the Capsuléen-Valois (Csul-Vls) methylosome complex. To monitor the distribution of SmB during oogenesis, the SmB coding sequence was cloned in frame with that of GFP under the control of the vasa (vas) germline-specific promoter (Sano et al., 2002) and a series of transgenic lines was generated; all tested lines produced similar amounts of GFP-SmB proteins in the ovaries (data not shown). As the methylated form of SmB, but not the unmethylated form, can interact in vitro with the germline-specific protein Tudor (Tud) (Anne et al., 2007), the distribution of GFP-SmB was first compared with that of Tud. In early stages of egg chamber development, GFP-SmB predominantly accumulated in the nuclei of nurse cells and oocyte (Fig. 1A-D) [see Spradling (Spradling, 1993) for a detailed description of egg chamber development]. From stage 9 onward, GFP-SmB was detected at the posterior pole of the oocyte (Fig. 1D and see Fig. S1 in the supplementary material). As described by Bardsley et al. (Bardsley et al., 1993), in early egg chambers Tud was found to be enriched in the nuage surrounding nurse cell nuclei and was present in the cytoplasm of the oocyte. At stage 7-8, Tud was localised temporarily at the anterior margin of the oocyte and from stage 9 became restricted Development 137 (17) to the pole plasm (Fig. 1A-D). Thus, before their common occurrence in the pole plasm, the two proteins displayed distinct distribution patterns. In particular, no GFP-SmB could be detected in the nuage or at the anterior cortex of the oocyte (Fig. 1C and 1D insets). In the oocyte nucleus of previtellogenic egg chambers, GFP-SmB was detected in a spherical structure attached to the karyosome (Fig. 1E), which may correspond to the Cajal body (or endobody) (Liu et al., 2006). Localisation of Sm proteins in the pole plasm Next, whether other Sm molecules could be detected in the pole plasm was investigated. As with SmB, transgenic lines producing GFP proteins fused to SmD1 or SmD3 were established. Both Sm proteins were selected because they contain sDMA residues in their C-terminal tails. No accumulation of GFP-SmD1 could be detected at the posterior pole of the oocyte of transgenic females (Fig. 2A), but a significant amount of GFP-SmD3 was detected at this location (Fig. 2B). To check whether the localisation of the GFPSmB fusion protein was similar to that of endogenous SmB, ovaries were stained using a monoclonal antibody (ANA125) directed against the human SmB/B⬘ (SNRPB – Human Genome Nomenclature Database) proteins. Only one protein band of the expected molecular mass was detected on a western blot using this antibody, demonstrating its specificity for SmB (Fig. 2C). Immunostained egg chambers showed an accumulation of endogenous SmB at the posterior pole of the oocyte, where it colocalised with the polar granule organiser Oskar (Osk) (Fig. 2D). The ANA125 antibody also detected the endogenous SmB protein at the posterior pole of the early embryos (data not shown). These data indicate that only a subset of the Sm proteins, such as SmB and SmD3, localises in pole plasm. Genetic requirement for SmB localisation in the pole plasm To examine the mechanism by which SmB becomes targeted to the pole plasm, the distribution of GFP-SmB in distinct genetic backgrounds was analysed. As osk plays a cardinal role in pole plasm formation (Ephrussi and Lehmann, 1992), the localisation of GFP-SmB was first investigated in osk oocytes. GFP-SmB was found to be absent from the posterior pole in oocytes of osk stage 10 egg chambers (Fig. 3A). The osk gene encodes two protein isoforms that are differentially distributed in the oocyte. The short form, which is active in pole plasm assembly, is a core component of the polar granules (Breitwieser et al., 1996), whereas the long form, which is required for their cortical anchoring, is associated with endocytic membranes along the posterior cortex (Vanzo et al., 2007). The localisation of GFP-SmB in egg chambers overexpressing one of the Osk isoforms was then investigated. The short and long Osk isoforms are carried by the p-{UAS-osk-M1RK10} and p-{UAS-osk-M139L-K10} transgenes, in which the 3⬘ UTR of the osk mRNA has been replaced by that of K10 and the initiation codon of the long Osk isoform substituted by a codon encoding an arginine or a leucine residue, respectively (Markussen et al., 1995). These transgenes, in combination with a p-{nos-Gal4} germline driver, directed the synthesis of high levels of either Osk isoform in both nurse cells and oocyte (Vanzo and Ephrussi, 2002). Examination of p-{GFP-Sm-B}; p-{UAS-osk-M1R-K10} /+; p-{nosGal4} /+ ovaries showed co-localisation of GFP-SmB with short Osk at the anterior margin of the oocyte (Fig. 3B, arrowheads), indicating that the short form of Osk could recruit GFP-SmB and consequently that SmB is a component of the polar granules. Analysis of p-{GFP-Sm-B}; p-{UAS-osk-M139L-K10} /+; p-{nos- DEVELOPMENT 2820 RESEARCH ARTICLE SmB methylation and germline development RESEARCH ARTICLE 2821 Gal4} /+ ovaries indicated that, in contrast to long Osk, which can be detected along the oocyte cortex, GFP-SmB is not observed at this location (see Fig. S2 in the supplementary material), suggesting that the long form of Osk is unable to recruit GFP-SmB. GFP-SmB distribution was also examined in two vas alleles. The vasO11 allele produces a Vas protein that is deficient in helicase activity (Liang et al., 1994), whereas vasQ7 synthesises no detectable full-length protein (Tomancak et al., 1998). In vasO11 stage 10 egg chambers, GFP-SmB was distributed normally at the posterior pole of the oocyte (Fig. 3C), suggesting that the accumulation of GFP-SmB in the pole plasm is independent of Vas helicase activity. By contrast, no GFP-SmB staining could be detected at the posterior pole of vasQ7 oocytes (Fig. 3D, upper panels), but a visible signal was associated with numerous punctate particles distributed throughout the cytoplasm in both nurse cells and oocyte of vasQ7 egg chambers (Fig. 3D, lower panels). In addition, the intensity of the GFP-SmB signal was markedly reduced in the Cajal body of the vasQ7 oocyte nucleus (Fig. 3D, lower-right panel) by comparison with that observed in vasO11. However, in vasO11 oocytes, the nuclear GFP-SmB-containing structure was frequently found to be dissociated from the karyosome or split into two particles (Fig. 3C, lower panels). These data indicate that Vas is needed for SmB localisation in the Cajal body and that the helicase activity of Vas is not required for SmB localisation in pole plasm. In tud egg chambers, GFP-SmB localisation at the posterior pole of the oocyte was abolished (Fig. 3E), indicating that Tud also plays an essential role in the accumulation of SmB in the pole plasm. Csul/Vls-dependent localisation of Sm proteins in the pole plasm To check whether arginine methylation is required for the localisation of SmB in the pole plasm, the distribution of GFPSmB in vls and csul mutant egg chambers was examined. Both genes encode crucial components of the methylosome and are required for the assembly of the pole plasm (Anne and Mechler, 2005; Gonsalvez et al., 2006; Anne et al., 2007). First, it was determined whether the GFP-SmB protein is a methylation target, similar to the endogenous SmB protein. For this purpose, western blots of ovarian protein extracts from wild-type and csul or vls mutant females carrying the p-{GFP-SmB} transgene were examined using SYM10 antibodies, which specifically recognise proteins containing multiple sDMA-glycine repeats (Boisvert et al., 2002). As shown in Fig. 4A, these antibodies predominantly reacted with a protein band of equivalent molecular mass to the endogenous SmB protein. In addition, they reacted with a band of ~50 kDa, which corresponds to the transgenic GFP-SmB fusion protein in the extract derived from p{GFP-SmB} flies. In ovarian protein extracts of p-{GFP-SmB}; csulRM and p-{GFP-SmB}; vls3 flies, the intensity of these two bands was strongly reduced, if not barely detectable. These results indicate that the methylation of GFP-SmB recapitulates that of endogenous SmB and confirm that the GFP-SmB protein is a methylation target of the Csul-Vls methylosome. The distribution of GFP-SmB in csul and vls mutant egg chambers was then analysed. In previtellogenic egg chambers, GFP-SmB was similarly distributed in wild-type and mutant nurse cell nuclei as well as in the Cajal body of the oocyte nucleus (Fig. DEVELOPMENT Fig. 1. Distribution of GFP-SmB during Drosophila oogenesis. The egg chamber is made up of a germline cyst surrounded by a monolayer of somatic follicle cells. Owing to incomplete cytokinesis, the cystocysts remain connected to each other through specialised cytoplasmic bridges. The oocyte is specified from one of these germline cells, whereas the remaining 15 cells differentiate into nurse cells. The oocyte, which is the posteriormost cell in the cyst, increases in size dramatically as oogenesis proceeds. In all panels, egg chambers are oriented with the posterior to the right. (A-D)Distribution of GFP-SmB and Tud in stage 4 (A), 6 (B), 8 (C) and 10 (D) egg chambers. GFP-SmB accumulates in nuclei of nurse cells and oocyte throughout egg chamber development. In stage 10 egg chambers, GFP-SmB localises at the posterior pole of the oocyte. Note that GFPSmB only co-localises with Tud in pole plasm (arrowhead). The insets show a single nurse cell nucleus, taken at a different focal plane to those shown in the whole stage 10 egg chamber, illustrating the absence of GFP-SmB accumulation in the nuage. (E)In previtellogenic egg chambers, GFP-SmB is detected in a dot in the oocyte nucleus that corresponds to the Cajal body (green in the overlay). This structure is attached to the karyosome, which is stained by the DNA-specific dye DAPI (red in the overlay). Fig. 2. A significant amount of GFP-SmD3, but not GFP-SmD1, accumulates in the pole plasm. (A,B)No GFP-SmD1 could be detected at the posterior pole of the oocyte in stage 10 egg chambers (A), whereas enrichment of GFP-SmD1 takes place in the pole plasm (B). Enlargements of the oocyte posterior region are provided on the right. (C)Anti-human SmB/B⬘ antibody (ANA125) recognises a single protein band with the expected molecular mass of SmB in Drosophila ovarian protein extracts. (D)Endogenous SmB (green in the overlay) colocalises with Osk (red in the overlay) in the pole plasm of stage 10 egg chambers. 4B-D). However, GFP-SmB was detected at the oocyte posterior pole in wild-type but not in mutant csul or vls stage 10 egg chambers (Fig. 4B⬘-D⬘). Whether SmD3 methylation by the Csul-Vls complex is also crucial for its pole plasm localisation was then examined. GFPSmD3 protein was found to be absent from the posterior pole of the csul or vls mutant oocyte (data not shown). These data thus demonstrate that the activity of the methylosome is required for the localisation of at least two Sm proteins, SmB and SmD3, in the pole plasm. Arginine methylation in RG repeats is essential for SmB localisation To extend the above results, the question of whether SmB localisation in the pole plasm is dependent on methylation of the arginine residues present in the RG repeats was addressed. The arginines located at positions 112, 130, 145, 174, 181, 189 and 196 in the SmB protein are present in RG dipeptide repeats and can be potentially methylated. These arginines are numbered 17 from the N-terminus to simplify mutant nomenclature. To investigate their role, a series of p-{GFP-SmB} transgenes was constructed, modified as follows: (1) deletion of the C-terminal tail from residue 108 (SmB1-107); (2) substitution of the seven arginines by leucine (L) or glycine (G) (SmBR1-7); (3) substitution of the three proximal arginines by L or G (SmBR1-3); and (4) substitution of the four distal arginines by L or G (SmBR4-7). All the modified sequences were inserted downstream of the vas promoter (Sano et al., 2002). Western blot analysis revealed that these transgenic flies produce similar amounts of GFP fusion proteins (Fig. 5A, upper panel). As shown in Fig. 5A (lower panel), SYM10 antibodies identified the endogenous SmB protein as well as the full-length unmodified GFP-SmB protein (1-199), but not the C-terminally deleted GFP-SmB1-107 Development 137 (17) Fig. 3. Genetic requirement for SmB localisation during oogenesis. (A)Pole plasm accumulation of GFP-SmB requires osk activity. GFP-SmB (green in the overlay) is detected at the posterior pole of the oocyte in wild-type (WT, top), but not in osk84/Df(3R)pXT103 (osk, bottom), egg chambers. (B)GFP-SmB is recruited by the short isoform of Osk. Overexpression of the short form of Osk during oogenesis leads to an accumulation of Osk (red in the overlay) and GFPSmB (green in the overlay) at the anterior margin of the oocyte (arrowheads) in stage 10 egg chambers. (C)In vasO11 stage 10 egg chambers, GFP-SmB can be detected in the pole plasm (top) and in the oocyte nucleus, either as a dot detached from the karyosome (bottom left) or as two dots (bottom right). (D)In vasQ7 stage 10 egg chambers, no GFP-SmB signal was observed at the posterior pole (top). However, GFP-SmB staining formed multiple dots dispersed in the nuclei and cytoplasm of nurse cells but not in the Cajal body (bottom). (E)Pole plasm accumulation of GFP-SmB requires tud activity. GFP-SmB remained undetected at the posterior pole of the tud1 oocyte. Higher magnifications of the posterior cortex of the oocyte are provided on the right (C-E). or the fully substituted GFP-SmBR1-7 proteins. For the partially substituted GFP-SmBR1-3 and GFP-SmBR4-7 proteins, the intensity of the signal was significantly decreased. The ability of the modified GFP-SmB proteins to accumulate in the pole plasm was then examined (Fig. 5B-F). In contrast to GFPSmB (1-199), which robustly accumulated in the pole plasm (Fig. 5B), the two methylation-deficient proteins (GFP-SmB1-107 and GFP-SmBR1-7) were unable to localise at the posterior pole of the oocyte (Fig. 5C,D). The two partially methylated proteins (GFPSmBR1-3 and GFP-SmBR4-7) displayed a moderate accumulation in the pole plasm (Fig. 5E,F). To further examine the role of arginine methylation in targeting SmB to the pole plasm another SmB mutant protein, in which the arginines of the tail were changed to lysine (GFP-SmBR1-7K), was expressed during oogenesis. The conservative arginine-to-lysine substitution retains the positive charge of the residue but removes its ability to be targeted by arginine methyltransferases. In agreement with the above results, DEVELOPMENT 2822 RESEARCH ARTICLE Fig. 4. Pole plasm accumulation of SmB requires methylosome function. (A)The level of GFP-SmB methylation is similar to that of endogenous SmB. Western blot of ovarian protein extracts from wildtype and transgenic females expressing GFP-SmB were probed with SYM10 antibodies (which detect symmetrical dimethylarginines). The GFP-SmB transgene was introduced into wild-type, csulRM and vls3 backgrounds. The intensity of the GFP-SmB and endogenous SmB SYM10-immunoreactive protein bands was strongly reduced in csulRM and vls3 protein extracts. The intensity of the other SYM10immunoreactive bands serves as an internal loading control. (B-D⬘) Distribution of GFP-SmB in wild-type (B,B⬘), csulRM (C,C⬘) and vls3 (D,D⬘) egg chambers. In stage 7 egg chambers (B-D), the GFP signal was detected with a similar intensity in the nuclei of wild-type and mutant germ cells (the oocyte nucleus is indicated by an arrow in B), whereas in mutant stage 10 oocytes (C⬘,D⬘) no GFP-SmB signal could be detected at the posterior pole. lysine substitution was found to abrogate the posterior pole localisation of SmB (Fig. 5H). These data demonstrate that the methylation of the arginine residues located in the RG repeats is essential for the localisation of SmB in the pole plasm. Arginine methylation of SmB in RG repeats is zygotically required for gonad formation Next, the function of SmB methylation during fly development was investigated. For this purpose, SmB, SmBR1-7 and SmBR1-7K cDNAs were cloned into a p-{UASp} transformation vector (Rorth, 1998) and transgenic strains were generated. The transgenes were then introduced into a mutant SmBBG02775/CyO heterozygous background and the flies were crossed with SmBl(2)SH0509/CyO flies carrying a ubiquitous tubulin-Gal4 driver. The SmBBG02775/SmBl(2)SH0509 combination (hereafter designated SmB for brevity) could be used to test whether expression of the SmB constructs restores viability. p-{UAS-SmB}, p-{UAS-SmBR1-7} and p-{UAS-SmBR1-7K} transgenes were found to be able to restore normal development of SmB animals, but the rescue efficiency was lower with the p-{UAS-SmBR1-7} and p-{UAS-SmBR1-7K} transgenes (~50% of the p-{UAS-SmB} transgene efficiency; Table 1). The transheterozygous combination was used because each SmBmutated chromosome 2 contained additional second-site lethal mutations (data not shown). These results suggest that methylation of the RG repeats of SmB is not crucial for fly viability. All rescued SmB flies with p-{UAS-SmBR1-7} or p-{UASSmBR1-7K} (collectively p-{UAS-SmBR1-7 or R1-7K}) were, however, found to be fully agametic (data not shown). To determine the developmental phase of defective gamete production, SmB; p{UAS-SmBR1-7 or R1-7K} embryos were collected and stained for Vas, a germ cell-specific marker (Hay et al., 1988). Identification of SmB; p-{UAS-SmBR1-7 or R1-7K} embryos was performed using RESEARCH ARTICLE 2823 Fig. 5. Pole plasm accumulation of SmB depends on the arginine residues in its RG-rich tail. (A,G)Western blots of ovarian protein extracts from transgenic females synthesising GFP-SmB fusion proteins, probed with anti-GFP antibodies. Anti-ribosomal p40 antibodies (p40) were used as loading control. Similar blots were probed with SYM10 antibodies. The white square in the lower panel of A indicates the position of the GFP-SmB1-107 band, as revealed by incubation of the same blot with anti-GFP antibodies (not shown). No SYM10 reactivity could be detected for GFP-SmB1-107, GFP-SmBR1-7 or GFP-SmBR1-7K, whereas GFP-SmBR1-3 and GFP-SmBR4-7 gave a reduced signal. (B-F,H) Distribution of the GFP-SmB proteins in stage 10 egg chambers. The size and structure of SmB and its derivatives are schematically depicted, with two Sm domains and the RG dipeptides shown as orange boxes and circles, respectively. Substituted arginine residues are shown as white (leucine or glycine) or red (lysine) circles. In contrast to GFP-SmB, which robustly localised at the posterior pole of the oocyte (B), GFP-SmB1-107, GFP-SmBR1-7 and GFP-SmBR1-7K proteins were absent from the pole plasm (C,D,H), and GFP-SmBR1-3 and GFP-SmBR4-7 proteins displayed a weak enrichment at this location (E,F). Higher magnifications of the posterior cortex of the oocyte are provided on the right. SYM10 antibodies. Control experiment showed that, by comparison with SYM10-positive wild-type and heterozygous embryos, the methylation-deficient SmB; p-{UAS-SmBR1-7 or R1-7K} embryos displayed only a weak signal (data not shown). In wild-type embryos, the precursors of the germ cells cross the primordial midgut epithelium and migrate bilaterally to reach the gonadal mesoderm on the dorsal surface of the midgut. Subsequently, germ cells associate, align and coalesce with the gonadal mesoderm to form the gonads [see Campos-Ortega and Hartenstein (Campos-Ortega and Hartenstein, 1985) for a detailed description of embryonic developmental stages] (Moore et al., 1998). Gonad formation was found to be abnormal in SmB; p-{UAS-SmBR1-7 or R1-7K} embryos. Although germ cells could DEVELOPMENT SmB methylation and germline development Development 137 (17) Methylation of R4-7 in SmB is required for germ cell development The involvement of arginine residues R1-7 of SmB in gonad formation prevents an analysis of their requirement for egg chamber development. To circumvent this problem, it was reasoned that decreasing the number of mutated arginines in SmB might produce a residual activity enabling gonad formation and egg chamber development. As the substitution of the arginines in the last four RG repeats of SmB was shown to reduce the efficiency of GFP-SmB targeting to the pole plasm, two transgenes, p-{UASSmBR4-7} and p-{UAS-SmBR4-7K}, were constructed and their in vivo activity analysed. Expression of either transgene driven by tubulin-Gal4 was able to rescue the zygotic lethality of SmB, albeit with a lower efficiency than a wild-type p-{UAS-SmB} transgene (Table 1). SmB; p-{UAS-SmBR4-7 or R4-7K} females developed ovaries but they were reduced in size (Fig. 7A⬘; data not shown). Ovary development in SmB; p-{UAS-SmB} females was similar to that of wild type, with ovarioles forming chains of developing egg chambers (Fig. 7B). By contrast, the ovaries of SmB; p-{UASSmBR4-7 or R4-7K} females contained predominantly previtellogenic egg chambers (Fig. 7A⬘; data not shown) and rarely egg chambers of later stages (see below). The majority of the egg chambers that reached the end of previtellogenic development showed signs of degeneration, including a condensed nuclear morphology (Fig. 7B⬘, upper panel; data not shown). To determine whether nuage formation could take place in early SmB; p-{UAS-SmBR4-7 or R4-7K} egg chambers, a Tud staining was performed. Tud was found to form a ring around the nurse cell nuclei in these egg chambers (Fig. 7B⬘, lower panel; Fig. 8G), suggesting that arginine methylation of SmB is not crucial for nuage assembly (see also Anne et al., 2007). These results indicate that a subset of arginine residues in the SmB tail is necessary for the growth of the germ cells and differentiation of the oocyte. Fig. 6. SmB is zygotically required for gonad formation. (A-B⬘) SmB; p-{UAS-SmB} (A,B) and SmB; p-{UAS-SmBR1-7} (A⬘,B⬘) embryos were collected and stained using Vas antibodies, which specifically label germ cells (red), and the DNA-specific dye DAPI (green). (A)In stage 10 SmB; p-{UAS-SmB} embryos, the germ cells migrated through the midgut epithelium, formed two bilateral clusters within the mesoderm, and aligned with the somatic gonadal precursors. (B)At stage 14, the two tissues coalesced to form the embryonic gonads. In SmB; p-{UAS-SmBR1-7} embryos, the germ cells were able to cross the epithelial wall of the midgut (A⬘), but were unable to coalesce with the gonadal mesoderm in later stages (B⬘). In SmB; p-{UAS-SmBR1-7}, the germ cells were scattered widely in the posterior half of the embryo. The upper and lower panels in B⬘ represent sagittal sections of the same embryo at two different focal planes. Fig. 7. SmB is maternally required for egg chamber development. (A-B⬘) Ovaries produced by SmB females rescued by p-{UAS-SmB} (A,B) or p-{UAS-SmBR4-7} (A⬘,B⬘) driven by p-{tubulin-Gal4} were dissected and stained for Tud (red) and for DNA with DAPI (green). By comparison with SmB; p-{UAS-SmB} ovaries, which displayed normally developing egg chambers (A), SmB; p-{UAS-SmBR4-7} ovaries were underdeveloped and contained only a reduced number of predominantly previtellogenic egg chambers (A⬘). A typical SmB; p{UAS-SmB} ovariole is made of developing egg chambers at different stages (B), whereas the largest egg chambers in SmB; p-{UAS-SmBR4-7} ovaries displayed signs of degeneration with pycnotic nurse cell nuclei (B⬘, top). Nuage formation, as indicated by the perinuclear accumulation of Tud protein, took place normally in SmB; p-{UASSmBR4-7} early egg chambers (B⬘, bottom). Table 1. Rescue efficiency of wild-type and mutant SmB transgenes Transgene WT R1-7 R4-7 R1-7K R1-3K R4-7K Rescue efficiency (%) 61 (n193) 32 (n238) 37 (n257) 28 (n174) 27 (n210) 30 (n171) For the wild-type (WT) and mutant SmB transgenes, the percentage reflects the ratio between the observed and expected number of homozygous flies. migrate through the midgut epithelium (Fig. 6A) they were unable to coalesce with the gonadal mesoderm. At stage 14 the Vaspositive germ cells were found to be dispersed in the posterior half of the SmB; p-{UAS-SmBR1-7 or R1-7K} embryos. Few germ cells, at most, were able to establish direct contact between them (Fig. 6B and see Fig. S3A in the supplementary material). These results show that SmB is zygotically required for the coalescence of germ cells with the gonadal mesoderm and indicate that the methylation of arginine residues in the tail of SmB is the key factor in this process. DEVELOPMENT 2824 RESEARCH ARTICLE SmB methylation and germline development RESEARCH ARTICLE 2825 Methylation of R4-7 in SmB is essential for polar granule anchoring As mentioned above, most of the SmB; p-{UAS-SmBR4-7} ovaries contain only previtellogenic egg chambers. However, a small fraction of the egg chambers was able to complete development in the rescued SmB; p-{UAS-SmBR4-7} females, which laid few eggs. These escapers allowed an analysis of the role of SmB methylation in pole plasm formation in the oocytes as well as in pole cell formation in the early embryos. By comparison with eggs laid by SmB; p-{UAS-SmB} females that formed blastoderm embryos with pole cells at their posterior pole (Fig. 8A), the embryos produced by SmB; p-{UAS-SmBR4-7} females built a normal layer of blastoderm cells but failed to form pole cells (Fig. 8A⬘). Furthermore, the amount of Vas protein localised at the posterior pole of early embryos derived from SmB; p-{UAS-SmBR4-7} females was significantly reduced compared with embryos produced by SmB; p-{UAS-SmB} females at similar stages (data not shown). Examination of vitellogenic SmB; p-{UAS-SmB} egg chambers revealed that, similarly to wild type, the Osk protein, which is only synthesised in the pole plasm (Markussen et al., 1995; Rongo et al., 1995), formed a dense and narrow crescent that was tightly anchored at the posterior pole of the oocyte (Fig. 8B,C). By contrast, Osk was found in a thicker structure that was loosely attached to the posterior cortex and in particles that were dispersed in the bulk ooplasm in SmB; p-{UAS-SmBR4-7 or R4-7K} oocytes at similar stages of development (Fig. 8B⬘,C⬘,F). A possible explanation of the failure to assemble Osk-containing granules at the oocyte posterior pole is a defective cytoskeleton network. To investigate this possibility, the distribution of the cortical matrix of F-actin in vitellogenic oocytes was analysed. In SmB; p{UAS-SmB} oocytes, the F-actin was aligned as a continuous layer underneath the oocyte cortex (Fig. 8C), whereas in SmB; p-{UASSmBR4-7 or R4-7K} oocytes the layer of F-actin appeared to be more diffuse at the posterior pole (Fig. 8C⬘,F). Examination of Tud, another pole plasm component, showed that its distribution bore the same relationship to that of F-actin as did Osk (Fig. 8D⬘). To assess whether Tud associates with Osk-containing particles, both proteins were simultaneously detected in SmB; p-{UAS-SmBR4-7} oocytes. Osk and Tud were found to co-localise in the released particles (Fig. 8E⬘). These data indicate that the methylation of arginine residues R4-7 of SmB is necessary for the anchoring of the polar granules at the posterior pole of the oocyte. DEVELOPMENT Fig. 8. Arginine methylation of SmB is maternally required for the anchorage of the polar granules at the posterior pole of the oocyte and for pole cell formation. Embryos or egg chambers of SmB mutant females expressing p-{UAS-SmB} (A-E), p-{UAS-SmBR4-7} (A’-E’) or p-{UAS-SmBR4-7K} (F,G) stained with the indicated antibodies and dyes. (A,A⬘) Pole cells formed at the posterior pole of blastoderm embryos laid by SmB; p-{UAS-SmB} females (A), whereas no pole cell could be detected in embryos laid by SmB; p-{UAS-SmBR4-7} females (A⬘). Germ cells were stained for Vas (red) and DNA with DAPI (green). (B,B⬘) In a stage 9 SmB; p-{UAS-SmB} egg chamber, Osk was tightly anchored at the posterior pole in a crescent-shaped pattern (B, red in the overlay), but was loosely associated with the oocyte posterior cortex in a stage 9 SmB; p-{UAS-SmBR4-7} egg chamber (B⬘). (C-F)Higher magnifications of the posterior cortex of SmB; p-{UAS-SmB} (C-E), SmB; p-{UAS-SmBR4-7} (C⬘-E⬘) and SmB; p-{UAS-SmBR4-7K} (F) stage 10 oocytes. Egg chambers were stained for Osk (red in the overlay of C,C⬘,F and green in the overlay of E,E⬘), Tud (D,D⬘,E,E⬘; red in the overlay), F-actin (C,C⬘,D,D⬘; green in the overlay) and DNA (blue in the overlay). In SmB; p-{UAS-SmB} oocytes, Osk and Tud particles formed a narrow crescent under the continuous cortical actin layer, whereas in SmB; p-{UAS-SmBR4-7} and SmB; p-{UAS-SmBR4-7K} oocytes the Osk and Tud particles were diffusely distributed in the cortical region and the actin organisation was severely disrupted at the posterior pole. Osk and Tud co-localised in the cortical polar granules in SmB; p-{UAS-SmB} (E) and in the loosely associated particles (arrowhead in E⬘ overlay) in SmB; p-{UAS-SmBR4-7} oocytes (E⬘). Note the correlation between the disorganised actin network and failure to maintain Osk particles at the cortex: the region indicated by the brackets in F shows that when the actin organisation seems unaffected, cortical localisation of Osk also appears normal, whereas both actin network and cortical accumulation of Osk are disrupted in the region outside the brackets. (G)In a stage 9 SmB; p-{UAS-SmBR4-7K} egg chamber, Tud was loosely associated with the oocyte posterior cortex but the nuage accumulation of Tud appeared unaffected. Methylation of R1-3 in SmB is zygotically required for gonad formation The above results also raise the question of whether the number or the location of methylated arginine residues in the SmB tail is functionally crucial. To address this point, a p-{UAS-SmBR1-3K} transgene was constructed and its in vivo activity analysed. Expression of the transgene driven by tubulin-Gal4 was able to rescue the zygotic lethality of SmB and with a similar efficiency to that of p-{UAS- SmBR1-7K} (Table 1). However, SmB; p-{UASSmBR1-3K} animals were found to produce no gametes (data not shown). The defect in gonad formation arose at the same developmental stages as observed with the p-{UAS- SmBR1-7K} transgene (see Fig. S3B in the supplementary material). The position, rather than the number, of methylated arginine residues thus appears to be crucial for SmB function during gametogenesis. As summarised in Fig. 9, the specific role of two subsets of arginine residues in the SmB tail can be defined: the R1-3 subset is required during embryogenesis for gonad formation, whereas the R4-7 subset is involved in pole cell formation by controlling polar granule maintenance at the posterior pole of the oocyte. DISCUSSION Arginine methylation-dependent localisation of Sm proteins in pole plasm Histochemical analysis indicates that the Sm proteins are predominantly present in the nucleus, but studies using cell fractionation techniques reveal that Sm proteins can be isolated as Sm rings in the cytoplasm. Here, it was determined that two Drosophila Sm proteins can specifically accumulate in the pole plasm during the final stage of oocyte differentiation until the formation of pole cells in blastoderm stage embryos. Inclusion of Sm proteins in cytoplasmic structures is a rather novel finding, although they have been detected outside of the nucleus in the germ cells of various species, including Caenorhabditis elegans (Barbee et al., 2002), Xenopus laevis (Bilinski et al., 2004), rat (Moussa et al., 1994; Toyooka et al., 2000) and mouse (Chuma et al., 2003). The present data show that access of SmB and SmD3 to the pole plasm depends on sDMA methylation. Although SmD1 contains arginine residues that are the targets of sDMA methylation, an accumulation of SmD1 in the pole plasm was not observed. This indicates that sDMA-modified Sm proteins, detected as GFP fusion proteins, are differentially targeted to the pole plasm. As sDMAmodified Sm proteins can interact with the Tud domains of SMN (Côté and Richard, 2005) and Tud has been previously shown to bind methylated SmB protein (Anne et al., 2007), it is possible to envisage that Tud confers specificity to the transport of SmB to the pole plasm. However, this hypothesis is nullified by the finding that SmB and Tud are conjointly detected only when both proteins have reached their destination at the posterior pole of the oocyte. Recent data indicate that the transport machinery responsible for the pole plasm localisation of SmB and SmD3 might correspond to a cytoplasmic osk mRNA RNP (Gonzalvez et al., 2010). SmB and SmD3 can form a stable association (Raker et al., 1996) but the unmethylated form of GFP-SmB was not detected in the pole plasm even in the presence of both endogenous SmB and SmD3. This suggests that the SmB and SmD3 proteins are independently transported to the pole plasm or that they are transported as heteromeric complexes in which the methylation of both Sm proteins is tightly coupled. In favour of a coupling methylation of an SmB-SmD3 complex is the finding that hypomethylation of SmD3 is accompanied by a decrease in SmB methylation (Gonzalvez et al., 2010). Development 137 (17) Fig. 9. Role of arginine methylation in germline establishment and development. The present study identifies two functional groups of arginine residues in the tail of SmB: the first group includes the three proximal arginine residues and is required for gonad formation during embryogenesis, whereas the second group comprises the last four arginine residues and is involved in at least two processes: development of the egg chambers within the ovaries and polar granule anchoring in the oocyte. As the phenotype associated with the loss of these arginine residues in SmB is distinct from that produced by capsuléen or valois mutations, it is likely that other arginine methyltransferases (such as Dart7) actively participate in SmB methylation during oogenesis. The existence of a methylated factor that is able to bind to Tud and is necessary for its localisation in nuage and pole plasm is inferred from the presence of Tud in the nuage of SmB ovaries expressing a hypomethylated form of SmB and from our previous work (Anne et al., 2007). Tud is also required for SmB localisation in the pole plasm. In a previous report, we determined that the methyltransferase activity of Csul is necessary for the localisation of Tud at the posterior pole of the oocyte (Anne et al., 2007). If one considers a possible association between Tud and methylated SmB in the polar granules, this might explain why unmethylated SmB proteins do not strongly accumulate in pole plasm. Zygotic function of SmB in germ cell migration SmB is first detected in the pole plasm during the early stages of vitellogenesis and persists at this location until pole cell formation. The effect of an absence of SmB methylation could only be uncovered when the maternal contribution of methylated SmB is depleted. The first zygotic abnormality resulting from the arginine substitution in the tail of SmB, and therefore from a defect in SmB methylation, is exposed when the germ cells initiate their migration towards the gonadal mesoderm during late embryogenesis. The function of SmB during these processes remains elusive, but a parallel can be drawn with the occurrence of Sm proteins in the spreading initiation centres (SICs) during the initiation of cell spreading (de Hoog et al., 2004). SICs contain numerous ribonucleoprotein complexes and exist temporarily during the very early stages of cell spreading as precursors of focal adhesions. Cellmatrix attachments play an essential role in many vital cellular processes, including motility, differentiation and survival. Drosophila germ cells leave the midgut and migrate towards the lateral mesoderm, displaying a highly polarised morphology, with a broad lagging edge and an unusually long and relatively stable rear protrusion (Sano et al., 2005). Whether Sm proteins are present in this cell protrusion remains to be determined. DEVELOPMENT 2826 RESEARCH ARTICLE What is the role of SmB in polar granule anchorage? After the transport of the osk mRNA to the posterior pole of the oocyte, this transcript is translated from in-phase alternative initiation codons into two isoforms: a long form and a short form. Owing to its unique ability to promote pole cell formation, only the short Osk isoform can direct the assembly of polar granules (Markussen et al., 1995; Breitwieser et al., 1996). Once polar granule components are recruited by the short form, they must be retained in place to ensure proper assembly of the pole plasm and their inheritance by the pole cells. Polar granule maintenance requires the long Osk form, which is necessary for the efficient anchoring of the short Osk form at the oocyte cortex (Markussen et al., 1995; Rongo et al., 1995; Vanzo and Ephrussi, 2002). Importantly, each Osk isoform exhibits a distinct localisation at the posterior pole: the short Osk is mainly present in the polar granules (Breitwieser et al., 1996), whereas the long Osk form is attached to endocytic membranes (Vanzo et al., 2007). The anchoring of the other pole plasm components to the cortex requires an Oskdependent induction of endocytic activity (Tanaka and Nakamura, 2008). Western blot analysis revealed that both isoforms of Osk are synthesised in SmB; p-{UAS-SmBR4-7} ovaries (data not shown), suggesting that the defective anchoring phenotype cannot be attributed to a failure to produce the long Osk form. Our results suggest that the absence of SmB in pole plasm modifies the organisation of the F-actin cytoskeleton and affects its ability to anchor polar granules at the posterior cortex. Although disruption of the microfilaments by treatment with the actindepolymerising drugs cytochalasin D or latrunculin A exerts only a mild effect on the posterior anchoring of Osk (Babu et al., 2004), several lines of evidence suggest that the F-actin cytoskeleton anchors polar granules to the subcortical posterior region of the oocyte. In particular, the actin-binding proteins Moesin and Bifocal are required for this process (Jankovics et al., 2002; Polesello et al., 2002; Babu et al., 2004). The cortical localisation of Bifocal in the oocyte depends on an intact F-actin cytoskeleton and Bifocal binds directly to F-actin filaments in vitro (Babu et al., 2004), suggesting that Bifocal acts to stabilise F-actin filaments. The endocytic pathway may also function downstream of long Osk to anchor the pole plasm components at the cortex by regulating the dynamics of F-actin (Tanaka and Nakamura, 2008). Projections of F-actin into the ooplasm appear to emanate from the F-actin bundles overlying the posterior cortex of the oocyte. These projections are oskdependent and become detectable from stage 10 onward, when anchoring is required, and are of sufficient length to span the distance between the plasma membrane and the underlying polar granules (Vanzo et al., 2007). In the SmB; p-{UAS-SmBR4-7 or R4-7K} egg chambers, there is a correlation between the loose anchoring of the polar granules and the disorganisation of the F-actin cytoskeleton. Although the possibility that SmB is also present at the cortex cannot be excluded at this stage, a role of SmB in the polar granules in establishing a link between these granules and the cortical cytoskeleton network is plausible. The existence of a positive-feedback loop maintenance mechanism, in which polar granules, possibly in concert with long Osk, enhance their own anchoring at the posterior pole, has indeed been proposed (Vanzo et al., 2007). Acknowledgements This work was performed in the laboratory of Bernard Mechler, whom I thank for constant support. I thank R. Bordonné, A. Ephrussi, S. Kobayashi, P. Lasko, A. Nakimura and the Bloomington Stock Center for providing reagents; G. Gonsalvez and G. Matera for sharing unpublished results; the two reviewers RESEARCH ARTICLE 2827 whose comments helped to improve the quality of the manuscript; and the Nikon Imaging Center (U. Engel and C. Ackermann) at the University of Heidelberg. This work was funded by the Deutsches Krebsforschungszentrum (DKFZ). Competing interests statement The author declares no competing financial interests. Supplementary material Supplementary material for this article is available at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.052944/-/DC1 References Amikura, R., Hanyu, K., Kashikawa, M. and Kobayashi, S. (2001). Tudor protein is essential for the localization of mitochondrial RNAs in polar granules of Drosophila embryos. Mech. Dev. 107, 97-104. Anne, J. and Mechler, B. M. (2005). Valois, a component of the nuage and pole plasm, is involved in assembly of these structures and binds to Tudor and the methyltransferase Capsuléen. Development 132, 2167-2177. Anne, J., Ollo, R., Ephrussi, A. and Mechler, B. M. (2007). Arginine methyltransferase Capsuléen is essential for methylation of spliceosomal Sm proteins and germ cell formation in Drosophila. Development 134, 137-146. Babu, K., Cai, Y., Bahri, S., Yang, X. and Chia, W. (2004). Roles of Bifocal, Homer, and F-actin in anchoring Oskar to the posterior cortex of Drosophila oocytes. Genes Dev. 18, 138-143. Barbee, S. A., Lublin, A. L. and Evans, T. C. (2002). A novel function for the Sm proteins in germ granule localization during C. elegans embryogenesis. Curr. Biol. 12, 1502-1506. Bardsley, A., McDonald, K. and Boswell, R. E. (1993). Distribution of tudor protein in the Drosophila embryo suggests separation of functions based on site of localization. Development 119, 207-219. Bilinski, S., Jaglarz, M. K., Szymanska, B., Etkin, L. D. and Kloc, M. (2004). Sm proteins, the constituents of the spliceosome, are components of nuage and mitochondrial cement in Xenopus oocytes. Exp. Cell Res. 299, 171-178. Boisvert, F.-M., Côté, J., Boulanger, M.-C., Cléroux, P., Bachand, F., Autexier, C. and Richard, S. (2002). Symmetrical dimethylarginine methylation is required for the localization of SMN in Cajal bodies and pre-mRNA splicing. J. Cell Biol. 159, 957-969. Boulanger, M. C., Miranda, T. B., Clarke, S., Di Fruscio, M., Suter, B., Lasko, P. and Richard, S. (2004). Characterization of the Drosophila protein arginine methyltransferases DART1 and DART4. Biochem. J. 379, 283-289. Brahms, H., Raymackers, J., Union, A., de Keyser, F., Meheus, L. and Lührmann, R. (2000). The C-terminal RG dipeptide repeats of the spliceosomal Sm proteins D1 and D3 contain symmetrical dimethylarginines, which form a major B-cell epitope for anti-Sm autoantibodies. J. Biol. Chem. 275, 1712217129. Brahms, H., Meheus, L., de Brabandere, V., Fischer, U. and Luhrmann, R. (2001). Symmetrical dimethylation of arginine residues in spliceosomal Sm protein B/B⬘ and the Sm-like protein LSm4, and their interaction with the SMN protein. RNA 11, 1531-1542. Breitwieser, W., Markussen, F.-H., Horstmann, H. and Ephrussi, A. (1996). Oskar protein interaction with Vasa represents an essential step in polar granule assembly. Genes Dev. 10, 2179-2188. Campos-Ortega, J. A. and Hartenstein, V. (1985). The Embryonic Development of Drosophila melanogaster. Berlin: Springer-Verlag. Chuma, S., Hiyoshi, M., Yamamoto, A., Hosokawa, M., Takamune, K. and Nakatsuji, N. (2003). Mouse Tudor Repeat-1 (MTR-1) is a novel component of chromatoid bodies/nuages in male germ cells and forms a complex with snRNPs. Mech. Dev. 120, 979-990. Côté, J. and Richard, S. (2005). Tudor domains bind symmetrical dimethylated arginines. J. Biol. Chem. 280, 28476-28483. de Hoog, C. L., Foster, L. J. and Mann, M. (2004). RNA and RNA binding proteins participate in early stages of cell spreading through spreading initiation centers. Cell 117, 649-662. Ephrussi, A. and Lehmann, R. (1992). Induction of germ cell formation by oskar. Nature 358, 387-392. Friesen, W. J., Massenet, S., Paushkin, S., Wyce, A. and Dreyfuss, G. (2001a). SMN, the product of the spinal muscular atrophy gene, binds preferentially to dimethylarginine-containing protein targets. Mol. Cell 7, 1111-1117. Friesen, W. J., Paushkin, S., Wyce, A., Massenet, S., Pesiridis, G. S., Van Duyne, G., Rappsilber, J., Mann, M. and Dreyfuss, G. (2001b). The methylosome, a 20S complex containing JBP1 and pICln, produces dimethylarginine-modified Sm proteins. Mol. Cell. Biol. 21, 8289-8300. Girard, C., Mouaikel, J., Neel, H., Bertrand, E. and Bordonné, R. (2004). Nuclear localization properties of a conserved protuberance in the Sm core complex. Exp. Cell Res. 299, 199-208. DEVELOPMENT SmB methylation and germline development Gonsalvez, G. B., Rajendra, T. K., Tian, L. and Matera, A. G. (2006). The Smprotein methyltransferase, dart5, is essential for germ-cell specification and maintenance. Curr. Biol. 16, 1077-1089. Gonsalvez, G. B., Tian, L., Ospina, J. K., Boisvert, F.-M., Lamond, A. I. and Matera, A. G. (2007). Two distinct arginine methyltransferases are required for biogenesis of Sm-class ribonucleoproteins. J. Cell Biol. 178, 733-740. Gonsalvez, G. B., Praveen, K., Hicks, A. J., Tian, L. and Matera, A. G. (2008). Sm protein methylation is dispensable for snRNP assembly in Drosophila melanogaster. RNA 14, 878-887. Gonsalvez, G. B., Rajendra, T. K., Wen, Y., Praveen, K. and Matera, A. G. (2010). Sm proteins specify germ cell fate by facilitating oskar mRNA localization. Development 137, 2341-2351. Hay, B., Ackerman, L., Barbel, S., Jan, L. Y. and Jan, Y. N. (1988). Identification of a component of Drosophila polar granules. Development 103, 625-640. Hermann, H., Fabrizio, P., Raker, V. A., Foulaki, K., Hornig, H., Brahms, H. and Lührmann, R. (1995). snRNP Sm proteins share two evolutionarily conserved sequence motifs which are involved in Sm protein-protein interactions. EMBO J. 14, 2076-2088. Jankovics, F., Sinka, R., Lukácsovich, T. and Erdélyi, M. (2002). MOESIN crosslinks actin and cell membrane in Drosophila oocytes and is required for OSKAR anchoring. Curr. Biol. 12, 2060-2065. Khusial, P. R., Vaidya, K. and Zieve, G. W. (2005). The symmetrical dimethylarginine post-translational modification of the SmD3 protein is not required for snRNP assembly and nuclear transport. Biochem. Biophys. Res. Commun. 337, 1119-1124. Liang, L., Diehl-Jones, W. and Lasko, P. (1994). Localization of vasa protein to the Drosophila pole plasm is independent of its RNA-binding and helicase activities. Development 120, 1201-1211. Liu, J. L., Buszczak, M. and Gall, J. G. (2006). Nuclear bodies in the Drosophila germinal vesicle. Chromosome Res. 14, 465-475. Markussen, F.-H., Michon, A.-M., Breitwieser, W. and Ephrussi, A. (1995). Translational control of oskar generates short OSK, the isoform that induces pole plasm assembly. Development 121, 3723-3732. Massenet, S., Pellizzoni, L., Paushkin, S., Mattaj, I. W. and Dreyfuss, G. (2002). The SMN complex is associated with snRNPs throughout their cytoplasmic assembly pathway. Mol. Cell. Biol. 22, 6533-6541. Mattaj, I. W. (1986). Cap trimethylation of U snRNA is cytoplasmic and dependent on U snRNP protein binding. Cell 46, 905-911. Meister, G. and Fischer, U. (2002). Assisted RNP assembly: SMN and PRMT5 complexes cooperate in the formation of spliceosomal UsnRNPs. EMBO J. 21, 5853-5863. Moore, L. A., Broihier, H. T., Van Doren, M., Lunsford, L. B. and Lehmann, R. (1998). Identification of genes controlling germ cell migration and embryonic gonad formation in Drosophila. Development 125, 667-678. Moussa, F., Oko, R. and Hermo, L. (1994). The immunolocalization of small nuclear ribonucleoprotein particles in testicular cells during the cycle of the seminiferous epithelium of the adult rat. Cell Tissue Res. 278, 363-378. Development 137 (17) Narayanan, U., Achsel, T., Lührmann, R. and Matera, A. G. (2004). Coupled in vitro import of U snRNPs and SMN, the spinal muscular atrophy protein. Mol. Cell 16, 223-234. Ospina, J. K., Gonsalvez, G. B., Bednenko, J., Darzynkiewicz, E., Gerace, L. and Matera, A. G. (2005). Cross-talk between snurportin1 subdomains. Mol. Biol. Cell 16, 4660-4671. Polesello, C., Delon, I., Valenti, P., Ferrer, P. and Payre, F. (2002). Dmoesin controls actin-based cell shape and polarity during Drosophila melanogaster oogenesis. Nat. Cell Biol. 4, 782-789. Raker, V. A., Plessel, G. and Lührmann, R. (1996). The snRNP core assembly pathway: identification of stable core protein heteromeric complexes and an snRNP subcore particle in vitro. EMBO J. 15, 2256-2269. Rongo, C., Gavis, E. R. and Lehmann, R. (1995). Localization of oskar RNA Regulates oskar translation and requires Oskar protein. Development 121, 27372746. Rorth, P. (1998). Gal4 in the Drosophila female germline. Mech. Dev. 78, 113118. Sano, H., Nakamura, A. and Kobayashi, S. (2002). Identification of a transcriptional regulatory region for germline-specific expression of vasa gene in Drosophila melanogaster. Mech. Dev. 112, 129-139. Sano, H., Renault, A. D. and Lehmann, R. (2005). Control of lateral migration and germ cell elimination by the Drosophila melanogaster lipid phosphate phosphatases Wunen and Wunen 2. J. Cell Biol. 171, 675-683. Scofield, D. G. and Lynch, M. (2008). Evolutionary diversification of the Sm family of RNA-associated proteins. Mol. Biol. Evol. 25, 2255-2267. Séraphin, B. (1995). Sm and Sm-like proteins belong to a large family: identification of proteins of the U6 as well as the U1, U2, U4, and U5 snRNPs, EMBO J. 14, 2089-2098. Spradling, A. C. (1993). Developmental genetics of oogenesis. In The Development of Drosophila melanogaster (ed. M. Bate and A. MartinezArias), pp. 1-70. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. Tanaka, T. and Nakamura, A. (2008). The endocytic pathway acts downstream of Oskar in Drosophila germ plasm assembly. Development 135, 1107-1117. Tomancak, P., Guichet, A., Zavorszky, P. and Ephrussi, A. (1998). Oocyte polarity depends on regulation of gurken by Vasa. Development 125, 17231732. Toyooka, Y., Tsunekawa, N., Takahashi, Y., Matsui, Y., Satoh, M. and Noce, T. (2000). Expression and intracellular localization of mouse Vasa-homologue protein during germ cell development. Mech. Dev. 93, 139-149. Vanzo, N. F. and Ephrussi, A. (2002). Oskar anchoring restricts pole plasm formation to the posterior of the Drosophila oocyte. Development 129, 37053714. Vanzo, N., Oprins, A., Xanthakis, D., Ephrussi, A. and Rabouille, C. (2007). Stimulation of endocytosis and actin dynamics by Oskar polarizes the Drosophila oocyte. Dev. Cell 12, 543-555. DEVELOPMENT 2828 RESEARCH ARTICLE