Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Psychoneuroimmunology wikipedia , lookup
Apical dendrite wikipedia , lookup
Neurotransmitter wikipedia , lookup
Subventricular zone wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Circumventricular organs wikipedia , lookup
Chemical synapse wikipedia , lookup
Microneurography wikipedia , lookup
Long-term depression wikipedia , lookup
Signal transduction wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Synaptogenesis wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
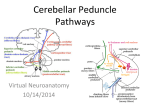
O Cerebellar Anatomy, Biochemistry, and Physiology Introduction The objective of this chapter is to provide an overview of the basic anatomic and functional organization of the cerebellum and its inflow and outflow pathways relevant to medical decision making in children. This information provides a context for understanding the symptoms of congenital, genetic, and acquired ataxias and aids in making decisions about a diagnostic assessment, particularly in cases where the initial clinical presentation and neuroimaging findings are nonspecific. Topics of more limited relevance to children, such as the circulatory system, are omitted. A number of challenges make diagnosis of cerebellar disorders and diseases more difficult in pediatric movement disorders. First, in children these diagnoses are made in the context of a developing motor system. Thus potentially abnormal findings must be judged in the context of the broad limits of normal motor development. Second, movement disorders in children are usually mixed, not pure. Multiple symptoms, including both involuntary movements from the basal ganglia and abnormal motor control or coordination from the cerebellum, may be involved in the same disease process. Third, in the presence of epilepsy, cognitive dysfunction, or behavior problems, medications may be prescribed that precipitate, exacerbate, or cause cerebellar (or basal ganglia) dysfunction. This chapter provides merely an overview. Overview of Cerebellar Structure, Function, and Symptoms Our present, incomplete understanding of cerebellar function and disease has evolved over the last 100 years through painstaking clinical and pathologic observation and ablational studies in animals.1,2 Neuroimaging of structure and function has vastly increased our understanding of the cerebellum in motor control, as well as other functions. Neurophysiologic studies in animals and humans continue to provide new information. Genetic discoveries and the back-and-forth interplay 2 between human genetics and animal models may eventually improve our therapeutics for these diseases. The child’s cerebellar gray and white matter are both developing, resulting in the child learning to control eye movements, muscles of speech, axial truncal muscles, and distal muscles. At a gross structural level, one can think about the spectrum of cerebellar signs in terms of the three functional divisions of the cerebellum: (1) the vestibulocerebellum, in the flocculonodular lobe, involved in axial control and balance and positional reflexes; (2) the spinocerebellum, in the vermis and intermediate part of the cerebellar hemispheres, involved in ongoing maintenance of tone, execution, and control of axial and proximal (vermis) and distal movements; and (3) the cerebrocerebellum, in the lateral part of the hemisphere, involved in initiation, motor planning, and timing of coordinated movements. Functional Anatomy of the Cerebellum and Associated Signs are presented in Table 2-1. Macroscopic to Microscopic Cerebellar Structure The cerebellum contains more than half of all neurons in the central nervous system.3 Its organization is hierarchic and highly regular. Understanding a simplified model of cerebellar neurotransmission and anatomy is helpful for understanding management of diseases and disorders causing ataxia. Understanding this system in greater detail may be useful for making more challenging diagnostic and treatment decisions, as well as for understanding the direction future research and treatments may take. Cerebellar Structural “Threes” Heuristically, three is a helpful mnemonic for remembering cerebellar anatomy. The cerebellum has three major anatomic components that may be affected by focal pathologic processes; three major functional regions that correspond moderately to these components and 9 10 Section 1 OVERVIEW Table 2-1 Functional Anatomy of the Cerebellum Eye Movements Anatomy Vestibulocerebellum Vestibular system afferents to the cerebellar flocculus, paraflocculus, dorsal vermis Function Integration of both position and velocity information so that the eyes remain on target Signs Nystagmus—oscillatory, rhythmic movements of the eyes Impairment with maintaining gaze Difficulties with smooth visual pursuit Undershooting (hypometria) or overshooting (hypermetria) of saccades Speech Anatomy Spinocerebellum—vermis Cerebrocerebellum Sensory afferents from face Corticocerebellar pathway afferents, via pons Function Ongoing monitoring, control of facial muscles Signs Dysarthria, imprecise production of consonant sounds (“ataxic dysarthria”) Dysrhythmia of speech production Poor regulation of prosody; slow, irregularly emphasized (i.e., scanning) speech Trunk Movements Anatomy Vestibulocerebellum Spinocerebellum Sensory, vestibular, and proprioceptive afferents Function Integration of head and body position information to stabilize trunk and head Signs Unsteadiness while standing or sitting, compensatory actions such as use of visual input or stabilization with hands Titubation—characteristic bobbing of the head and trunk Limb Movements Anatomy Spinocerebellum Cerebrocerebellum Sensory and proprioceptive afferents to the spinocerebellum Corticocerebellar pathway afferents via pons Function Integration of input from above—cortical motor areas—about intended commands allows for control of muscle tone in the execution of ongoing movement The spinocerebellum monitors and regulates ongoing muscle activity to compensate for small changes in load during activity and to dampen physiologic oscillation The cerebrocerebellar pathway input contains information about intended movement Signs Hypotonia—diminished resistance to passive limb displacement Rebound—delay in response to rapid imposed movements and overshoot Pendular reflexes Imprecise targeting of rapid distal limb movements Delays in initiating movement Intention tremor—tremor at the end of movement seen on finger-to-nose and heel-to-shin testing Dyssynergia/asynergia—decomposition of normal, coordinated execution of movement—errors in the relative timing of components of complex multijoint movements Difficulties with spatial coordination of hand and fine fractionated finger movements Dysdiadochokinesia—errors in rate and regularity of movements, including alternating movements; the terms asynergia or dyssynergia refer to the inability to coordinate voluntary movements Chapter 2 Cerebellar Anatomy, Biochemistry, and Physiology 11 Table 2-1 Functional Anatomy of the Cerebellum—Cont’d Gait Anatomy Vestibulocerebellum Spinocerebellum Cerebrocerebellum Function Maintenance of balance, posture, tone, ongoing monitoring of gait execution Signs Broad-based, staggering gait subserve somewhat distinct functions; three sets of paired peduncles that carry information into and out of the cerebellum via the pons; three cortical cell layers that interconnect via predominantly glutamatergic and GABAergic signals; and three deep cerebellar output nuclei that transmit cerebellar signal out to ascending and descending tracts. The Three Anatomic Regions— Structures and Afferent Connections The cerebellum has surface gray matter, medullary white matter, and deep gray matter nuclei. Analogous to cerebral gyri and sulci, folia make up the surface of the cerebellum. Beneath the folia, the white matter myelinates during childhood and is susceptible to a wide variety of diseases affecting white matter. Innermost are the deep cerebellar nuclei. The clefts between folia run transversely, demarcating the three main anatomic regions, the flocculonodular, anterior, and posterior lobes, as shown in Figure 2-1 and described in Table 2-2. The Three Cerebellar Functional Regions Connect to Three Deep Cerebellar Nuclei Decades of clinical observations, laboratory animal ablation studies, and more recent imaging studies have informed current views of the three functional regions of the cerebellum. These regions subserve basic functions of execution and integration of information about balance, body position and movement, and motor planning and timing. Output from these regions goes to the deep nuclei. The deep cerebellar nuclei, arranged medially to laterally, are the fastigial, interposed, and dentate nuclei, with the interposed consisting of two nuclei, the globose and emboliform. Anatomy, output nuclei, and function of these regions are described in Table 2-3. The Three Cerebellar Peduncles Three paired sets of peduncles carry fibers to and from the cerebellum. Unlike the basal ganglia, the cerebellum has a direct connection to the spinal cord. Cerebellar connections with the spinal cord Hemisphere Vermis Primary fissure Anterior lobe Horizontal fissure Posterior lobe Posterior fissure Flocculonodular lobe Flocculus Nodulus Figure 2-1. Schematic of the three lobes of the cerebellum (anterior, posterior, flocculonodular) and three anatomic regions (hemispheres, vermis, nodulus). (From Kandel: Principles of neuroscience, ed 4. McGraw Hill Medical, 2000.) Table 2-2 Lobes and Pathways in the Cerebellum Anatomic Region Structures Input Flocculonodular Flocculus—two Vestibular lobe small appendages inferiorly located Nodulus—inferior vermis Anterior lobe A smaller region of the cerebellar hemispheres and vermis anterior to the primary cerebellar fissure Spinal cord— spinocerebellar pathways Posterior lobe Largest, most lateral, and phylogenetically latest region of cerebellar hemispheres Cerebrocortical, via pons and body (spinocerebellar) are ipsilateral. Cerebellar connections with the cerebrum (cerebrocerebellar, via dentate-rubral-thalamic tract) are contralateral. 12 Section 1 OVERVIEW Table 2-3 Summary of Cerebellar Structure and Function Functional Anatomic Output Nuclei Function Vestibulocerebellum Flocculonodular Vestibular nuclei (medulla, not cerebellum) Balance, vestibular reflex, axial control Spinocerebellum Vermis Fastigial nuclei Motor control and execution, axial and proximal muscles Cerebrocerebellum Medial aspect of cerebellar Interposed (globose plus hemispheres emboliform) nuclei Motor control and execution, distal muscles Lateral cerebellar hemispheres Planning, timing coordinated movements Dentate nuclei That is, motor control of the right side of the body is controlled by the left cerebrum with the right cerebellum. Connections from the cerebrum to the cerebellum, via pons, therefore cross on entry and exit. Ascending connections from the spinal cord largely do not. Figure 2-2 shows a schema of the key pathways through the peduncles, and additional detail is provided in Table 2-4. Types of Afferent Fibers There are two distinct types of afferent fibers that carry excitatory signals, predominantly via the inferior and middle peduncles, into the cerebellum. These are the mossy and climbing fibers, as shown in Table 2-5. Both of these fiber types send a few collateral axons to the deep cerebellar nuclei. Corticopontine fibers Neurotransmitters in the Cerebellum Understanding the neurotransmitter systems in basal ganglia allows for more rational decisions about pharmacotherapy. Superior cerebellar peduncle Cerebellum Pons Pontine mossy fibers Middle cerebellar peduncle To vestib. nuclei Dentate Interposed Fastigial Inferior cerebellar peduncle The Three Layers of Cerebellar Cortex Three layers make up the cerebellar cortex.4 A schema of the predominant cells and their interactions is shown in Figure 2-3, and additional detail about these layers and their predominant cell types and functional connections are shown in Table 2-6. To thalamus and red nucleus Climbing fibers from inferior olive Proprioceptive information from spinocerebellar tract (mossy fibers) Figure 2-2. Schematic of the three primary afferent (inferior peduncles and, middle peduncles) and efferent (superior peduncles) pathways of the cerebellum. (From Washington University School of Medicine: Neuroscience tutorial. Basal ganglia and cerebellum. Retrieved from http://thalamus.wustl. edu/course/cerebell.html, 21 September 2009.) See Table 2-4. Table 2-4 Cerebellar Peduncles, Fiber Bundles, and Deep Cerebellar Nuclei Targets Peduncles Afferent and Efferent Fibers Inferior Afferent fibers (to cerebellum) from multiple sources: the vestibular nerve, the inferior olivary nuclei, the spinal cord (dorsal and rostral spinocerebellar, cuneocerebellar, and reticulocerebellar tracts) Efferent fibers (from cerebellum): fastigiobulbar tract projecting to vestibular nuclei, completing a vestibular circuit Middle Afferent fibers: from pons (crossed fibers from cerebral cortex to pontine gray matter nuclei to middle peduncle) Superior Afferent fibers: few fibers from ventral spinocerebellar, rostral spinocerebellar, and trigeminocerebellar projections Efferent fibers: rubral, thalamic, reticular projections from deep cerebellar nuclei—dentate, interposed nuclei Chapter 2 Cerebellar Anatomy, Biochemistry, and Physiology At present, this is much less true in the cerebellum because the main neurotransmitters in the cerebellum are glutamate and gamma-aminobutyric acid (GABA).5 There are limited therapeutic options involving glutamatergic and GABAergic systems for improving ataxia. Glutamate Glutamate, the main excitatory neurotransmitter in the brain, acts at both ionotropic and metabotropic Table 2-5 Functional Anatomy of Mossy and Climbing Fibers Mossy fibers— Excitatory, originating from the primary multiple brainstem nuclei and afferents spinocerebellar tracts, synapse at the granule cells, carry tactile and proprioceptive information Climbing fibers— afferent Excitatory, originating from the inferior olivary nucleus in the cerebellum, climb up to the outer, molecular layer and synapse on the soma and dendrites of the Purkinje cells; carry information critical for error correction Purkinje cell Parallel fiber receptors. The ionotropic glutamate receptors are a diverse group classified into three types—AMPA (alpha-amino3-hydroxy-5-methyl-4-isoxazolepropionic acid), NMDA (N-methyl-d-aspartic acid), and kainate. These are ligand-gated ion channels, meaning that when glutamate binds, charged ions pass through a channel in the receptor center. Both basket and stellate cells in the molecular layer express presynaptic AMPA receptors, to which overflow glutamate from climbing fibers can bind.6 The metabotropic glutamate receptors, which are G-protein–coupled receptors acting via second messengers, are expressed in a developmentally dependent fashion in the cerebellum,7 with mGluR1 receptors playing a significant role in postsynaptic, Purkinje cell signaling. Clinically, this is relevant in paraneoplastic and autoimmune cerebellar diseases.8 For example, mGluR1 antibodies, which can occur in Hodgkin’s disease, cause a combination of acute, chronic/plastic, and degenerative effects in Purkinje cells.9 Glutamate Transporters Glutamate transporters are important for glutamatergic neurotransmission, as well as excitatory neuropathology. Excitatory amino acid transporters (EAAT) 1, 2, and 3 Parallel fiber Purkinje cell + Molecular layer Purkinje cell layer Granular layer + Molecular layer + + + Purkinje cell layer Purkinje cell + Granule cell Climbing fiber Mossy fiber Granule cell Stellate cell Basket Climbing cell fiber Purkinje cell axon Mossy fiber Golgi cell 13 Cerebellar − nuclear cell Granular layer + + To thalamus and descending motor tracts From inferior olive From brainstem nuclei and spinal cord Figure 2-3. Schematic of the three primary cell layers (granular, molecular, and Purkinje) of the cerebellum. (From Apps R, Garwicz M: Anatomical and physiological foundations of cerebellar information processing, Nature Rev Neurosci 6:297–311, 2005.) 14 Section 1 OVERVIEW Table 2-6 Cerebellar Layers, Cell Types, and Function Layer Cells Innermost—granular cell Granule cells layer Outermost—molecular layer Middle—Purkinje cell layer Input/Output and Function Densely packed granule cells receive excitatory input from ascending mossy fibers. Granule cells are the only excitatory cells within the cerebellum. Axons ascend toward outer, molecular layer where they synapse and form parallel fibers. Golgi cells Receive excitatory, glutamatergic26 input from granule cells and provide negative GABAergic feedback to granule cells. Receive glycinergic and GABAergic input from the Lugaro cells.27 Lugaro cells Low prevalence interneurons, receive serotonergic input.19 Inhibit golgi cells. Parallel fibers These are the bifurcated axons from granule cell layers. They have excitatory synapses directly on Purkinje cell dendrites and on stellate and basket interneurons. Stellate and basket cells Excited by glutamatergic input from parallel fibers from granule cells. Output inhibitory on Purkinje cells. Purkinje cells These cells have extensive dendrites in the molecular layer. Cell bodies are in a single layer. Output is inhibitory to deep cerebellar nuclei. are expressed in the motor cortex, but EAAT1 predominates in the cerebellum,10 where it is expressed in Bergmann glial cell processes and is also known as the glutamate aspartate transporter (GLAST). This plays an important role in glutamate reuptake shortly after synaptic release. Excitatory amino acid transporter 4 (EAAT4) is found on extrasynaptic regions of Purkinje cell dendrites and reduces spillover of glutamate to adjacent synapses.11 Colocalization of these transporters with perisynaptic mGluR1 receptors results in competition for glutamate, and this interaction modulates neuroplasticity in the cerebellum.12,13 Gamma-Aminobutyric Acid (GABA) GABA is the major inhibitory neurotransmitter in the cerebellum, as well as the cerebrum. Its synthesis from glutamate is catalyzed by the enzyme glutamic acid decarboxylase (GAD). Anti-GAD antibodies have been reported in adults with ataxia.14 GABA acts via chloride channels to hyperpolarize neurons. GABA receptors include GABA-A and GABA-C receptors, which are ionotropic, and GABA-B receptors, which are metabotropic, G-protein–coupled receptors. GABA-A receptors also have allosteric binding sites for other compounds including barbiturates, ethanol, neurosteroids, and picrotoxin. Baclofen is a GABA-B agonist. GABA-A receptors are predominantly in the granule cell layer,15 where they receive GABA input from the Golgi cells, and to a lesser extent they are present on the molecular layer interneurons, the basket and stellate cells.16 GABA-B receptors are predominantly in the molecular layer.17 Ethanol affects cerebellar function via GABA-a receptor binding, but may also suppress responses in Purkinje cells to mGluR1 excitation from climbing fibers.18 Acetylcholine, Dopamine, Norepinephrine, and Serotonin Acetylcholine, dopamine, norepinephrine, and serotonin19 and their receptors occur in the cerebellum. However, the clinical effects of these neurotransmitter systems in the cerebellum are poorly understood and at this time seem not to be very helpful for ataxia. In general, the medications physicians use to suppress or modify movement disorders, or to improve mood or cognition, do not improve or worsen ataxia. Recognition, in mixed movement disorders, of the cerebellar ataxia component can help with realistic assessment of the probable benefits of pharmacologic interventions. For example, in mixed dystonia and ataxia, the dystonia may respond to anticholinergics but cerebellar symptoms will not. Endocannabinoids Another important neurotransmitter system in the cerebellum is the endocannabinoid (endogenous cannabinoid) system.20,21 This system is involved in so-called retrograde signaling in the hippocampus, basal ganglia, and cerebellum, whereby postsynaptic neurons release endocannabinoids from their dendrites. These endocannabinoids bind to cannabinoid receptor 1 (CB1) on the presynaptic terminal, resulting in a transient suppression of presynaptic neurotransmitter release. Chapter 2 Cerebellar Anatomy, Biochemistry, and Physiology Activation, on Purkinje cells, of metabotropic glutamate receptors subtype 1 (mGluR1) reduces neurotransmitter release from excitatory climbing fibers via this system. In addition, it has recently been shown that GABAergic basket and stellate cells, in the molecular layer, regulate presynaptic neurotransmission from excitatory parallel fibers, from the granule cells. This system is also involved in cerebellar neuroplasticity22,23 and may thereby affect cerebellar contribution to learning. The significance of pathology within this system in children is currently unknown, although both active marijuana use and the exposure to cannabis prenatally may have adverse cognitive effects involving the cerebellum.24,25 Conclusion This overview of cerebellar function provides a framework for understanding cerebellar disorders and diseases, including mixed movement disorders that involve cerebellar function. REFERENCES 1. Holmes G: Clinical symptoms of cerebellar disease and their interpretation. The Croonian lecture III, Lancet 2:59–65, 1922. 2. Holmes G: The cerebellum of man, Brain 62:1–30, 1939. 3. Ghez C: The cerebellum. In Kandel ER, Schwartz JH, Jessell TM, editors: Principles of neural science, New York, 1991, Elsevier. 4. Ito M: Cerebellar circuitry as a neuronal machine, Prog Neurobiol 78:272–303, 2006. 5. Masugi-Tokita M, Shigemoto R: High-resolution quantitative visualization of glutamate and GABA receptors at central synapses, Curr Opin Neurobiol 17:387– 393, 2007. 6. Liu SJ: Biphasic modulation of GABA release from stellate cells by glutamatergic receptor subtypes, J Neurophysiol 98:550–556, 2007. 7. Douyard J, Shen L, Huganir RL, Rubio ME: Differen- tial neuronal and glial expression of GluR1 AMPA receptor subunit and the scaffolding proteins SAP97 and 4.1N during rat cerebellar development, J Comp Neurol 502:141–156, 2007. 8. Hadjivassiliou M, Boscolo S, Tongiorgi E, et al: Cerebellar ataxia as a possible organ-specific autoimmune disease, Mov Disord 23:1370–1377, 2008. 9. Coesmans M, Smitt PA, Linden DJ, et al: Mechanisms underlying cerebellar motor deficits due to mGluR1autoantibodies, Ann Neurol 53:325–336, 2003. 10. Arriza JL, Fairman WA, Wadiche JI, et al: Functional comparisons of three glutamate transporter subtypes cloned from human motor cortex, J Neurosci 14:5559–5569, 1994. 11. Takayasu Y, Iino M, Kakegawa W, et al: Differential roles of glial and neuronal glutamate transporters in Purkinje cell synapses, J Neurosci 25:8788–8793, 2005. 15 12. Otis TS, Brasnjo G, Dzubay JA, Pratap M: Interactions between glutamate transporters and metabotropic glutamate receptors at excitatory synapses in the cerebellar cortex, Neurochem Int 45:537–544, 2004. 13. Wadiche JI, Jahr CE: Patterned expression of Purkinje cell glutamate transporters controls synaptic plasticity, Nature Neurosci 8:1329–1334, 2005. 14. Manto MU, Laute MA, Aguera M, et al: Effects of antiglutamic acid decarboxylase antibodies associated with neurological diseases, Ann Neurol 61:544–551, 2007. 15. Palacios JM, Young WS 3rd, Kuhar MJ: Autoradiographic localization of gamma-aminobutyric acid (GABA) receptors in the rat cerebellum, Proc Natl Acad Sci U S A 77:670–674, 1980. 16. Trigo FF, Chat M, Marty A: Enhancement of GABA release through endogenous activation of axonal GABA(A) receptors in juvenile cerebellum, J Neurosci 27:12452–12463, 2007. 17. Wilkin GP, Hudson AL, Hill DR, Bowery NG: Autoradiographic localization of GABA-B receptors in rat cerebellum, Nature 294:584–587, 1981. 18. Carta M, Mameli M, Valenzuela CF: Alcohol potently modulates climbing fiber-Purkinje neuron synapses: role of metabotropic glutamate receptors, J Neurosci 26:1906–1912, 2006. 19. Dieudonne S, Dumoulin A: Serotonin-driven longrange inhibitory connections in the cerebellar cortex, J Neurosci 20:1837–1848, 2000. 20. Herkenham M, Groen BG, Lynn AB, et al: Neuronal localization of cannabinoid receptors and second messen gers in mutant mouse cerebellum, Brain Res 552: 301–310, 1991. 21. Moldrich G, Wenger T: Localization of the CB1 cannabinoid receptor in the rat brain. An immunohistochemical study, Peptides 21:1735–1742, 2000. 22. Brenowitz SD, Regehr WG: Associative short-term synaptic plasticity mediated by endocannabinoids, Neuron 45:419–431, 2005. 23. Levenes C, Daniel H, Soubrie P, Crepel F: Cannabinoids decrease excitatory synaptic transmission and impair long-term depression in rat cerebellar Purkinje cells, J Physiol 510:867–879, 1998. 24. Chang L, Yakupov R, Cloak C, Ernst T: Marijuana use is associated with a reorganized visual-attention network and cerebellar hypoactivation, Brain 129:1096– 1112, 2006. 25. Smith AM, Fried PA, Hogan MJ, Cameron I: Effects of prenatal marijuana on response inhibition: an fMRI study of young adults, Neurotoxicol Teratol 26:533– 542, 2004. 26. Petralia RS, Wang YX, Niedzielski AS, Wenthold RJ: The metabotropic glutamate receptors, mGluR2 and mGluR3, show unique postsynaptic, presynaptic and glial localizations, Neuroscience 71:949–976, 1996. 27. Dumoulin A, Triller A, Dieudonne S: IPSC kinetics at identified GABAergic and mixed GABAergic and glycinergic synapses onto cerebellar Golgi cells, J Neurosci 21:6045–6057, 2001.