Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Nonsynaptic plasticity wikipedia , lookup
Nerve growth factor wikipedia , lookup
Neurotransmitter wikipedia , lookup
Activity-dependent plasticity wikipedia , lookup
Optogenetics wikipedia , lookup
Electrophysiology wikipedia , lookup
NMDA receptor wikipedia , lookup
End-plate potential wikipedia , lookup
Brain-derived neurotrophic factor wikipedia , lookup
Chemical synapse wikipedia , lookup
De novo protein synthesis theory of memory formation wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Synaptogenesis wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Channelrhodopsin wikipedia , lookup
G protein-gated ion channel wikipedia , lookup
Molecular neuroscience wikipedia , lookup
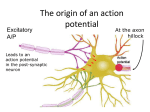
G-protein coupled receptors GPCRs contain seven transmembrane domains with their N’ exoplasmic and their C’ cytosolic. They include receptors for hormones, NTs, photons and thousands of odorant receptors in the nose. The signal transducing G proteins contain three subunits designated alpha, beta and gamma. During signaling, the b-y subunits remain bound together (Gby subunit). Ga subunit is a GTPase that alternates between an active (on) state with bound GTP and an inactive (off) state with bound GDP. Stimulation of a receptor causes activation of the G protein, which mediated the activation of the effector protein. Both subunits may transduce the signal. All effector proteins are ion channels or enzymes that catalyze formation of second messengers (cAMP, DAG, IP3). There are multiple G proteins encoded in the genome. Human have 27 different Ga, 5 Gb and 13 Gy subunits. Both Ga and Gy subunits are linked to the membrane by covalently attached lipids. When no ligand is bound to the receptor, the Ga subunit is bound to GDP and complexed with Gby. In the resting state, the Ga subunit is bound to GDP and complexed with Gby. When an agonist attaches the receptor, it changes its conformation, causing it to bind to the Ga subunit in such a way that GDP is displaced from Ga and GTP becomes bound to the receptor, so the receptor acts as a guanosine exchanging factor (GEF). Once the exchange of nucleotides has occurred, the Ga-GTP complex dissociates from the Gby subunit, but both remain anchored in the membrane. At this point Ga-GTP interacts with and activates an associated effector protein. GTP is hydrolyzed to GDP in seconds, catalyzed by Ga GTPase activity. The resulting Ga-GDP quickly reassociates with Gby, terminating effector activation. A good way to detect the efficiency of metabotropic receptors, is by analyzing them with radiolabeled-GTP in presence or absence of agonists. G-Protein Effectors GP modulation of ion channels On cell recording. Adding ACh to the medium does not increase probability of channel opening. Perfusing with ACh by replacing the solution inside the patch pipette did induce channel opening. When an inside-out patch is formed, the currents disappear again, suggesting an intracellular component regulating the channels. Adding GTP to the medium bathing the inside surface of the membrane causes the channels to reappear. Pertussis toxin, which blocks GDP dissociation from Ga, the channel activity disappears. Finally activity is restored by the mere perfusion with Gby, suggesting these subunits directly open the K channel. By that way, the effects of ACh are localized to a small region, increased cAMP would diffuse through the cytosol, making no difference whether ACh is added by the pippetor or to the bath. Direct modulation of K rectifying currents are thought to occur for GABA, adenosine, glutamate, dopamine, 5-HT and opioid peptides. Rate activation of GPCR Bipolar cells are retinal interneurons that receive synaptic input from rods and cones. The ON cells depolarize when glutamate secretion from photoreceptors is decreased (respond to a light stimulus) and OFF cells hyperpolarize in the same situation. OFF have kainic acid Glut receptors, and ON have mGLuR6, which are GP mediated. We can compare the activation rate of these two kinds of cells. Fast solution change methods are used to make “solution jumps”, that is to place the whole of the bipolar cell into a glutamate containing solution as fast as possible. Cells are jumped first into a solution with an altered K concentration, in order to measure how fast the membrane can be depolarized. The two different cells were then exposed to glutamate and the responses were compared. For the OFF cells the traces are indistinguishable, indicating that glutamate functions as fast as the solution can be changed. ON cells show a 10 ms delay, which is very short. GPCR activated by light Rhodopsin, a GPCR activated by light is localized to the membrane disks that make up the outer segment of rod cells. The G protein coupled to rhodopsin, often called transducin (Gt) is found only in rod cells. There are ~ 10^7 rhodopsin molecules, which consist in the 7-spanning protein, linked to the light absorbing pigment 11-cis-retinal. When a photon is absorbed, the retinal moiety of rhodopsin is converted to the all trans-isomer, causing a conformational change in the opsin portion that activates it. Activated rhodopsin interacts and activates Gt. After ms Gt rapidly dissociates into its component parts and thereby terminating visual signaling. Rod cells potential is -30 mV, and in that depolarization state they are continuously secreting neurotransmitter and the bipolar neurons they stimulate are constantly stimulated. The depolarized state is a consequence of many Na, Ca and K non selective open channels. Absorption of light leads to closing of these channels, causing hyperpolarization. The more absorbed photons, the more negative the membrane potential and the less neurotransmitter released. A single photon produces a decrease in the membrane potential of about 1 mV. Rod cells contains 70 uM of cGMP (cyclic guanosine monophosphate), which is continuously formed from GTP in a reaction catalyzed by guanylyl cyclase. Light absorption induces activation of a cGMP phosphodiesterase, which hydrolyzes cGMP to 5’-GMP, decreasing cGMP concentration upon illumination. In the dark, high levels of cGMP maintain cGMP gated cation channels open, the decrease leads to channel closing and reduces neurotransmitter release. cGMP phosphodiesterase is the effector protein for Gt. Gtalpha-GTP subunit complex generated after light absorption binds to the inhibitory y subunits of cGMP phosphodiesterase, releasing the active catalytic alpha and beta subunits, which then convert cGMP to GMP. A single activated opsin activates 500 Gt molecules. Rod cells adaptation Rod cells are inhibited at high light levels. Visual adaptation permits a rod cell to perceive contrast over a 100000 fold range of ambient light levels. A process contributing to this adaptation involves phosphorylation of activated opsin by rhodopsin kinase. Opsin has three serine phosphorylation sites, the more sites phosphorylated, the less able O to activate Gt and thus induce closing of cGMP gated channels. The extent of O phosphorylation is proportional to the amount of time each O molecule spends in the light activated form, it is a measure of background level of light. Under high light conditions, phosphorylated opsin is abundant and activation of Gt is reduced, thus a greater increase in light level will be necessary to generate visual signal. At high ambient light, the level of opsin phosphorylation is such that the protein b arrestin binds to it, preventing interaction of Gt with O shutting down rod cell activity. In dark adapted cells, virtually all Gta and Gby subunits are in the outer segments, but exposure for 10 minutes to moderate daytime intensities causes over 80 percent of the subunits to move out of the outer segments into other cellular compartments. As a result, Gt proteins are physically unable to bind activated opsin. GPCR that activate phospholipase C Phosphatidylinositol (PI) is a phospholipids which an inositol group extending into the cytosol adjacent to the membrane. The inositol group can be phosphorylated, yielding different membrane bound phosphoinositides. The levels of phosphoinositides are regulated by extracellular signals. One of them, PIP2 binds many cytosolic proteins to the plasma membrane, where they function. PIP2 is also cleaved by the plasma membrane associated enzyme phospholipase C (PLC) generating two second messengers: 1,2 diacylglicerol (DAG) a lipophilic molecule that remains in the membnrane, and inositol 1,2,5-triphosphate (IP3), which diffuses in the cytosol. Ligand binding to receptors coupled to Go or Gq protein (epinephrine and some ACh receptors) induces activation of the isoform PLC-beta by the general GP mechanism. IP3 acts as a ligand to Ca channels in the ER, thus regulating intracellular Ca levels. IP3 binding induce opening of this channel, allowing ions to enter the cytosol. Shortly after being generated, one phosphate of IP3 is hydrolyzed, yielding IP2, which does not stimulate Ca release. Ca increases IP3 affinity to its receptor, thus elevating Ca levels, but higher Ca concentrations reduces that affinity, thus creating Ca oscillations in cells by the IP3 pathway. It is not clear what the purpose of these Ca spikes, rather than increasing cytosolic Ca. One possibility is that a sustained rise in Ca may be toxic to cells. DAG, in turn, remains associated with the membrane. Its principal function is to activate a family of kinases collectively termed protein kinase C (PKC). PKC is present as a soluble cytosolic protein catalytically inactive. A rise in Ca causes PKC to bind to the cytosolic leaflet of the membrane, where DAG activates it. PKC activation thus depend on both IP3 and DAG pathways. GPCR effects on adenylyl cyclase – activation of transcritption The enzyme adenylyl cyclase catalyzes synthesis of the second messenger cAMP. Adenylyl cyclase activation is proportional to the total concentration of Ga-GT resulting from binding of ligands. Different receptors up or down regulate adenylyl cyclase activity. All the effects of cAMP are mediated through protein kinase a (PKA). PKA is a tetramer consisting of two regulatory (R) subunits and two catalytic (C). Each R subunit has two distinct cAMP binding sites. Binding of cAMP results in the release of the associated C subunit, unmasking its catalytic site and activating its inase activity. Gs protein stimulation by receptors leads to activation of PKA PKA translocates to the nucleus and phosphorylates CRE binding protein (CREB), which is found only in the nucleus. Phosphorylated CREb binds to cAMP-response elements (CRE) containing target genes and also interacts with the co-activator CBP/300, which links CREB to the transcriptional machinery, permitting CREB to stimulate transcription. Neurotrophins Initial evidence Neurotrophins were originally characterized because of their functions in differentiation and survival of neurons. The first neurotrophin, NGF, based purified on its ability to promote the neurite growth of sympathetic neurons in chicken. Removal of limb buds from the developing chick embryo leads to a reduction in the number of sensory neurons and motoneurons innervating the bud. Gradting additional limb tissue to a limb bud leads to an increase in the number of neurons in the corresponding regions of the spinal cord and sensory ganglia. This relation was found to result from the selective survival of neurons rather than changes in their proliferations. The ability of neurons to grow axons towards increasing concentrations of NGF finally led to the target field hypothesis of the development of innervation. This postulates that, as most of the developing neurons die during embryogenesis, their survival is dependent on the ability of the neuron to compete for the limited amount of a trophic factor. Those neurons that fail to grow axons to the target then die apoptotically. TFs are synthesized by a wide variety of cell types, including neurons and glia. The factors are secreted extracellularly where they diffuse and act in paracrine fashion. Association of GF with their receptors leads to activation of complex signal transduction pathways, leading to biochemical changes in the cells that mediate cellular survival as well as the acquisition and maintenance of the cellular phenotype. Types of GFs Neurotrophins is the only class acting exclusively in NS that act to support survival and growth of neurons in embryogenesis, and also modify synaptic structure in the adult CNS. Neurotrophic cytokines are a group of molecules highly related to cytokines. Glial derived NTFs exist also as homodimers, but don’t have sequence homology with other NFs. They are synthesized mainly in glial cells. NTFs All neurotrophins have similar biochemical characteristics. They are synthesized as precursor proteins, and then modified to mature forms of 13 kDA, secreted, and they all act as dimers. Approximately half of the amino acid residues in the mature protein are common to all neurotrophins and they all share six cysteine residues at identical positions. This leads to the formation of the disulfide bridges and the similar tertiary structure of the molecules. Neurotrophin receptors The neurotrophins bind to two types of receptors, tyrosine kinase receptors TrkA, TrkB or TrkC and a common neurotrophin receptor p75 (p75NTR) that has no tyrosine kinase domain. NGF binds specifically to TrkA, BDNF and NT-4/5 to TrkB, and NT-3 to TrkC. In addition to these specific receptors, all neurotrophins bind to the p75NTR. Trk receptors also have different splice variants, some of which lack the catalytic tyrosine kinase intracellular domain but the function of these noncatalytic variants is not known. The signal transduction of Trk receptors has been intensively studied and many downstream target molecules identified. Receptor Tyrosine Kinases All RTK have an extracellular ligand binding site, a single hydrophobic transmembrane helix and a cytosolic domain that includes a region with protein tyrosine kinase activity. RTKs are usually monomeric, and receptor binding induces dimerization. In the resting state, the intrinsic kinase activity of TRK is very low. In the dimeric receptor, the kinase in one subunit can phosphorylate one or more tyrosine residues in the other subunit. This leads to a conformational change that facilitates binding of ATP in some receptors, and binding of protein substrates in others. Phosphorotyrosine residues serve as docking sites for proteins involved in downstream signal transduction. Many of these residues interact with adapter proteins, small proteins containing SH2, SH3 domains. These proteins couple activated RTKs to other components of signal transduction pathways, such as those involving Ras activation. Signal Transduction to Ras Ras is a GTP binding protein that, like Galpha subunits, alternates between an activate on state with a bound GTP and an inactive off state with a bound GDP. Ras is not directly linked to cell surface receptors. GTP is present in higher quantities than GDP, therefore it binds to empty Ras molecules, forming active Ras-GTP. Hydrolysis of GTP deactivates Ras. Deactivation of Ras requires the assistance of another protein, GTPase activating protein (GAP). The average lifetime of a GTP bound to Ras is about 1 minute. GAP binds to specific phosphotyrosines in activated RTKs, bringing it close enough to membrane bound Ras GTO to exert its effect on GTP hydrolysis. The protein adapter GRB2, which contains an SH2 domain, binds to a specific phosphorotyrosine residue in the activated receptor. GRB2 binds Sos. Sos catalyzes the conversion of inactive GDP Ras to the active GTP bound form. Ras-GTP activates. Trks react different depending on their cell location. There is evidence supporting that receptors located at distal synapses are endocytosed and retrogradely transported to the cell body, where they activate their signal. NTF effects on adult NS Neurotrophin synthesis, secretion and transport across synapses are tightly regulated by highfrequency activity during synaptic plasticity. In addition, synaptic activity can regulate TrkB incorporation into the plasma membrane, enhancing the chances of ligand–receptor interactions, and consequently the rapid transmission of the signal. Neural activity affects NTFs Figure: Depolarization and cAMP induces BDNF release from AtT20 cells transfected with BDNF. This release is Ca dependent. Neural activity induces expression of NGF, BDNF and NT3. Depolarization induces Trk receptor translocalization to Schaeffer collaterals area from the cellular layer in CA1. It also induces transcript trafficking to the active synapse, where it is translated (the mechanism of this transport is still unknown) NTFs effects on synapses NTFs presynaptic effects During maturation of the presynaptic terminal, neurotrophins might regulate synapse-related protein synthesis and/or modulate protein–protein interactions by regulating post-translational modifications. These actions could lead to an increase in protein incorporation into synaptic vesicles during vesicle biogenesis (1), vesicle accumulation into the reserve pool (2) and/or stabilization of the SNARE (soluble N-ethylmaleimide-sensitive fusion protein-attachment protein receptor) complex — SNAP25 (synaptosome-associated protein, 25 kDa), syntaxin and synaptobrevin. Brain-derived neurotrophic factor (BDNF) might also activate synaptic vesicle mobilization from a reserve pool to a docked pool, possibly through the BDNF-mediated regulation of synapsin 1 phosphorylation (synapsin attaches vesicles to the cytoskeleton, and following AP they are phosphorylated and this apparently induces vesicles movement to docked pool) (3). This model is based on several studies that are discussed in the main text. NTFs postsynaptic effects TrkA-mediated PLCy activation decreases levels of cellular PIP2, which leads to the opening of Transient receptor potentials (TRP) as TRPV1 channels. TRPV1 (VR1) channel or capsaicin receptor, a non-selective cation channel that is activated by heat, noxious vanilloid compounds such as capsaicin, and extracellular protons. Previously, NGF was shown to potentiate the responses of nociceptive sensory neurons to capsaicin. This indicated that crosstalk between capsaicin and NGF occurred within sensory neurons. The idea that TRPV1 channels are necessary for NGF-induced thermal hypersensitivity was also underscored by observations of mice lacking TRPV1. Fig: 10 min NGF incubation of cells expressing TrkA and TRPV1 receptor induces a potentiation in TRPV1 response to extracellular protons (a natural agonist), but does not alter 5-HT response to extracellular 5-HT. A similar effect is obtained by inserting an inside-out patch pipet in a medium containing IP3. PLC AB application blocks this effect. Finally, NGF is known to pronounce pain, as showed by decreased paw withdrawal latency. This effects is abolished in TRPV1 -/- mice. Strikingly, NGF produced an approximately 30-fold increase in proton-evoked currents in Xenopus oocytes that co-expressed TrkA and TRPV1. Diminution of phosphatidylinositol-4,5-bisphosphate levels through antibody sequestration or PLC-mediated hydrolysis mimicked the potentiating effects of NGF at the cellular level. In contrast to NGF-injected normal mice, which showed a marked decrease in paw withdrawal latency in response to a thermal stimulus, injection of NGF into TRPV1-deficient mice did not produce any sensitization. In the case of TrkB and TRPC3, TrkB-mediated PLCy activity leads to the opening of TRPC3 through inositol-1,4,5-triphosphate (Ins(1,4,5)P3) generation and store-operated calcium release. These two receptors are tightly colocalized. BDNF application induces depolarizations and Ca influx through TRPC3 (refer to the article to see why the current was exclusively attributed to this channel). These associations have been confirmed by co-immunoprecipitation experiments. PtdIns(4,5)P2, phosphatidylinositol-4,5-bisphosphate. BDNF was induces rapid depolarization by TrkB interaction with Nav 1.9. A brief (40 ms) application of BDNF to a wide range of neurons (shown a central neuron) induces a rapid response similar than that achieved by a thousand fold higher concentration of glutamate. These responses are dose-dependent. The responses are maintained in a Ca free medium, in the presence of the AP blocker TTX, and the glutamatergic inhibitor CNQX. I.e., the response is not mediated Ca channels or by glutamate release. Na imaging reveals an increase in intracellular Na concentration in response of BDNF application. The effect is blocked by K-252a, a Trk specific inhibitor. Since tyrosine phosphorylation events require nearly a minute, this is difficult to reconcile with the rapid activation pattern. Increasing numbers of interactions between Trk receptors and ion channels are being discovered. Increased tyrosine phosphorylation of NMDA and voltage-gated potassium channels occurs as a result of treatment with BDNF71, 72. In the hippocampus, Trk receptors are expressed both pre- and postsynaptically, and both pre- and postsynaptic mechanisms have been proposed to account for changes in synaptic activity50, 73-76. Electrophysiological measurements show that BDNF can actually suppress KV1.3 currents72 and block postsynaptic -amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptor-mediated currents77. The catalytic activity of TrkB receptors is required for the decrease in AMPA receptor activity, implying that there might be a close association between TrkB and AMPA receptors (Fig. 3). Alternatively, the exo- and endocytosis of AMPA receptors78, which determine activity-dependent changes of synaptic efficacy, could be influenced by BDNF signalling. Fig: LTP induced by pairing a brief BDNF application with a burst of EPSPs. Reconstructed image of a granule cell and a schematic drawing of the experimental arrangement of the stimulation and the BDNF-ejection pipettes. The horizontal lines indicate the orientation of the medial perforant path fibers (MPP). BDNF application alone has no effect on EPSCs. Inset (circled b), train of action potentials produced by the BDNF application. EPSCs (circled a and circled c, top row) were obtained at the time points indicated in the lower part of the graph and are each averages of eight consecutive responses. LTP was induced (at time point 0, open arrowhead) by pairing six EPSPs (50 Hz) with a BDNF pulse of 300 ms. A burst of six EPSPs alone [delivered at the time point marked with an arrow (boxed inset b)] did not change the amplitudes of the test EPSCs. The top row depicts EPSCs before (circled a and c) and after (circled e) induction of LTP. The traces are averages of eight consecutive responses. Inset b, burst of six consecutive EPSPs (stim alone) delivered at the time point marked with an arrow. Inset d, voltage response evoked during pairing a BDNF pulse with an EPSP burst (BDNF + stim). (D) Pooled data from eight experiments; the data points represent the average value of four consecutive responses. Regulation of TrkB trafficking by neuronal and/or synaptic activity. (a) Activity-dependent and BDNF-independent insertion of TrkB at the cell surface. Neuronal and/or synaptic activity induces Ca2+ influx through NMDA receptors and Ca2+ channels even in the absence of BDNF. Subsequent activation of CaMKII facilitates fusion of TrkB-containing vesicles to the cell membranes. (b) BDNF-induced recruitment of TrkB into lipid rafts. Neuronal and/or synaptic activity could trigger local secretion of BDNF. Binding of ligands (BDNF or NT4/5) activates TrkB receptor tyrosine kinase (indicated by red stars), which is crucial for lateral movement of TrkB from non-raft to raft regions of plasma membranes. BDNF-induced recruitment of TrkB into rafts could serve as a mechanism to ensure synapse-specific regulation by BDNF. (c) cAMP-induced translocation of TrkB into the postsynaptic density and gating of TrkB signaling by cAMP. Neuronal and/or synaptic activity also triggers an increase in [cAMP]i, which could contribute synapse-specific modulation in two ways: (i) by facilitating movement of TrkB into spines and/or the postsynaptic density (green arrow) or (ii) by serving as a gate for the signaling of TrkB, which could happen either inside (righthand blue arrow) or outside (left-hand blue arrow) synapses. (d) Activity-dependent endocytosis of the BDNF–TrkB complex. Ca2+ influx through NMDA receptors and Ca2+ channels, as a consequence of local synaptic activity, could enhance TrkB receptor tyrosine kinase activity, which in turn facilitates ligand-induced internalization of TrkB. Endocytosed TrkB remains active (ligandbound) and, together with associated signaling molecules (MAPK, PI3K and PLCγ), forms signaling endosomes. We will consider a model neuron that is receiving multiple excitatory and inhibitory inputs (excitatory and inhibitory postsynaptic potentials — EPSPs and IPSPs) in the somatodendritic region that summate and bring about membrane potential changes at the axon initial segment (AIS). It is in this region that voltage-gated sodium (Nav) and certain voltage-gated potassium (Kv) channels such as the KCNQ channel determine the threshold for firing an action potential, thereby causing action potential generation Kv channels and hyperpolarization-activated cyclic nucleotide-gated (HCN) cation channels on dendrites further control action potential back-propagation, and the time course and extent of the passive spread of synaptic potentials. Back-propagating action potentials might signal the occurrence of recent neuronal excitation and influence synaptic plasticity6, 7, The interaction of these -subunits with auxiliary subunits ( 2, , or ) as well as other proteins can modulate channel function and selectively target some channels (such as Nav, Kv1 and KCNQ) to the axon, other channels (such as HCN, Kv2 and Kv4) to somatodendritic regions, and Kv3 and various Cav isoforms to axons and dendrites (FIG2) Targeting voltage-gated ion channels to axons At the nodes of Ranvier, Nav and KCNQ channels allow currents that spread from one node to initiate an action potential at the next node. Regarding mechanisms for axonal targeting, studies of proteins such as neuron–glia cell adhesion molecule (NgCAM) and vesicle-associated membrane protein-2 (VAMP2) have elucidated at least three feasible models36, 37: directed targeting, transcytosis and selective retention. NgCAM, a chick homologue of L1 CAM, might be transported to the axonal membrane by directed targeting or by transcytosis36, 37, which involves insertion of NgCAM into the somatodendritic membrane, followed by its endocytosis and redistribution to the axonal membrane. By contrast, VAMP2, a synaptic vesicle v-SNARE (soluble N-ethylmaleimide-sensitive fusion protein attachment protein (SNAP) receptor), was uniformly inserted into both the somatodendritic and axonal membranes and then endocytosed from the somatodendritic membrane, leaving VAMP2 surface channels along the axon — a mechanism of selective retention (also known as selective endocytosis or elimination) at the axonal membrane37. These strategies might be used singly or in combination by axonal ion channels. Nav channels. Nav channels associate, or are localized, with a number of molecules that might have a role in anchoring or retaining these channels at the nodes of Ranvier. Nav -subunits have an extracellular Ig-like domain that is similar to those of Ig-family CAMs19, 20, and these -subunits colocalize with several CAMs of this family — neuronal cell adhesion molecule (NrCAM), neurofascin-186 (Nf186) and contactin — around the nodes of Ranvier49, 50. The 1 and 3 subunits interact with Nf186 through their extracellular domains51, whereas NrCAM and Nf186 bind to ankyrin G through a conserved FIGQY motif in their cytoplasmic C-termini, which connects these CAMs to the actin cytoskeleton through IV spectrin52.