Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Biochemical cascade wikipedia , lookup
Oligonucleotide synthesis wikipedia , lookup
Basal metabolic rate wikipedia , lookup
Metalloprotein wikipedia , lookup
Electron transport chain wikipedia , lookup
Light-dependent reactions wikipedia , lookup
Photosynthesis wikipedia , lookup
NADH:ubiquinone oxidoreductase (H+-translocating) wikipedia , lookup
Lactate dehydrogenase wikipedia , lookup
Photosynthetic reaction centre wikipedia , lookup
Microbial metabolism wikipedia , lookup
Biosynthesis wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Nicotinamide adenine dinucleotide wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Blood sugar level wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Biochemistry wikipedia , lookup
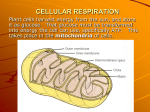
Chapter 4 Carbohydrate Metabolism Glucose transport Metabolism….. Time to put some LIFE into the subject What is Life? What are the properties of life? Movement Turnover of components Reproduction of one’s kind Energy Transformations Chemical Energy is the Dominant Energy Form in a Living System Metabolism: The process by which a living system derives or uses energy through chemical change Energy A B Anabolism: Synthesis. Putting free energy to work Endergonic Catabolism: Degradation. Deriving free energy Exergonic ATP: Energy currency. The standard that is used to gauge all energy compounds The 5 Rules of Energy Metabolism Rule: Living system are able to conserve energy Rule: Heat is wasted energy Heat is energy that cannot be conserved Rule: Living systems will do their utmost to prevent lost of free energy as heat Rule: Exergonic biochemical transformations channel a large part of the free energy into chemical bonds of the product. Rule: Catabolic reactions drive anabolic reactions Anaerobic Aerobic Oxidized cofactors (recycle back Reduced cofactors (drive Ox Phos) The Glycolysis Pathway Major anaerobic pathway in all cells NAD+ is the major oxidant Requires PO4 Generates 2 ATP’s per glucose oxidized End product is lactate (mammals) or ethanol (yeast) Connects with Krebs cycle via pyruvate Glycolysis a-D-Glucose CH2OH O OH Hexokinase Glucose-6-Phosphate ATP CH2OPO3 O OH Phosphoglucoisomerase Fructose-6-Phosphate CH2OPO3 O CH2OH OH CH2OPO3 O CH2OH Fructose-6-Phosphate OH Phosphofructokinase-I Fructose 1,6-Bisphosphate ATP CH2OPO3 O CH2OPO3 OH CHO CH2OPO3 C=O Aldolase CH2OH Dihydroxyacetone-Phosphate H-C-OH CH2OPO3 Glyceraldehyde-3Phosphate ALDOLASE Fructose 1,6bisphosphate CH2OP C=O HO-C-H .. H-C-OH C-OH CH2OP CH2OP C=O HO-C-H .. H + CHO H-C-OH Dihydroxy Acetone Phosphate (DHAP) C-OH CH2OP Glyceraldehyde-3-P Triose Stage Dihydroxy acetone phosphate (DHAP) CH2OPO3 CHO Glyceraldehyde 3-phosphate H-C-OH C=O CH2OH CH2OPO3 Triose phosphate isomerase CHO PO4 H-C-OH CH2OPO3 NAD+ Glyceraldehyde-3-P Dehydrogenase O C ~OPO3 COO H-C-OH ADP H-C-OH ATP CH OPO CH2OPO3 Phosphoglycerate 2 3 NADH + H+ Glycerate 1,3-Kinase Glycerate 3bisphosphate phosphate COO COO H-C-OH H-C-OPO3 CH2OPO3 3-PGA -H2O C~OPO3 PEP CH2OH 2-PGA Phosphoglyceromutase COO CH2 ADP Enolase Pyruvate kinase ATP Back to Glycolysis COO NADH + H+ COO C=O HO-C-H CH3 L-lactate NAD+ CH3 Pyruvate Regulation of Glycolysis 6-phosphofructokinase-1 Allosteric enzyme negative allosteric effectors Citrate , ATP Positive allosteric effectors AMP, fructose1,6-bisphosphate, fructose2,6-bisphosphate Changes in energy state of the cell (ATP and AMP) Regulation of Glycolysis fig.6-4 Regulation of Glycolysis Pyruvate Kinase Allosteric enzyme Isoenzyme in liver Hexokinase Different isoenzymes Hexokinase IV activated by fructose 1,6 bisphosphate glucose 6-phosphate is an allosteric inhibitor inhibited by alanine promote biosynthesis Inhibited by ATP. Regulated by phosphorylation and dephosphorylation The Significance of Glycolysis Glycolysis is the emergency energyyielding pathway Main way to produce ATP in some tissues red blood cells, retina, testis, skin, medulla of kidney In clinical practice Aerobic Oxidation of Glucose 1. 2. 3. Glucose oxidation Oxidation of glucose to pyruvate in cytosol Oxidation of pyruvate to acetylCoA in mitochondria Tricarboxylic acid cycle and oxidative phosphorylation Mechanism of pyruvate dehydrogenase complex Fig.6-6 O2 O2 O2 O2 O2 METABOLISM OF PYRUVATE O2 O2 O2 Its time to get aerobic Pyruvate Structure Look for one NAD+ for each glyceraldehyde-3-PO4 oxidized to pyruvate Carboxyl group (acid) Ketone group (carbonyl) a ketoacid COO -2 0 -OH C=O CH3 Methyl group –2 +2 Net = –02 Oxidation of Carbon –2 CH3 –2 C-OH +2 H-C-OH P CH2OH Glyceraldehyde 3-Phosphate –2 –2 O C +2 C=O O +2 C=O +2 CHO O Decarboxylation Reactions :C O O O +C C O O O Two Types: non-oxidative and oxidative Non-oxidative CO2 H3C-C:COOO NAD+ NADH Oxidative H3C-C: O H+ H3C-C:H O No change in oxidation state of carbonyl C CO2 H3C-C+ O H2 O H3C-C-OH O Oxidized carbonyl C The Energy Story of Glycolysis Overall ANAEROBIC (no O2) Glucose + 2ADP + 2Pi Yeast Glucose + 2ADP + 2Pi 2 Lactate + 2ATP + 2H2O 2 Ethanol + 2CO2 +2ATP + 2H2O Overall AEROBIC Glucose + 2ADP + 2Pi + 2NAD+ 2 Pyruvate + 2ATP + 2NADH + 2H+ + 2H2O 5 ATPs Aerobic C6H12O6 + 6O2 6CO2 + 6H2O Anaerobic CHO H-C-OH OH-C-H H-C-OH H-C-OH CH2OH D-Glucose Go’= -2,840 kJ/mol COOC=O CH3 COO- 146 100 = 2,840 Energy used C=O CH3 2 Pyruvates C6H12O6 2 C3H4O3 Glucose 2 Pyruvate Go’= -146 kJ/mol 5.2% Anaerobic Lactate Glycolysis Glucose Galactose Fructose Mannose pyruvate Fatty Acids Acetyl-Coenzyme A Aerobic Amino Acids Krebs Complex Cycle Pyruvate dehydrogenase 1Pyruvate FADH2 dehydrogenase Dihydrolipoyl transacetylase Thiamin pyrophosphate 3 NADH Lipoic acid Dihydrolipoyl dehydrogenase Oxidative Coenzyme A phosphorylation FAD NAD O2 H2 O + CH2 N N CH3 N CH2 CH2 O P O P O CH3 Vitamin B-1 NH2 O O .. O S Carbanion CH3 C : COO + CO2 O Pyruvate Thiamin pyrophosphate COENZYME A Acetyl-Coenzyme A NH2 N N N O O N O H CH 3 HO H HS-CH2-CH2-N-C-CH2-CH2-N-C-C-C-CH2-O-P-O-P-O CH2 O O O HO CH3 Pantothenate O OH B-vitamin O P O CH3C Acetyl Group O Thioester bond O Adenosine-3’phosphate Dihydrolipoate Long hydrocarbon chain CH2 CH2 CH CH2 CH2 CH2 CH2 COO HS SH 6,8 Dithiooctonoate (Reduced, gained 2 electrons) CH2 CH2 S CH CH2 CH2 CH2 CH2 COO S Disulfide bond (Oxidized, lost 2 electrons) Pyruvate Dehydrogenase Complex Acetyl-CoA .. HS-CoA C-CH3 O .. CH3-C O .. CH3-C NAD+ TPP E1 E2 E3 FAD H2 .. NADH .. OH Pyruvate Dehydrogenase Dihydrolipoyl Transacetylase Dihydrolipoyl dehydrogenase Tricarboxylic Acid Cycle All Mean the Same Features Acetyl-CoA enters forming citrate Citrate is oxidized and decarboxylated 3 NADH, 1 FADH2, and 1 GTP are formed Oxaloacetate returns to form citrate CO2 6 Citrate CO2 5 a-ketoglutarate cis-Aconotate Isocitrate 4 Succinyl-CoA Succinate Cycle Intermediates Fumarate Malate Oxaloacetate More Reduced More Oxidized CH3C ~ S-CoA CARBON BALANCE O 4 Oxaloacetate 4 Malate Citrate 6 2 carbons in 2 carbons out 4 Fumarate Isocitrate 6 CO2 a-ketoglutarate 5 CO2 4 Succinate Succinyl-CoA 4 Reactions of Acetyl-CoA Split here H H C-C~S-CoA .. H O CH3-C~S-CoA O Acetylations or Acylations HS-CoA S-CoA O C=O COO H2C C-OH COO C=O CH2 COO OAA Carbanion Citrate Synthase (a lyase) CH2 COO Citroyl-CoA Citrate CH3-C~SCoA O Citrate Synthase COO- COOC=O CH2 HS-CoA -OOC-CH 2- C-OH CH2 COO- COO- Oxaloacetate (OAA) CH2COOHO-C-COO- Acetyl-CoA CH2COO- Citric Acid or Citrate Isocitrate Formation CH2COO-H2O HO-C-COOH-C-COOH Citrate CH2COOC-COO- +H2O H C-COO- cis-Aconitate Aconitase CH2COOH-C-COO- HO-C-COOH Isocitrate CH2COOH-C-COO- CO2 HO-C-COOH NAD+ NADH + H+ Isocitrate COOCH2 CH2 C=O COO- a-Ketoglutarate Isocitrate Dehydrogenase COOCH2 CH2 C=O NAD+ FAD Lipoic acid HS-CoA TPP COO- a-Ketoglutarate CO2 COOCH2 CH2 C~SCoA O Succinyl-CoA a-Ketoglutarate dehydrogenase Complex Thioester bond energy conserved as GTP COOCH2 CH2 C~SCoA O Pi + GDP GTP Succinyl-CoA HS-CoA COOCH2 CH2 COO- Succinate Succinyl-CoA Synthetase FAD FADH2 NAD+ NADH + H+ H2O COOH COOH C C C H C COOH Succinate H COOH COOH C OH C=O H C COOH Fumarate COOH Malate C COOH Oxaloacetate ATP Generated in the Aerobic Oxidation of Glucose There are two ways for producing ATP Substrate level phosphorylation G1,3-BP to G-3-P, PEP to Pyruvate, SCoA to succinate Oxidative phosphorylation ATP Generated in the Aerobic Oxidation of Glucose In aerobic oxidation of glucose 5 NAD+, 1 FAD Stoichiometry: 2.5 ATP per NADH 1.5 ATP per FADH Table 6-1 Regulation of the Kreb’s Cycle Pyruvate Dehydrogenase complex Pyruvate + TPP Acetal-TPP + CO2 Acetal-TPP + S-S Ac-S ^ SH + TPP Ac-S ^ SH + HS-CoA AcS-CoA + HS ^ SH HS ^ SH + FAD S-S + FADH2 FADH2 + NAD+ FAD + NADH + H+ Pyruvate + HS-CoA + NAD+ Acetyl-CoA + NADH + H+ Regulators-Activators Regulators- Inhibitors and AMP Fatty acids and ATP Key Regulatory Points: 1. Pyruvate dehydrogenase Complex Inhibited by NADH and Acetyl-CoA NADH [NAD+] Acetyl-CoA HS-CoA High NADH means that the cell is experiencing a surplus of oxidative substrates and should not produce more. Carbon flow should be redirected towards synthesis. High Acetyl-CoA means that carbon flow into the Krebs cycle is abundant and should be shut down and rechanneled towards biosynthesis Mechanism: 1. Competitive Inhibition NADH and acetyl-CoA reverse the pyruvate dehydrogenase reaction by competing with NAD+ and HS-CoA 2. Covalent Modification (second level regulation) E-1 subunits of PDH complex is subject to phosphorylation TPP Active FAD HPO4= 1 2 Insulin 3 E1-OH PDH phosphatase H2O ATP PDH kinase E1-OPO3 Inactive ADP Epinephrine Glucagon Cyclic-AMP protein kinase ATP Regulation of the Citric Acid Cycle Primary modes: 1. Substrate availability (key enzymes are subsaturated) Allostery is not a primary mode 2. Product inhibition 3. Feedback inhibition (competitive) Key regulators: 1. Acetyl-CoA (controls citrate synthase) 2. OAA (controls citrate synthase, regulated by NADH) 3. NADH (controls citrate synthase, isocitrate dehydrogenase 4. Calcium (stimulates NADH production) See Fig. 6-9 Pentose Phosphate Pathway PENTOSE PHOSPHATE Pathway Glucose-6-PO4 Ribose-5-PO4 Synthesize NADPH for fatty acid synthesis Metabolize pentoses Take Home: The PENTOSE PHOSPHATE pathway is basically used for the synthesis of NADPH and D-ribose. It plays only a minor role (compared to GLYCOLYSIS) in degradation for ATP energy. 1) NADPH (Nicotinamide Adenine Dinucleotide Phosphate, reduced form) is essentially identical in structure to NADH, with the exception of the phosphate at the 2’-position of the ribose ring of the adenine nucleotide. Just as NADH, the molecule consists of two nucleotides (heterocyclic, aromatic base attached to a ribose sugar at carbon-1 attached to a phosphate at carbon-5) attached to one another by a phosphoanhydride bond linking their 5’-phosphates. NADPH differs from NADH physiologically in that its primary use is in the synthesis of metabolic intermediates (NADPH provides the electrons to reduce them), while NADH is used to generate ATP by contributing its reducing power to the electron transport chain Basic Process Found in cytosol Two phases Oxidative nonreversible Nonoxidative reversible 2) The pentose phosphate pathway serves substantially two functions in cells: to provide ribose (a pentose) and its derivative 2-deoxyribose for nucleic acid synthesis (ribose is the sugar in RNA, 2-deoxyribose in DNA), and to provide NADPH as a reducing agent. The oxidation and decarboxylation of glucose-6-phosphate to ribulose-5phosphate occurs in three steps, accompanied by the generation of two molecules of NADPH. The first step is the oxidation of the hydroxymethylene group at position one to a carbonyl group, yielding a lactone (cyclic ester) and a molecule of NADPH. The second step is then to hydrolyze the lactone to the free carboxylic acid. The carboxyl group of the carboxylic acid is then removed by oxidative decarboxylation, converting the 6-carbon sugar acid to a 5-carbon sugar, with the accompanying production of another molecule of NADPH. 3) Once glucose-6-phosphate has been oxidized and decarboxylated to ribulose-5-phosphate, this latter keto-sugar is converted to the corresponding aldose, ribose-5-phosphate, by the enzyme phosphopentose isomerase. The ribose-5-phosphate produced in this way can now be used in the synthesis of nucleotides for incorporation into nucleic acids. The reaction proceeds through an enediol (C=C double bond and two hydroxyl groups) intermediate, as the enzyme takes advantage of the dissociability of the hydrogen on the terminal hydroxyl group to generate an oxyanion and move the C=O double bond to the terminal carbon, producing the aldehyde and reducing the ketone to an alcohol. 4) In order to control ribose synthesis, a mechanism exists to remove this sugar when it is in excess, by converting it to glycolytic intermediates. A series of three enzymatic steps are carried out, transferring two- and three-carbon fragments from one sugar to another, and all of these steps are similar in mechanism to an aldol condensation (remember that aldolase, the enzyme in glycolysis which fragments the six-carbon, bisphosphorylated sugar fructose1,6-bisphosphate to two phosphorylated three-carbon fragments, breaks the carbon-carbon bond through the reverse mechanism of the aldol condensation). In these cases, however, the enzyme functions by cleaving a fragment from the donor sugar by a reverse aldol condensation, and then attaches it to the acceptor sugar using the forward reaction. The enzymes are transketolase, which transfers a two-carbon fragment terminating on the interior side in a carbonyl, and transaldolase, which transfers a three-carbon fragment terminating on the interior side in a hydroxymethylene group. 5) The first reaction which assists in the conversion of ribose-5phosphate to glycolytic intermediates, catalyzed by transketolase, is the transfer of the 1- and 2-carbons from xylulose-5-phosphate to the 1-carbon of ribose-5-phosphate. This leaves the last three carbons from xylulose-5-phosphate as glyceraldehyde-3-phosphate, the first three-carbon fragment encountered in glycolysis, and sedoheptulose7-phosphate, formed from the ribose-5-phosphate, which is a sevencarbon sugar. 6) Xylulose-5-phosphate is an unusual sugar which is produced from ribulose-5-phosphate, simply by inverting the configuration at carbon-3. This reaction is carried out by the enzyme phosphopentose epimerase, and is freely reversible. Thus, in the first reaction converting ribose-5-phosphate to glycolytic intermediates, both ribose-5-phosphate and ribulose5-phosphate (the latter in the form of xylulose-5-phosphate) are being degraded to other species, and ultimately carried off in glycolysis. 7) The second reaction which leads from intermediates in the pentose phosphate pathway to glycolytic intermediates is mediated by transaldolase. This enzyme transfers a three-carbon fragment (carbons 1, 2 and 3) from the sedoheptulose-7-phosphate just formed in the first reaction to the glyceraldehyde-3-phosphate just formed in the first reaction, yielding a four-carbon fragment, erythrose-4phosphate, and a six-carbon fragment, fructose-6-phosphate. The fructose-6-phosphate is now free to enter the glycolytic pathway. 8) The final reaction leading from intermediates in the pentose phosphate pathway to glycolytic intermediates is carried out by transketolase, just as was the first reaction. In this reaction, another molecule of xylulose-5-phosphate is cleaved, and the two-carbon fragment consisting of carbons 1 and 2 is transferred to the molecule of erythrose-4-phosphate just formed in the transaldolase reaction, yielding a molecule of glyceraldehyde-3-phosphate and another molecule of fructose-6-phosphate. Both of these products are capable of entering glycolysis directly, and so there are no leftover fragments produced in this overall conversion. Because another molecule of xylulose-5-phosphate has entered the reaction, the overall conversion consists of two molecules of xylulose-5-phosphate and one molecule of ribose-5-phosphate going to two molecules of fructose-6-phosphate and one molecule of glyceraldehyde-3phosphate; the xylulose-5-phosphate can be produced from ribose-5-phosphate through ribulose-5-phosphate, and so the net reaction is the removal to glycolysis of three molecules of ribose-5-phosphate. 9) Because the NADPH and ribose-5-phosphate produced by the pentose phosphate pathway are used for quite different purposes, it is sometimes necessary to produce them in different amounts. Therefore, the cell has different modes in which the pentose phosphate pathway can function. In the case where much more ribose-5-phosphate is required than NADPH, the ribose-5-phosphate is produced from glyceraldehyde-3-phosphate and fructose-6-phosphate by running the transaldolase and -ketolase reactions in reverse. This allows the cell’s NADP+ supply to remain essentially unaffected 10) When both NADPH and ribose-5-phosphate are needed in large amounts, the predominant reaction used by the cell to generate them is the conversion of glucose-6-phosphate to ribose-5-phosphate, with the liberation of two molecules of NADPH for each molecule of glucose-6-phosphate converted. 11) When much larger amounts of NADPH are required than ribose5-phosphate, the conversion of glucose-6-phosphate to ribose-5phosphate is the main reaction used, but the ribose-5-phosphate is immediately recycled through the transaldolase and -ketolase reactions, with gluconeogenesis returning the fructose-6-phosphate and glyceraldehyde-3-phosphate to glucose-6-phosphate for another round. 12) An alternative use of the pentose phosphate pathway can be implemented when NADPH is needed in great quantity while ribose-5-phosphate is not. This use involves not recycling the ribose-5-phosphate to glucose-6-phosphate, but rather carrying the glycolytic intermediates forward, rather than backward. The final destination of the ribose-5-phosphate in this case is thus pyruvate, which can enter the Citric Acid Cycle as acetyl CoA and produce ATP. This mode is implemented when the cell requires both NADPH and ATP or NADH, rather than predominantly NADPH. 13) An important use of the NADPH produced in the pentose phosphate pathway is in the maintenance of a reducing environment in the cell. In order to reduce oxidized sulfhydryls back to their free states in the laboratory, we use mercaptoethanol or dithiothreitol, but the cellular equivalent of this reducing agent is glutathione. Glutathione is a tripeptide, similar in structure to Glu-Cys-Gly, but with the exception that the glutamate residue is ligated to the cysteine through the R-group carboxyl, rather than the normal peptide-forming carboxyl (attached to the a-carbon). The sulfhydryl group of the cysteine R-group functions as the reducing agent, and recombines with disulfide bonds in a variety of molecules to release as a free sulfhydryl one of those partners in the disulfide. Another molecule of glutathione carries out the same reaction on the glutathione-subject molecule disulfide, releasing the other partner and producing an oxidized glutathione dimer. NADPH is used to reduce both glutathiones back to the sulfhydryl form, such that they can carry out this reaction again. In this way, the cell protects its components from the activities of reducing agents, as free sulfhydryls perform a variety of needed functions in cellular molecules. Glycogen Formation and Degradation 93% of glucose units are joined by a-1,4glucosidic bond 7% of glucosyl residues are joined by a1,6-glucosidic bonds Fig.6-11 Glycogen Formation and Degradation Main Chain: branch point every 3 units Branch: 5-12 glucosyl residues High Solubility many terminals 4 hydroxyl groups More reactive points for synthesis and degradation. GLYCOGEN SYNTHESIS ENZYMES UDP-glucose pyrophosphorylase Glycogen Synthase forms UDP-glucose major polymerizing enzyme a1.,4->1,6-glucantransferase Glycogen Synthesis Glycogen Degradation Glucose-6-PO4 Synthesis Glucose-1-PO4 UDP-Glucose GLYCOGEN SYNTHESIS ACTIVATION OF D-GLUCOSE GLYCOSYL TRANSFER BRANCHING ACTIVATION UDP-GLUCOSE G-1-P + UTP UDP-GLUCOSE + PPi UDP-Glucose pyrophosphorylase 2 Pi O CH2OH H HO O OH H HN O O O N O P O P O CH2 O H OH O O HO Uridine diphosphate (UDP) Glucose OH Glycogen Glycogen Synthase Phosphorylase UTP Glucose 1-PO4 PPi Activated glucose UDP-Glucose UDP-glucose pyrophosphorylase Go’(kJ mol-1) Glucose 1-PO4 + UTP H2O + PPi Glucose 1-PO4 + UTP + H2O UDP-Glucose + PPi 2 Pi ~0 -33.5 UDP-Glucose + 2 Pi -33.5 The hydrolysis of pyrophosphate drives this reaction O CH2OH H HO HN O O O O OH H GLYCOSYL TRANSFER N O P O P O CH2 O H OH O O HO UDP OH CH2OH O H HO CH2OH O H OH H O H OH OH H O H OH NON-REDUCING END CH2OH CH2OH NEW H HO O OH H H OH H O CH2OH O OH H H OH H O O OH H H OH O BRANCHING Cleave Glycogenin a1.,4->1,6-glucantransferase Glycogen Degradation (Glycogenolysis) Glycogenolysis is not the reverse of glycogenesis Glycogen Breakdown Glycogen Phosphorylase and PO4 Debranching Enzyme Glucose-1-Phosphate Phosphoglucomutase Glucose-6-Phosphate Glucose Glycolysis Take home: Glycogen contributes glucose to glycolysis and to blood glucose (Liver) Phosphorylase O O O HO-P-OH HO-P-OH O HO CH2OH O CH2OH O HO-P-OH O O O O CH2OH CH2OH O O O HO O-P-OH HO O Glucose-1-PO4 PHOSPHORYLYSIS O Glycogen Phosphorylase C N Glycogen Storage Site Can accommodate on 4-5 sugars Pyridoxal 5’-PO4 at active sites N C Phosphorylase: A Homo Dimer * More active 2 H2O Phosphorylase Phosphatase 2 PO4 PHOS A 2 ADP Phosphorylase B Kinase Covalent 2 ATP PHOS B Less Active + 2 AMP + Immediate - 2 AMP Allosteric PHOS B More active Cyclic AMP Hormonal Regulation Debranching Enzyme Highly branched core Phosphorylase Phosphorylase Glycogen Limit Branch a1,41,4 glucantransferase a1,6-gluglucosidase + D-glucose TAKE HOME: DEGRADATION What activates glycogen degradation inactivates glycogen synthesis. SYNTHESIS What activates glycogen synthesis inactivates glycogen degradation H2O PO4 PO4 Phosphorylase a Active Phosphorylase b Less Active Glucose-6-PO4 Glycogen ADP H2O ATP PO4 PO4 Glycogen synthase b Less Active Glycogen synthase a Active Glucose-1-PO4 ADP ATP UDP-Glucose The Significance of Glycogenesis and Glycogenolysis Liver maintain blood glucose concentration Skeletal muscle fuel reserve for synthesis of ATP Glycogen Storage Diseases Deficiency of glucose 6-phosphatase liver phosphorylase liver phosphorylase kinase branching enzyme debranching enzyme muscle phosphorylase Table 6-2 Gluconeogenesis The process of transformation of noncarbohydrates to glucose or glycogen Principal organs liver, kidney Non-carbohydrates glucogenic amino acids lactate glycerol organic acids Phosphatase Blood Glucose PO4 H2O Glucose Kinase G6P Ribose 5-PO4 Glycogen F6P Kinase F1,6bisP PO4 Phosphatase H2O Gly-3-P DHAP 1,3 bisPGA Kinase 3PGA 2PGA PEP Kinase L-lactate Pyruvate OAA Gluconeogenesis Synthesis of glucose de novo (from scratch) An anabolic pathway for the synthesis of glucose from L-lactate or smaller precursors. Significance: Primarily in the liver (80%); kidney (20%) Maintains blood glucose levels The anabolic arm of the Cori cycle Stage I Gluconeogenesis F1,6BP Gly3P DHAP Glycerol 1,3BPGA 3PGA Pyruvate Carboxylase 2PGA L-aspartate 2 PEP PEP carboxykinase PEPCK OAA L-malate 1 L-lactate Pyruvate L-alanine OAA L-malate Mitochondria R5P Pentose Phosphate 4 Glycogen Glucose-6-phosphatase Glucose G6P G1P UDP-glucose Hexokinase F6P 3 Fructose 1,6 - PFK-1 bisphosphatase F1,6BP Stage II Gluconeogenesis Problems: 3 irreversible reactions PEP Go’ = -61.9 kJ per mol Pyruvate F-1,6 bisPO4 F-6-PO4 Glucose-6-PO4 Glucose Go’= -17.2 kJ per mol Go’= -20.9 kJ per mol Take home: Gluconeogenesis feature enzymes that bypass 3 irreversible KINASE steps Second Entry Point for Pyruvate new carboxyl group CH3CCOOH + HCO3 + ATP O HOOC-CH2CCOOH+ ADP + PO= 4 O Pyruvate carboxylase O || ATP + HCO3 C HO + ADP OPO3 CO2 Fixation Reactions O O O || || || C HN Swinging Arm NH HN C O C C NH O N NH O O (CH2)4 COO CH2(CH2)3C CH2(CH2)3C S S Biotin Biocytin N H Lys S Carboxybiocytin Biotin’s only function is to fix CO2 N H Lys Biocytin (the cofactor of biotin) O O C- N Carboxy Biotin N O Carboxy group S CH2 CH2 CH2 CH2 C=0 Swinging Arm HN CH2 CH2 CH2 CH2 Attach to Enzyme at lysine -amine group C Carboxylase Enzyme 3 Bypasses in Gluconeogenesis COO C=O COO GTP GDP CH2 COO OAA C~OPO3 CH2 CO2 PEP PEP Carboxykinase H2O PO4 Fructose 1,6bisPO4 Fructose 1,6 bisphosphatase PO4 H2O Glucose-6-PO4 Glucose 6 phosphatase Fructose-6-PO4 Glucose Liver is a major anabolic organ L-lactate Blood Lactate D-glucose THE CORI CYCLE L-lactate Blood Glucose D-glucose Muscle is a major catabolic tissue Cori Cycle REGULATION FOCUS ON CARBON FLOW L-lactate Glucose (Synthesis) Glucose Pyruvate (Degradation) ENZYMES (Allosteric, cAMP-dependent, organ-specific isozymes) Rule 1. Allosteric are targets of metabolite regulators (effectors) RECIPROCAL REGULATION Rule 2. Kinases in glycolysis; phosphatases in synthesis Exception: PEPCK in synthesis - cAMP POSTIVE EFFECTORS Rule 3. ATP, citrate, acetyl-CoA, G6P turn on synthesis AMP, F2,6BP,turn on degradation NEGATIVE EFFECTORS Rule 4. ATP, acetyl-CoA, citrate,G6P turn off degradation AMP, F2,6BP turn off synthesis The Significance of Gluconeogenesis Replenishment of glucose and maintaining normal blood sugar level Replenishment of liver glycogen “three carbon” compounds Regulation of Acid-Base Balance Clearing the products lactate, glycerol Glucogenic amino acids to glucose Blood Sugar and Its Regulation Blood sugar level 3.89-6.11mmol/l Major source of blood glucose digestion and absorption of glucose from intestine Glycogenolysis and gluconeogenesis Fig.6-18 Regulation of Blood Glucose Concentration Insulin decreasing blood sugar levels Glucagon, epinephrine glucocorticoid increasing blood sugar levels