Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cryobiology wikipedia , lookup
Ultrasensitivity wikipedia , lookup
Lactate dehydrogenase wikipedia , lookup
Proteolysis wikipedia , lookup
Basal metabolic rate wikipedia , lookup
Mitogen-activated protein kinase wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Paracrine signalling wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Signal transduction wikipedia , lookup
Biochemical cascade wikipedia , lookup
Lipid signaling wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Citric acid cycle wikipedia , lookup
Blood sugar level wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
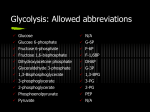
Hormonal regulation and pathologies of carbohydrate metabolism. Diabetes mellitus. Regulation of Glycolysis The rate glycolysis is regulated to meet two major cellular needs: (1) the production of ATP, and (2) the provision of building blocks for synthetic reactions. There are three control sites in glycolysis - the reactions catalyzed by hexokinase, phosphofructokinase 1, and pyruvate kinase These reactions are irreversible. Regulation of Glycolysis Their activities are regulated by the reversible binding of allosteric effectors by covalent modification by the regulation of transcription (change of the enzymes amounts). The time required for allosteric control, regulation by phosphorylation, and transcriptional control is typically in milliseconds, seconds, and hours, respectively. Phosphofructokinase 1 Is the Key Enzyme in the Control of Glycolysis Phosphofructokinase 1 is the most important control element in the mammalian glycolytic pathway. Phosphofructokinase 1 in the liver is a tetramer of four identical subunits. The positions of the catalytic and allosteric sites are identical. High levels of ATP allosterically inhibit the phosphofructokinase 1 in the liver lowering its affinity for fructose 6-phosphate. AMP reverses the inhibitory action of ATP, and so the activity of the enzyme increases when the ATP/AMP ratio is lowered (glycolysis is stimulated as the energy charge falls). A fall in pH also inhibits phosphofructokinase 1 activity. The inhibition of phosphofructokinase by H+ prevents excessive formation of lactic acid and a precipitous drop in blood pH (acidosis). Phosphofructokinase 1 is inhibited by citrate, an early intermediate in the citric acid cycle. A high level of citrate means that biosynthetic precursors are abundant and additional glucose should not be degraded for this purpose. Fructose 2,6-bisphosphate (F-2,6-BP) is a potent activator of phosphofructokinase 1. F-2,6-BP activates phosphofructokinase I by increasing its affinity for fructose 6-phosphate and diminishing the inhibitory effect of ATP. Fructose 2,6-bisphosphate is formed in a reaction catalyzed by phosphofructokinase 2 (PFK2), a different enzyme from phosphofructokinase 1. Fructose 2,6-bisphosphate is hydrolyzed to fructose 6phosphate by a specific phosphatase, fructose bisphosphatase 2 (FBPase2). Both PFK2 and FBPase2 are present in a single polypeptide chain (bifunctional enzyme). Regulation of Glycolysis by Fructose 2,6-bisphosphate When blood glucose level is low the glucagon is synthesized by pancreas Glucagon binds to cell receptors, stimulates the protein kinase A activity Protein kinase A phosphorylates the PFK-2 inhibiting its kinase activity and stimulating its phosphatase activity As result the amount of F-2,6-BP is decreased and glycolysis is slowed. Regulation of Hexokinase Hexokinase is inhibited by its product, glucose 6-phosphate (G-6-P). High concentrations of G-6-P signal that the cell no longer requires glucose for energy, for glycogen, or as a source of biosynthetic precursors. Glucose 6-phosphate levels increase when glycolysis is inhibited at sites further along in the pathway. Glucose 6-phosphate inhibits hexokinase isozymes I, II and III. Glucokinase (isozyme IV) is not inhibited by glucose 6-phosphate. The role of glucokinase is to provide glucose 6-phosphate for the synthesis of glycogen. Regulation of Pyruvate Kinase (PK) Several isozymic forms of pyruvate kinase are present in mammals (the L type predominates in liver, and the M type in muscle and brain). Fructose 1,6-bisphosphate allosterically activates pyruvate kinase. ATP allosterically inhibits pyruvate kinase to slow glycolysis when the energy charge is high. Finally, alanine (synthesized in one step from pyruvate) also allosterically inhibits the pyruvate kinases (signal that building blocks are abundant). The isozymic forms of pyruvate kinase differ in their susceptibility to covalent modification. The catalytic properties of the L (liver) form—but not of the M (brain) form controlled by reversible phosphorylation. When the blood-glucose level is low, the glucagon leads to the phosphorylation of pyruvate kinase, which diminishes its activity. Inhibition 1) PFK-1 is inhibited by ATP and citrate 2) Pyruvate kinase is inhibited by ATP and alanine 3) Hexokinase is inhibited by excess glucose 6-phosphate Regulation of Glycolysis Stimulation 1) AMP and fructose 2,6bisphosphate (F2,6BP) relieve the inhibition of PFK-1 by ATP 2) F1,6BP stimulate the activity of pyruvate kinase Alanine Regulation of Hexose Transporters Several glucose transporters (GluT) mediate the thermodynamically downhill movement of glucose across the plasma membranes of animal cells. GluT is a family of 5 hexose transporters. Each member of this protein family consists of a single polypeptide chain forming 12 transmembrane segments. GLUT1 and GLUT3, present in erythrocytes, endothelial, neuronal and some others mammalian cells, are responsible for basal glucose uptake. Their Km value for glucose is about 1 mM. GLUT1 and GLUT3 continually transport glucose into cells at an essentially constant rate. GLUT2, present in liver and pancreatic -cells has a very high Km value for glucose (15-20 mM). Glucose enters these tissues at a biologically significant rate only when there is much glucose in the blood. GLUT4, which has a Km value of 5 mM, transports glucose into muscle and fat cells. The presence of insulin leads to a rapid increase in the number of GLUT4 transporters in the plasma membrane. Insulin promotes the uptake of glucose by muscle and fat. The amount of this transporter present in muscle membranes increases in response to endurance exercise training. GLUT5, present in the small intestine, functions primarily as a fructose transporter. Regulation of Gluconeogenesis Gluconeogenesis and glycolysis are reciprocally regulated - within a cell one pathway is relatively inactive while the other is highly active. The amounts and activities of the distinctive enzymes of each pathway are controlled. The rate of glycolysis is determined by the concentration of glucose. The rate of gluconeogenesis is determined by the concentrations of precursors of glucose. AMP stimulates phosphofructokinase, whereas ATP and citrate inhibit it. Fructose 1,6bisphosphatase is inhibited by AMP and activated by citrate. Fructose 2,6-bisphosphate strongly stimulates phosphofructokinase 1 and inhibits fructose 1,6-bisphosphatase. During starvation, gluconeogenesis predominates because the level of F-2,6-BP is very low. High levels of ATP and alanine, which signal that the energy charge is high and that building blocks are abundant, inhibit the pyruvate kinase. ADP inhibits phosphoenol-pyruvate carboxykinase. Pyruvate carboxylase is activated by acetyl CoA and Gluconeogenesis is favored when the cell is rich inhibited by ADP. in biosynthetic precursors and ATP. Regulation of the Enzymes Amount by Hormones Hormones affect gene expression primarily by changing the rate of transcription. Insulin, which rises subsequent to eating, stimulates the expression of phosphofructokinase and pyruvate kinase. Glucagon, which rises during starvation, inhibits the expression of these enzymes and stimulates the production of phosphoenolpyruvate carboxykinase and fructose 1,6bisphosphatase. Transcriptional control in eukaryotes is much slower than allosteric control; it takes hours or days in contrast with seconds to minutes. Regulation of Glycogen Metabolism • Muscle glycogen is fuel for muscle contraction • Liver glycogen is mostly converted to glucose for bloodstream transport to other tissues • Both mobilization and synthesis of glycogen are regulated by hormones • Insulin, glucagon and epinephrine regulate mammalian glycogen metabolism Hormones Regulate Glycogen Metabolism Insulin • Insulin is produced by b-cells of the pancreas (high levels are associated with the fed state) • Insulin increases rate of glucose transport into muscle, adipose tissue via GluT4 transporter • Insulin stimulates glycogen synthesis in the liver via the second messenger phosphatidylinositol 3,4,5-triphosphate (PIP3) Glucagon • Secreted by the a cells of the pancreas in response to low blood glucose (elevated glucagon is associated with the fasted state) • Stimulates glycogen degradation to restore blood glucose to steady-state levels • Only liver cells are rich in glucagon receptors and therefore respond to this hormone Epinephrine (Adrenalin) • Released from the adrenal glands in response to sudden energy requirement (“fight or flight”) • Stimulates the breakdown of glycogen to G1P (which is converted to G6P) • Increased G6P levels increase both the rate of glycolysis in muscle and glucose release to the bloodstream from the liver and muscles • Both liver and muscle cells have receptors to epinephrine Effects of hormones on glycogen metabolism Reciprocal Regulation of Glycogen Phosphorylase and Glycogen Synthase • Glycogen phosphorylase (GP) and glycogen synthase (GS) control glycogen metabolism in liver and muscle cells • GP and GS are reciprocally regulated both covalently and allosterically (when one is active the other is inactive) • Covalent regulation by phosphorylation (-P) and dephosphorylation (-OH) • Allosteric regulation by glucose-6-phosphate (G6P) Activation of GP and inactivation of GS by Epinephrine and Glucagone Activation of GS and inactivation of GP by Insulin