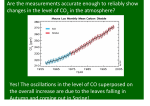
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Global warming controversy wikipedia , lookup
ExxonMobil climate change controversy wikipedia , lookup
Heaven and Earth (book) wikipedia , lookup
Global warming hiatus wikipedia , lookup
Fred Singer wikipedia , lookup
Economics of climate change mitigation wikipedia , lookup
Climate resilience wikipedia , lookup
Climate change denial wikipedia , lookup
2009 United Nations Climate Change Conference wikipedia , lookup
Low-carbon economy wikipedia , lookup
Climatic Research Unit documents wikipedia , lookup
Climate engineering wikipedia , lookup
Mitigation of global warming in Australia wikipedia , lookup
Climate governance wikipedia , lookup
Climate sensitivity wikipedia , lookup
Climate change adaptation wikipedia , lookup
Citizens' Climate Lobby wikipedia , lookup
Instrumental temperature record wikipedia , lookup
Global warming wikipedia , lookup
Physical impacts of climate change wikipedia , lookup
Media coverage of global warming wikipedia , lookup
United Nations Framework Convention on Climate Change wikipedia , lookup
General circulation model wikipedia , lookup
Economics of global warming wikipedia , lookup
Attribution of recent climate change wikipedia , lookup
United Nations Climate Change conference wikipedia , lookup
Climate change in Tuvalu wikipedia , lookup
Politics of global warming wikipedia , lookup
Global Energy and Water Cycle Experiment wikipedia , lookup
Scientific opinion on climate change wikipedia , lookup
Climate change feedback wikipedia , lookup
Solar radiation management wikipedia , lookup
Public opinion on global warming wikipedia , lookup
Climate change in Saskatchewan wikipedia , lookup
Effects of global warming on human health wikipedia , lookup
Climate change in Australia wikipedia , lookup
Effects of global warming wikipedia , lookup
Carbon Pollution Reduction Scheme wikipedia , lookup
Climate change and agriculture wikipedia , lookup
Surveys of scientists' views on climate change wikipedia , lookup
Climate change and poverty wikipedia , lookup
Effects of global warming on humans wikipedia , lookup
Global Change Impacts on Australian Rangelands S.M. Howden, G.M. McKeon and P.J. Reyenga Working Paper Series 99/09 Report to the Australian Greenhouse Office 1999 CSIRO Wildlife and Ecology, GPO Box 284, Canberra ACT 2601 AUSTRALIA Disclaimer The views expressed are not necessarily the views of the Commonwealth, and the Commonwealth does not accept responsibility in respect of any information or advice given in relation to or as a consequence of anything contained herein. The information contained in this report is provided for the purpose of general research and policy development and should not be relied upon for the purpose of a particular matter. Legal advice should be obtained before any action is taken on the basis of any material in this report. CSIRO does not assume liability of any kind whatsoever resulting from any person’s use or reliance upon the contents of this report. Contributing Authors and Affiliations S.M. Howden CSIRO Division of Wildlife and Ecology, GPO Box 284, Canberra, ACT 2601, G.M. McKeon, W.B. Hall, D. Bruget, J.O. Carter, K.A. Day, A. Beswick,and N. Flood Climate Impacts and Grazing Systems, Queensland Department of Natural Resources, 80 Meiers Rd, Indooroopilly, Queensland, 4068. P.J. Reyenga Bureau of Rural Sciences, PO Box E11, Kingston, ACT, 2604, Australia J.L. Moore NERC Centre for Population Biology, Imperial College, Berkshire SL5 7PY UK H. Meinke Agricultural Production Systems Research Unit, DPI/DNR/CSIRO, P.O. Box 102, Toowoomba, Qld, 4350 J.R. Turnpenny Department of Physiology and Environmental Science, University of Nottingham, Leicestershire, LE12 5RD, UK. L. Walker and A.J. Ash CSIRO Tropical Agriculture, PMB Aitkenvale, Qld 4814 J.P. Conroy and 0. Ghannoum University of Western Sydney, Locked Bag 1, Richmond NSW 2753 J.C. Scanlan, Robert Wicks Pest Animal Research Centre, PO Box 318, Toowoomba. Qld. 4350, Australia M. Entel Meteorology CRC, Monash University, Wellington Road, Clayton, Vic 3168 ii Executive Summary • Rangelands are complex systems with the dominant influence of climate affecting strong interactions between plant growth, livestock and human management with additional local controls through factors such as soils and landforms. Consequently, assessments of global change (increased CO2 levels and climate change) impacts and adaptation options are difficult, needing multi-component studies. We have focussed on key drivers from a biophysical viewpoint limiting our assessment of economic and social issues to farm-level economics and management. A later study will deal with broader social, economic, tenure and policy issues within the Western Division of New South Wales. • Doubling of CO2 concentrations by itself is likely to have substantially positive impacts on rangeland forage production and sustainability in many regions of Australia. This occurs largely because increased CO2 concentrations make plants more water-efficient resulting in potentially more growth in water limited environments such as rangelands. The added growth would be particularly pronounced in dry years and in dry regions, resulting in damping of variability in livestock production giving substantially increased farm returns. • The likely increase in grass growth will potentially improve ground cover, thus reducing runoff and soil erosion rates. Safe carrying capacities of livestock are thus likely to increase (0 to 34% assuming no climate change) • The response to CO2 is likely to be limited in sites where soil nutrition is limiting. This will particularly affect the higher rainfall rangelands and those with either poor or degraded soils. • High CO2 levels are likely to significantly increase (+20%) the deep drainage component under pastures which may increase the risks and rates of salinisation in areas where there is a potential hazard. The increase in drainage is particularly likely in sites with poor soil nutrient status and in areas with strongly seasonal rainfall patterns. • If climate change occurs in combination with CO2 rises, the above results will differ. There will remain a sensitivity to rainfall with substantial reductions in rainfall still reducing productivity although in some cases (-10% rainfall) this will just be balanced out by the positive CO2 effects giving pasture productivities little changed from the present. Greater reductions in rainfall (~20%) will reduce plant productivity by about 15%, liveweight gain by 12% and substantially increase variability of stocking rates compared with the present thus having strongly negative impacts on farm income. These assessments do not explicitly take into account increased frequency of El Niño events - an increasing concern. It is important to also note that the latest climate change scenarios suggest strong drying trends. • If rainfall increases along with CO2 levels, there will be some increase in pasture production but this will be limited by nutrient availability more frequently than at present. This will be more of an issue in the higher rainfall regions than in the more arid areas. If warming occurs as well, there could be enhanced nitrogen mineralisation which could partly offset such impacts. The deep soil drainage component is likely to substantially increase (+50% in the site investigated) under high rainfall and high CO2 scenarios giving rise to greatly increased salinity risk and higher rates of salinisation. iii • Increases in CO2 in combination with warmer conditions are likely to lengthen the pasture growing season in many regions (subject to directional change in rainfall) particularly in autumn, resulting in increased liveweight gain potential by livestock. However this is likely to be offset by two factors: 1) forage quality is likely to decrease due to both decreased nutrient contents of leaves with high CO2 and decreased digestibility of C4 grasses with increases in temperature, and 2) there is likely to be significantly increased frequency of heat stress days in livestock. The second issue may be able to be addressed by a combination of breeding strategies and improved animal husbandry but there remains a likelihood of lower productivities across northern Australia. There has been no attempt to combine in a formal way these different factors - the modelling capacity does not currently exist in Australia. • There have been assessments in the literature that increased CO2 concentrations would result in almost total global replacement of C4 grasses (which are currently the major group in all but the southern regions of Australia) by C3 grasses. If true, such a change would have immense implications for productivity, biodiversity and ecosystem function across Australia’s rangelands. We considered a further assessment was needed using an approach that incorporated the strong climatic components that demonstrably affect grass species distributions in Australia. Our assessment is that doubling of CO2 concentrations will result in only limited changes in the distributions of C3 and C4 grasses and that such changes will be moderated by any temperature change (eg small southern movement if temperature warms). • Increases in CO2 have the potential to result in greater storage of carbon in the rangeland region studied (7-17%) provided that fire is kept out of the pastures to the greatest extent possible. However, increased pasture growth is likely to increase burning opportunities substantially and, if these opportunities were used, this could reduce carbon stores and increase net greenhouse gas emissions. Climate changes in addition to CO2 increase is likely to result in only moderate variations to rates of carbon storage although the driest scenarios are likely to slow this down somewhat (4-25%) compared with historical rates. However, with grazing under existing burning practices, all CO2 and climate change scenarios increased the already positive net emissions from the system (by up to a factor of ten) due to increased fuel loads and increased stocking rates. Under most management regimes, there are strong trade-offs in terms of carbon storage (or net emissions) and livestock productivity on the same unit of land (ie. it is possible to have high C storage and low livestock productivity OR vice versa but not high values for both). This doesn’t however, preclude effective integration by using a mosaic of landcover - a strategy already used by some graziers to provide drought reserves. • Under some global change scenarios, increased landuse competition from cropping may occur as crop yields are simulated to increase more than animal productivity in those sites investigated. This will tend to reduce the size of the grazing industries, but perhaps more importantly in the regions affected, result in the higher productivity land-units being allocated to cropping rather than grazing. This could reduce flexibility in meeting new market specifications for livestock. There are also major considerations relating to biodiversity conservation and the potential emission of large quantities of greenhouse gases from land clearing and cultivation. • The global change impacts on farm income addressed above, whilst significant, are expected to be less than current variation in income arising through changes in livestock iv prices or through different management strategies. Global change impacts are expected to be generally greater than impacts arising from changing policies and fuel costs. • There remain several methodological and other issues that need attention. At a fundamental level, the representation of climate change itself can vary the results. Whilst we have adopted approaches that appear to be fairly robust, further research is needed into downscaling climate change data. There remains a lack of integration across the various components of grazing systems with improvements needed in soil nutrient dynamics, livestock production and mortality, landscape-level processes, CO2 responses and treegrass interactions. Entire regional plant communities remain largely unassessed (eg the southern chenopod shrublands and mallee communities). Potentially major issues such as the implications of global change for biodiversity conservation remain unaddressed let alone integration of biodiversity with sustainable agriculture. Integration with social, policy and tenure issues is being attempted within the project through the Western Division of NSW study, however, the resources available for that work are a bare fraction of those needed for a comprehensive analysis. • The studies presented here have generally incorporated some degree of autonomous adaptation to global change, however, more research is needed into explicit adaptation strategies and how these strategies and global change interact to alter greenhouse gas emissions or sinks - thus bringing together research into impacts, adaptation and mitigation. A related concept is to attempt to close the gap between climate variability studies (which tend to address the past and assume that risk as time-independent) and climate change studies (which address the far future and implicitly assume risk is timedependent). Adaptation studies which assess existing trends in climate and projections of future risk in the one to three decade timescale have the potential to merge these two powerful streams of scientific activity and at the same time provide policy-makers and agricultural managers with information pertinent to decisions which fit into shorter, perhaps more tractable timeframes (eg those relating to the Kyoto Protocol). v Contents Executive Summary iii Introduction 1 1. Summary of collaborative studies 3 2. Impacts on native pastures in south-east Queensland, Australia. 8 3. Impacts on fire dynamics in the mulga woodlands of south-west Queensland. 23 4. Potential impacts on C3-C4 grass distributions in eastern Australian rangelands 33 5. Climate change impacts on heat stress and water requirements of cattle in Australia 38 6. Past and future competitiveness of wheat and beef cattle production in Emerald, NE Queensland. 52 vi Introduction The project proposal ‘Global change impacts on Australian Terrestrial Ecosystems’ required delivery on six main themes plus several subsidiary ones. Rangelands is one of the six main themes. There is a large array of work needed to comprehensively address global change impacts on complex systems such as rangelands (Fig. 1). The demands of such a comprehensive analysis was significantly at variance with the relatively small resources available in the project for this theme. For the rangelands component of the project, to achieve the project goals there was a need to: . . . assess existing work on global change impacts on Australian rangelands identify components which have not been effectively addressed to date and which are critical for understanding possible impacts focus on analysis of the above components through direct work and via collaboration with other projects to add value to those so that they address issues of concern here. An excellent synthesis of likely impacts of global change on Australian rangelands was made in 1994 (Stafford Smith et al. 1994) through a workshop involving many of the key Australian and international researchers. Previous work had largely focussed on climate change impacts on forage production (eg McKeon et al. 1988) and on adaptations to such change (eg McKeon et al. 1993). Consequently, the synthesis identified many shortcomings in terms of our knowledge of likely impacts including: . . . . . . . ‘CO2 fertilisation’ impacts sustainability implications landuse change heat tolerance of livestock analyses of species changes and boundary changes, woody weed and fire management carbon sequestration Issues such as impacts on tourism, mining industry, urban areas and conservation management identified in the workshop proceedings were beyond the scope of this study. Since the workshop, three major projects have been initiated in relation to global change and Australian rangelands (apart from this study). These are ‘Evaluation of the impact of climate change on northern Australian grazing industries’ (QDNR and RIRDC), ‘Landuse change in Northern Australia’ (CSIRO and LWRRDC) and ‘Learning from History’ (QDNR and AGO). The focus of these projects was identifying likely climate change impacts as an input into developing more sustainable grazing systems. Through contributions of this project, they were expanded to include impacts of CO2 change (Chapter 2), thus addressing one of the key gaps identified in the workshop synthesis. The inclusion of CO2 into these analyses is critical as this factor is the most probable global change likely over the next century and because of its pervasive impacts on the functioning of ecosystems. The main investigative path of these studies was to look at impacts of global change on plant production through animal production to farm economics. Thus they address several major elements of Fig. 1. However, key elements not addressed include the direct impact of temperature increases on animals, changes in floristics of grasslands (eg alteration of abundance and distribution of C3 and C4 grasses), the changing impacts of fire on system dynamics, carbon stores and emissions and 1 potential competition with other landuses. This report addresses these gaps in knowledge (Chapters 3 to 6) after a review of the results of the collaborative projects (Chapter 1). Region 3 ..... Region 2 ..... Global atmosphere and climate Region 1 by landuse - grazing (cattle, sheep etc), conservation, tourism, Aboriginal use etc Water use efficiency Soil Moisture CO2 Regional climate El Nino Soil C levels Foliage quality Plant production (cover, standing dry matter, growing season, woody weeds) (reduced by high CO2 and temperature) CH4 N2O Animal intake Animal production Heat stress Stock management Other climate CO2 CH4 Farm economics Biodiversity Fire management Emissions policy CO2 CH4 N2O + Figure 1: 2 CO2 source or sink Species 3 Species 2 Species 1 Rainfall Temperature Runoff Erosion Drainage + Social stability Rural policy (climate change adaptation, sustainable resources, farms and societies, biodiversity conservation, education, infrastructure, tenure systems) Conceptual diagram of rangelands systems in relation to global change. 1. Summary of collaborative studies Representation of climate change Pasture growth models require daily climate data and so trials were made using different representations of climate change (McKeon et al. 1998b). These included assessments with actual data, combinations of actual values and calculated variables, the output of stochastic weather generators parameterised with either actual data or data from GCM output. Generally, there were small differences (<10%) between climate change representations of hydrological, biological and managerial effects when averaged across sites although there was some variability in response between sites particularly in the managerial variables (eg stocking rates, burning opportunities). This suggests that the most simple representation of climate change (eg addition of 2oC to daily temperatures or proportionate change in daily rainfall) is suitable for evaluations where the likely change is greater than 10%. This result contrasts with similar studies of wheat production systems (Howden et al. 1999) where the representation of climate resulted in gross differences in mean yields due to the sensitivity of planting rules to the timing and amounts of rain. Howden et al. (1999) found that downscaled GCM data or data derived from stochastic weather generators were less suitable for the task of climate change impacts than simple changes due to their inability to generate an adequate baseline simulation. Further development of these capabilities is needed. Preliminary investigations of CO2 increase A scoping analysis of the effects of doubling of CO2 to 700ppm was performed for six locations (Gayndah, Charters Towers, Charleville, Julia Creek, Alice Springs, Kidman Springs) across northern Australia (Howden et al. 1998). The impacts varied considerably depending on the variable investigated and the location. Pasture growth averaged across the locations was simulated to increase by 15% whilst grass transpiration decreased by 13%. The increased grass cover decreased runoff (12%) which resulted in enhanced soil moisture status which both increased the drainage component (27%) and the length of the growing season (5%). Increased pasture growth over a longer season resulted in increased livestock production (23%), increased opportunities for control burns (33%) and reduced variability of stock numbers (24%). Nitrogen limitations in wet years and in higher productivity sites limited various responses (eg pasture growth, burning opportunities) whilst enhancing others (eg drainage). When increased temperatures and changes in rainfall based on the CSIRO Climate Change Scenarios were evaluated in combination with increased CO2 concentrations, there was generally a small net change from the current situation for northern and north-eastern Australia (0-13% increase in pasture growth) but quite variable impacts for more southern, arid locations (13 to 100% increase). Statewide analyses of impacts of global change on ‘safe’ carrying capacity were made for Queensland (Hall et al. 1998). Baseline case models were based on property data and expert opinion. These models were re-run for a factorial combination of rainfall (+10%), temperature (no change or 3oC increase) and CO2 changes (350ppm or 700ppm). These impacts varied 3 considerably across the State depending on whether moisture, temperature or nutrients were the limiting factors. Without the effect of doubled CO2, warmer temperatures and rainfall changes resulted in -35 to +70% changes in safe carrying capacity. When CO2 was doubled, these changes were -12 to +115% across scenarios and locations. When aggregated to a whole-of-State level the combined scenarios resulted in changes of +3% to +45% in carrying capacity. The results of the above and later studies on CO2 response are based on small-scale, physiologically-based studies often without any management-related treatments. There is growing evidence that such responses are not a complete guide to possible impacts of increased CO2 in the complex, semi-natural Australian rangelands. For example, recent work demonstrates that important Australian C4 grasses may have responses to defoliation which interact with CO2 concentration and that this interaction varies with species (Walker et al. 1999). Field-based experiments such as Free Air CO2 Experiments (FACE) or open-topped chambers are needed with treatments such as clipping and burning to investigate these unexpected responses. An industry-supported proposal for such an experiment has been provided to the AGO (Ash et al. 1999). Assessment of CO2 and climate changes in relation to price, policy and productivity changes Stafford Smith et al. (1999a,b,c) have undertaken an integrated modelling analysis of regional enterprises to simulate how the profitability of a variety of management strategies in grazing systems across northern Australia changes with external drivers including CO2 increase and climate change. The regions addressed are around Charters Towers (NE Qld), Alice Springs (central Australia), the Sturt Plateau/Victoria River District (NT) and the Kimberleys (WA). In each case, intensive collaborative work with property managers had been undertaken to develop a picture of the main management strategies and tactics used in the region, including through accessing detailed property economic records. They used this information in massive combinatorial simulations to attempt to derive simple validated ‘rulesof-thumb’ guidelines for future policy development. Management strategies and tactics (eg trading vs constant stocking vs target pasture utilisation), various CO2 responses and various climate changes were run for site-specific combinations of three levels of pasture response or perennial grass composition, effective production area (+10% and +20% of normal year prices and +20% and +40% of dry year prices), three levels of tree basal area, and with or without policy-related costs associated with transport, the number of livestock, numbers of animals sold or the value of sales. From the huge array of results, a general assessment is that increased economic productivity favours increased stocking rates and higher cash flows regardless of the cause of the increased productivity (eg a ‘better’ climate, CO2 ‘fertilisation’ or higher prices) however, whilst positive climate and CO2 effects do not necessarily place increased pressure on the resource base, changes due to prices or policies may lead to reduced resilience and this change can be quantified. Generally, prices and choice of management strategies had greater effects than climate change and CO2 increase. Policy options generally had small direct effects but could affect choice of management strategy. For Central Australia, climate change, even with modest reductions in rainfall, resulted in higher production which favoured higher stocking rates, improving economic output by $2-9 per animal equivalent (AE) per year, as well as, 4 reducing risks of degradation. This occurred as warming enabled additional growth in cooler seasons. CO2 impacts were not evaluated due to uncertainty of response on these annual pastures but are likely to be positive. For north-east Queensland the climate change scenarios by themselves changed cash flows by +$6/AE/year, whilst increases in CO2 may increase cash flows by $9/AE/year for no significant change in resource impact. In contrast, climate changes are likely to reduce cash flows in the Northern Territory VRD region by -$3 to $9/AE/year whilst CO2 increase may increase returns by $8 to$9/AE/year. In both regions, climate and CO2 effects were approximately additive (ie a negative climate impact and an equal positive CO2 impact cancel each other out). Results for the Kimberleys were not available at the time of going to press. It is important in all the above studies to note that they have not fully incorporated possible changes in climate that would occur if there were significant changes in the frequency and/or intensity of El Niño. There is growing concern that El Niño will become more frequent with global warming (Meehl and Washington 1996, Wilson and Hunt 1997, Timmerman et al. 1999) and suggestions that the increase in El Niño events over recent decades may be related to the observed global warming over this century (Cai and Whetton in prep). Even without El Niño-induced drought, the most recent scenarios of global change from synthesis of many global climate model results suggests substantial reductions in rainfall across Australia (Hulme and Sheard 1999). Mid-range scenarios for the year 2080 suggest 0 to 30% reduction in the rangelands (presumably somewhat drier for the year 2100) and no scenarios suggest significant increase in rainfall. Temperature changes suggested for 2080 are in the range of 2.5 to 3.4oC for mid-range scenarios. References Ash, A.J, Walker, L., Holtum, J., Howden, S.M., and McKeon, G.M. (1999) Climate change and biological carbon sinks: a role for Australia’s rangelands. Proposal to the Australian Greenhouse Office. Cai, W. and Whetton, P.H. (in prep) Evidence for a time-varying pattern of greenhouse warming in the Pacific Ocean. CSIRO Atmospheric Research, 15pp. Hall, W.B., McKeon, G.M., Carter, J.O., Day, K.A., Howden, S.M. & Scanlan, J.C (1998) Climate change in Queenslands grazing lands: II. An assessment of the impact on animal production from native pastures. Rangeland Journal 20: 177-205. Howden, S.M., P.J. Reyenga and H. Meinke, (1999) Global Change Impacts on Australian Wheat Cropping. Report to the Australian Greenhouse Office. CSIRO Wildlife and Ecology, Canberra. pp 121 Howden, S.M., Walker, L., McKeon, G.M., Hall, W.B., Ghannoum, O., Day, K.A., Conroy, J.P., Carter, J.O. & Ash, A.J. (1998) Simulation of changes in CO2 and climate on native pasture growth. pp141-184. In: Evaluation of the impact of climate change on northern Australian grazing industries. Final Report for the Rural Industries Research and Development Corporation. RIRDC, Canberra, Australia. 5 Hulme, M. and Sheard, N. (1999) Climate change scenarios for Australia. Climate Research Unit, Norwich, UK. 6pp. http://www.cru.uea.ac.uk/~mikeh/research/australia.pdf McKeon, G.M., Carter, J.O., Day, K.A., Hall, W.B. and Howden, S.M. (1998a) Evaluation of the impact of climate change on northern Australian grazing industries. Final Report for the Rural Industries Research and Development Corporation. RIRDC, Canberra, Australia. pp 287 McKeon, G.M., Charles, S.P., Bates B.C., and Hall, W.B. (1998b) Methods for evaluating climate change impacts on pasture growth. pp 113-140. In: Evaluation of the impact of climate change on northern Australian grazing industries. Final Report for the Rural Industries Research and Development Corporation. RIRDC, Canberra, Australia. McKeon, G.M., Howden, S.M., Abel, N.O.J. & King, J.M. (1993) Climate change: adapting tropical and subtropical grasslands. Proc. XVII Int. Grasslands Congress, Palmerston North, New Zealand, 1181-1190. McKeon, G.M., S.M. Howden, D.M. Silburn, J.O. Carter, J.F. Clewett, G.L. Hammer, P.W. Johnstone, P.L. Lloyd, J.J. Mott, B. Walker, E.J. Weston & J.R. Wilcocks (1988) The effect of climatic change on crop and pastoral production in Queensland, In Greenhouse. Planning for Climate Change, (ed.) Pearman, G.I., CSIRO pp. 546-563. Meehl, G.A. and Washington, W.M. (1996) El Niño-like climate change in a model with increased atmospheric CO2 concentrations. Nature, 382:56-60. Stafford Smith, D.M., Campbell, B., Steffen, W. and Archer, S. (1994) State-of-the-Science Assessment of the likely impacts of global change on the Australian rangelands. GCTE Working Document No. 14. Canberra, Australia. pp72. Stafford Smith, D.M., Buxton, R., Breen, J., McKeon, G.M., Ash, A.J., Howden, S.M. and Hobbs, T.J. (1999a) Land Use Change in Northern Australia - The impacts of markets, policy and climate change. Regional Report No. 1 - Charters Towers. Report to the Rural Industries Research and Development Corporation and Environment Australia (in prep.) Stafford Smith, D.M., Buxton, R., Breen, J., McKeon, G.M., and Hobbs, T.J. (1999b) Land Use Change in Northern Australia - The impacts of markets, policy and climate change. Regional Report No. 2 - Sturt Plateau and Victoria River District. Report to the Rural Industries Research and Development Corporation and Environment Australia (in prep.) Stafford Smith, D.M., Buxton, R., Breen, J., McKeon, G.M., and Hobbs, T.J. (1999c) Land Use Change in Northern Australia - The impacts of markets, policy and climate change. Regional Report No. 3 - Central Australia. Report to the Rural Industries Research and Development Corporation and Environment Australia (in prep.) Timmermann, A., Oberhuber, J., Bacher, A., Esch, M., Latif, M. and Roeckner, E. (1999) Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature, 398:694-697. 6 Walker, L., Ash, A.J., and Brown, J., (1999) Response of C4 perennial pasture grasses to elevated CO2 and clipping. In: D. Eldridge & D. Freudenberger (eds) Proceedings of the VI International Rangeland Congress, pp 262-263, Townsville, Australia. Wilson, S.G. and Hunt, B.G. (1997) Impact of Greenhouse warming on El Niño/Southern Oscillation behaviour in a high resolution Coupled Global Climate Model. Report to the Department of Environment, Sport and Territories, Australia. 7 2. Impacts on native pastures in south-east Queensland, Australia. S.M. Howden, G.M. McKeon, L. Walker, J.O. Carter, J.P. Conroy, K.A. Day, W.B. Hall, A.J. Ash and 0. Ghannoum, 1. Introduction Since pre-industrial times, atmospheric CO2 concentrations have increased 28% from 280 parts per million volume (ppm) in 1800 to 358 ppm in 1994 (Houghton et al. 1996) mainly as a result of human activities such as burning fossil fuels and landuse change. Further increases in atmospheric CO2 concentrations will occur with predictions for the year 2100 ranging from about 480 ppm to over 800 ppm depending on the economic, resource use and population scenarios used (Houghton et al. 1996). Increasing atmospheric concentrations of CO2 are likely to have significant impacts on plant production and through this on livestock production and resource sustainability. This impact will be through the ‘CO2 fertilisation effect’ where increased CO2 concentrations enhance plant growth as well as through climate changes. This study investigates these potential impacts on grazing in south-east Queensland as part of a larger study looking at global change impacts on Australian agriculture. The response of plants with C3 photosynthetic pathways (some grasses, most forbs, trees and shrubs) to increasing CO2 has been extensively reviewed (e.g. Kimball et al. 1993, Poorter 1993). Controlled environment studies generally demonstrate both increased photosynthetic rates (about 20-30%) and reduced stomatal conductance although there is variation in the degree of response and some exceptions have been reported (Poorter 1993). These changes result in increased biomass accumulation through both enhanced assimilate supply and increased water use efficiency. However, there is strong interaction with other variables such as temperature, soil moisture and soil nutrient availability (Kimball et al. 1993). Hence, it is uncertain whether the potential enhancement will occur, particularly given the low nutrient status of natural ecosystems in northern Australian grazing lands (McKeon et al. 1990). The majority of tropical Australian grazing lands have C4 grasses as the dominant component of understorey vegetation (Hattersley 1983, Hattersley and Watson 1992). The mulga lands are an important exception with C3 grass species dominating in some management situations. The impact of increased CO2 concentrations on C4 grasses is less well documented than for C3 plants. Poorter (1993) reviewed existing experimental data and found an average increase of 28% in dry matter production for C4 species (compared with 71% for C3 species) with doubled CO2. This increase was due to improved water use efficiency as there was no significant difference in assimilation rates. However, some recent studies have shown moderate increases in photosynthetic rates in response to increasing CO2 (Hunt et al. 1996, Morgan et al. 1994) whilst others have shown none (Ghannoum et al. 1997, Nie et al. 1992a, Kirkham et al. 1991), or even reduced photosynthetic rates in moist conditions (Nie et al. 1992b). The increased photosynthesis reported with enhanced CO2 under water limiting conditions (Nie et al. 1992b, Nie et al. 1993) may be due to wetter soil profiles from more conservative water use. 8 The improvement in water use efficiency under high atmospheric CO2 levels is likely to be the most significant effect on C4 pasture grasses. Increases in water use efficiency in such situations occur as a result of reduced stomatal conductance reducing moisture loss while the increased atmospheric CO2 levels maintain internal CO2 concentrations and thus photosynthesis. Increases in water use efficiency of up to 36% have been found in field conditions (Ham et al. 1996, Owensby et al. 1993, Nie et al. 1992b, Kirkham et al. 1991, Morgan et al. 1994, Read and Morgan 1996) with these effects being reduced in wet years (e.g. Knapp et al. 1993). However, in some circumstances, the increase in leaf temperature caused by reduced transpiration (Morison and Gifford 1984, Kirkham et al. 1991, Nie et al. 1992b, Ham et al. 1996) may feedback to increase water use as suggested by Hunt et al. (1996). Several field studies have recorded higher soil moisture contents in elevated CO2 treatment plots (e.g. Nie et al. 1992b, Nie et al. 1993). These findings are potentially important in northern Australian grazing lands as they suggest the possibility of more conservative soil moisture use by pastures and thus an increase in the number of ‘green days’, and hence animal liveweight gain (McCown 1981). In subtropical Queensland the benefit of more conservative water use could be enhanced if the trend of increasing autumn minimum temperatures continues (McKeon and Howden 1993). The longevity of enhanced growth responses to increased CO2 concentrations has been raised as an important issue. There remains some uncertainty as to the mechanisms involved in this ‘acclimation’ process and the degree to which it may occur in the field. Hunt et al. (1996) and Morgan et al. (1994) studying intact grassland communities and Ghannoum et al. (1997) with glasshouse trials have found evidence of photosynthetic acclimation but report that the stomatal responses appeared to be more stable than the photosynthetic responses. Long-term acclimation (as evidenced by reductions in stomatal density over periods of decades) may result in an improved response under drought conditions (Woodward 1987). Improved resilience to drought conditions under conditions of high CO2 may also result through reducing the effects of increasing vapour pressure deficit (VPD). For example, Seneweera et al. (1998) studying the C4 grass Panicum coloratum found that enhanced levels of CO2 offset the impacts of high VPD and low soil water by maintaining higher leaf water potentials. Similar results have been found in other studies (e.g. Nie et al. 1992b, 1993). The above processes suggest potential increases in plant production in Australian tropical grazing lands with increasing CO2 concentrations. Increased grass production provides the opportunity to reduce soil erosion by increasing plant cover and to increase feed availability for livestock. However, increased CO2 may decrease plant nitrogen concentrations (e.g. Hunt et al. 1996) which may reduce liveweight gains in those situations where dietary nitrogen limits feed intake and thus animal growth (Hendricksen et al. 1982). However, Hungate et al. (1997) found no effect of increased CO2 on nitrogen concentration in cool season rangeland annuals. Hence, there remains a considerable need for additional research on the issue of nitrogen dynamics and animal nutrition. Enhanced plant growth under elevated CO2 levels may result in alterations to the relative competitive abilities of species in tropical grasslands. For example, Polley et al. (1994) suggest that increased CO2 levels may have been associated with increased shrub invasions in US savannas during this century due to increased competitive abilities of C3 shrubs over the C4 grasses. This issue is particularly pertinent in Queensland given the existing issue of managing woody weeds and regrowth from cleared areas. However, Archer et al. (1995) and Burrows (1995) have shown that management effects such as frequency of pasture burning 9 are likely to dominate the ecosystem response and that climate and CO2 effects are likely to be of secondary importance. The possibility of changes in the ratio of C3 and C4 grass species has also been raised (Carter and Peterson 1983). However, Owensby et al. (1993) and Nie et al. (1992c) found no evidence that increased CO2 concentrations significantly changed grass species composition with community structure providing a strong buffering capacity. Nevertheless, there remains the possibility that changes in partitioning to roots and shoots (Rogers et al. 1994) or changes in seedhead production (Ghannoum et al. 1997) may result in long term changes. Changes in response to defoliation regimes may also affect community composition although Wilsey et al. (1994) found no effect of elevated CO2 on regrowth rates or allocation patterns in the one African shortgrass savanna species (Sporobolus kentrophyllus) that they studied. A major problem facing the grazing industry is to make reasonable predictions of how increases in CO2 and climate change will interact. The challenge is to extrapolate the physiological knowledge gained on a limited number of species under artificial growing conditions to the complex semi-natural ecosystem of native pastures in northern Australia. Our approach is to use a currently operational model of pasture and animal production (GRASP: McKeon et al. 1990, Carter et al. 1996, Day et al. 1997, Littleboy and McKeon 1997) and examine to what extent it can be modified to represent the CO2 effects reviewed above. Such an approach builds a logical and repeatable pathway to the future allowing the implications of new research to be rapidly evaluated. 2. Simulating CO2 effects 2.1 Description of GRASP GRASP is a model simulating the above-ground yield of a sward dominated by perennial native grasses. GRASP includes a four layer soil water balance and a plant growth model which calculates the processes of run-off, infiltration, drainage, soil evaporation, tree and grass transpiration, pasture growth, consumption and decay, nitrogen uptake, pasture management effects (i.e. stocking rate and pasture burning) and plant density (i.e. perennial grass basal cover). GRASP calculates pasture growth as a function of grass transpiration, radiation interception, temperature, VPD, nitrogen availability and regrowth potential. The model has been parameterised for over 40 native pasture communities in Queensland and has been derived from the results of the last 50 years of field experimentation and grazier experience. Descriptions of the model development include Rickert and McKeon (1982), McKeon et al. (1982), Hendricksen et al. (1982) and McKeon et al. (1990). A full description of each equation is given in Littleboy and McKeon (1997) and evaluation, calibration and validation are described in Carter et al. (1996) and Day et al. (1997). In this preliminary analysis we have yet to consider the effects of CO2 increase and climate change on the other flows of dry matter (detachment, decomposition and consumption) in the grazing system. However, given the generality of parameters for these other processes across a wide range of climates (Day et al. 1997) it is reasonable to concentrate initially on plant growth. 10 The modelling approach used in GRASP has both benefits and limitations in simulating CO2 effects on plant production. The benefits are that the model has been parameterised for a wide range of native pasture communities, soils and locations, and is operational in simulating grazing trials, grazing properties and statewide drought alerts. However, GRASP does not simulate: individual species/varieties in the sward; root growth; phenological development; nor the partitioning of net-photosynthate between roots and shoots in perennial plants which varies with species, phenology, grazing history and soil water. Given the lack of data for native pastures, GRASP has been parameterised, calibrated and validated with only aboveground sward yields. 2.2 CO2 effects on growth, water use and nitrogen parameters in GRASP To simulate pasture growth under enhanced CO2 levels, there is a need to determine how processes and several parameters in GRASP are likely to change. The key parameters are those relating to radiation interception, transpiration and nitrogen dynamics. In this study we do not consider acclimation and hence are concentrating on potential responses derived from growth studies. The evidence reviewed previously suggests that radiation use efficiency (RUE) does not increase in the long term in C4 grasses. Theoretical models (e.g. Chen et al. 1993) suggest an increase in net photosynthesis of 4-10% for a doubling of CO2. For a doubling of CO2, we assume an increase of RUE by 5%. However, we note that Walker et al. (unpublished data) have measured substantial increases (13-60%) in photosynthetic rate for two native C4 grasses in the first year at elevated CO2 under glasshouse conditions. As native pastures are frequently burnt and heavily grazed, regrowth response is an important parameter. The experiment of Walker et al. (unpublished data) provides data specifically on the response of two northern Australian tussock grasses to increased CO2 under weekly defoliation. Doubling CO2 increased regrowth rate by 10% on average. Other studies on young plants and theoretical analysis support this order of magnitude response (Agren 1994) and hence a 10% change has been used in this simulation study. Decreases in stomatal conductance for C4 and C3 species with increasing CO2 are well documented. Reviews of available data suggest that doubling of CO2 concentrations increases water use efficiency, calculated over the seasonal growth period of 100-180 days, by an average 30 - 40%. Similarly, instantaneous measurements of transpiration efficiency (TE: µ mol CO2 s-1 per mmol H2O m2 s -1) show that large increases are possible (30-150% for a doubling of CO2). Despite these increases in instantaneous TE, most studies show that total seasonal water use does not change substantially with increasing CO2, probably due to increased leaf area. Hence we expect GRASP to simulate little change in seasonal water use, but increased green cover and increased seasonal transpiration efficiency expressed as plant growth per mm of seasonal transpiration. Based on the measurements for tropical grasses (Walker et al. unpublished data) and reviewed data we use here a 40% increase in TE for doubled CO2. Transpiration is reduced in GRASP for doubling of CO2 by changing the relationship between green yield (DM kg/ha) and the ratio of potential transpiration to potential evapo-transpiration (ET). Thus changes in the TE parameter are linked to changes in parameters describing the above relationship. However, the relationship between green yield and proportion of radiation intercepted is not expected to change. 11 Modelling studies (e.g. Sellers et al. 1996) and experimental data referred to earlier suggest that the above effect of reducing daily transpiration with increasing CO2 could increase land and canopy ‘surface’ temperature above the increase that would result from just the CO2 effect on radiative forcing. In GRASP the effect of reducing transpiration on leaf surface temperature could be represented by re-calculating daily temperature, vapour pressure deficit and potential evapo-transpiration (e.g. Class A pan) which have been input from the daily climate file. To determine the value of such an approach the effect of transpiration on ambient temperatures and VPD was examined using a spatial version of GRASP (Carter et al. 1996) simulating soil water in Queensland’s grazing lands over the last 40 years. For one month (January) years with similar solar radiation conditions (21-24 MJ/m2/day) were used to establish multiple regression relationships between monthly averages of solar radiation and VPD. The relationship changed with simulated monthly evapo-transpiration (calculated from soil evaporation and grass transpiration). From this analysis it was calculated that a reduction in monthly evapo-transpiration of 20% (e.g. from 60mm to 48mm) results in a 0.4oC increase of average temperature and maximum temperature; 1.2 hPa increase in VPD; and 0.4mm increase in potential ET (i.e. Class A pan). However, even in January in the middle of the ‘wet’ season, monthly rainfall is well below potential ET, and hence it is unlikely that changing daily TE and daily transpiration as indicated above will actually change monthly ET and the driving climate variables at a monthly or seasonal time scale. Sensitivity studies indicated small effects of the above CO2 changes on seasonal ET. Thus we do not currently represent any effect of changing CO2 on daily climatic inputs. GRASP includes relationships between senescence (death of green material) and frost and soil water. Of particular importance is the relationship between maximum possible green cover and soil water availability. Measurement of leaf area of maize under increasing water stress showed that about 50% higher leaf area could be maintained under high CO2 (Gifford 1988) at a given soil moisture level. This was similar to the change in stomatal conductance, and hence we assume in GRASP that the green cover able to be supported for a given level of soil water changes in proportion to transpiration efficiency, i.e. 40% increase in this case. A nitrogen mineralisation index is calculated in GRASP as a function of surface (0-10cm) soil moisture and air temperature using a similar approach to Parton et al. (1988). Although we suggest that the index represents the climatic potential for nitrogen (N) mineralisation, we have not used it in the following simulation studies to change nitrogen availability for the following reasons. The calculated mineralisation index was compared with measured nitrogen yield for over 100 years x site combinations of regularly burnt or mown exclosures across Queensland. The results indicated a general plateau of N yield of about 20-25 kg N/ha for undisturbed native pastures (Day et al. 1997). This plateau of total seasonal N uptake was in some cases reached by the middle of the growing season. Even if subsequent soil moisture conditions were apparently favourable for mineralisation, no further uptake occurred (e.g. Norman 1963). However, increases in soil moisture under enhanced CO2 appear to have increased nitrogen mineralisation in mediterranean ecosystems (Hungate et al. 1997) and increased temperatures are also likely to increase mineralisation rates (Parton et al.1988). Thus, the incorporation of a soil carbon:nitrogen model into GRASP may be needed before these effects can be adequately simulated and at this stage the parameter ‘maximum aboveground N uptake per year’ is not changed with increasing CO2. In GRASP, the rate of N uptake, before maximum uptake is reached, is parameterised as kg N/ha per 100 mm of transpiration (3-10 kg N/ha per 100 mm) depending on location and species. Reduced transpiration with increased CO2 will reduce rate of N uptake in this case. 12 However, reduced uptake and increased mineralisation conditions are likely to lead to increased N concentrations in soil water, and hence increased rate of uptake per mm of transpiration. Sensitivity studies showed that a 20% increase in this parameter for doubled CO2 resulted in approximately the same rate of N uptake as occurred in ambient conditions (e.g. Hungate et al. 1997). GRASP uses a critical nitrogen content in dry matter (%N) to calculate a nitrogen index used to limit radiation use efficiency and regrowth. C4 grasses are able to continue above-ground growth with concentrations of nitrogen as low as 0.4 to 0.8 % N in dry matter with variation relating to differing leaf/stem ratios. Any changes in leaf/stem partitioning with enhanced CO2 levels will thus influence plant critical nitrogen levels as well as affecting potential dry matter production through changes in nitrogen use efficiency. However, data for northern Australia native grasses (L. Walker et al. unpublished data) do not indicate substantial changes in proportion of stem and in other tropical grasses the effects are inconsistent, with both decreases (Seneweera et al. 1998) and increases (Ghannoum et al. 1997) observed. Reductions in the concentrations of Rubisco in leaves under high CO2 levels have been frequently measured and there is some experimental evidence (e.g. Rogers et al. 1996) that this reduces critical leaf nitrogen contents in crop plants although there is no evidence of the same effect in tropical grasses. Considering the above, no change is made in critical nitrogen concentrations with CO2. The availability of young green leaf in native pastures is one of the major driving variables of steer liveweight gain (Poppi et al. 1981, Ash et al. 1982) through the effects of diet selection for leaf on dietary nitrogen concentration (Hendricksen et al. 1982). Several approaches to simulating animal (sheep and cattle) liveweight gain have been evaluated with GRASP ranging from nitrogen and energy balance systems (Hendricksen et al. 1982, Hall 1996) to annual climatic indices (McCown 1981, Hall 1996). Simple seasonal or annual climatic indices have so far proved more successful than mechanistic models in accounting for the observed spatial and temporal variation (Hall 1996) and this approach has been used here to simulate the effect of CO2 change on annual liveweight gain through its effects on soil water and temperature. However, possible animal production effects of changes in pasture composition (Henderson et al. 1994) in response to climate change and CO2 increase are yet to be considered. The above review has examined in detail the necessary changes to parameters in GRASP to represent the effects of doubled atmospheric CO2. Confirmation of many of the changes is dependent on current research on response of native grasses and trees to CO2. It is vital that research on those species important to the productivity of Australia’s grazing lands is continued to be supported so that prediction of impact of future changes can be made. Day et al. (1997) have parameterised GRASP for several sites in south-east Queensland. Parameter values for the typical black speargrass communities in south-east Queensland for the major driving relationships (as described above) and their changes under enhanced CO2 levels are shown in Table 1. The model simulates pasture growth reasonably effectively over the current range of year-to-year climatic variability (Figure 1) although testing of the model under extreme climatic conditions is not yet complete. Table 1 Parameter values changed in GRASP for current (1 x CO2) and doubled CO2 (2 x CO2) scenarios for C4 native pastures in south-east Queensland. 13 GRASP parameters Potential regrowth (kg/ha/day) Potential regrowth /unit grass basal cover Transpiration efficiency (kg/ha/mm @ 20hPa) Green yield at which potential transpiration is 50% of potential ET Rate of N uptake (kg N/ha per 100 mm transpiration) Radiation use efficiency (kg/ha per MJ/m2) 1xCO2 15.0 3.5 13.5 1000 2 xCO2 16.5 3.85 18.9 1400 6.0 12 7.2 12.6 Simulated yield (kg/ha) 5000 y = 1.0168x R2 = 0.7828 4000 3000 2000 1000 0 0 1000 2000 3000 4000 5000 Observed yield (kg/ha) Figure 1 Observed and simulated yield (i.e. peak standing pasture dry matter) for a native C4 pasture sward at ‘Ronnies Paddock’ site, Brian Pastures Research Station, south-east Queensland for 1987-96 with Gayndah daily rainfall and other climate variables. 3. Scenario definition GRASP was used with the above parameter changes to explore possible outcomes from a range of global change scenarios for the location of Gayndah in south-east Queensland. The CO2 and climate scenarios were; 1) current climate + CO2 (350 ppm); 2) current climate + doubled CO2 (700 ppm); 3) temperature increase (+2.76oC) + 700 ppm CO2; 4) reduced rainfall (-24% summer, -12% winter) + temp. change + 700 ppm CO2; and 5) increased rainfall (+12% winter) + temp. change + 700 ppm CO2. These scenarios are named ‘Baseline’, ‘CO2’, ‘Warm’, ‘Warm/Dry’ and ‘Warm/Wet’ respectively. These scenarios are drawn from the CSIRO 1996 scenarios as described in Reyenga et al. (1997) and were used to modify the historical climate record from 1957 to 1996. The grazing management strategies adopted were to: 1) adjust stocking rates in June of each year to use 30% of standing pasture dry matter over the next six months; and 2) burning of a third of the pasture in years when standing pasture dry matter exceeded 1200 kg/ha. These strategies resulted in an implicit adaptation of overall stocking rate and burning practice to changes in pasture growth. 4. Results Doubling of CO2 concentration was simulated to have significant impacts on soil hydrology, plant growth and animal production. Mean seasonal transpiration efficiency was increased by 14 about 10% as a result of both reductions in transpiration (-2%) and increases in growth (8%). Reduced transpiration and runoff enhanced soil moisture status, increasing through-drainage by 20% (Table 2). The growth response to enhanced CO2 was greatest during dry years, showing up to 65% increases when compared with current CO2 levels (Figure 2 and Table 3). Runoff was decreased due to increased mean dry matter levels and higher minimum cover (61%, Table 4). Increased dry matter levels and small increases in growing days (1%) resulted in both increased animal production per hectare (10%) and a markedly reduced coefficient of variation of stocking rates (-27%) (Table 3). However, growth was nitrogen-limited in these simulations with seasonal nitrogen uptake reaching its maximum in several years and this possibly restricted expression of climatic variability. The 6% increase in the climate-derived mineralisation index and increased dry matter flow suggest that nitrogen supply could change under these scenarios and hence there is a need for a more detailed analysis of nitrogen mineralisation in future studies. Table 2 Mean annual transpiration (mm), runoff (mm) and drainage (mm) for the baseline conditions and mean % change from baseline for the four scenarios. Scenario Baseline CO2 Warm Warm/Dry Warm/Wet Transpiration 399 -2 0 -28 +4 Runoff 19 -21 -11 -53 -11 Drainage 20 +20 +20 -65 +50 Growth response (2CO2/1CO2) 2 1.5 1 0.5 0 0 200 400 600 800 Rainfall (mm) 1000 1200 Figure 2. The simulated growth response to enhanced CO2 (ratio of 2CO2/1CO2) with rainfall for a C4 native pasture in south-east Queensland. Each point represents one year from 1959 to 1996. Table 3 Mean pasture growth (kg/ha/year), liveweight gain (kg/ha/year), the coefficient of variation (%) of the stocking rate, and simulated wheat yields (g/m2) from Reyenga et al. 1997 for the baseline conditions and mean % change from baseline for the four scenarios. Scenario Baseline CO2 Warm Warm/Dry Growth 4516 +8 +7 -13 LWG/ha 75.4 +11 +19 -12 CV of SR 36.1 -27 -10 +40 Wheat yield 206 +35 +15 0 15 Warm/Wet +7 +20 -13 +26 Table 4 Mean percentage of days of the year with growth index greater that 0.05, minimum standing dry matter and mean mineralisation index for the baseline conditions and mean % change from baseline for four scenarios. Scenario Baseline CO2 Warm Warm/Dry Warm/Wet %GI days 81 +1 +13 +9 +13 SDM min. 324 +61 -13 -15 +71 Mineralisation 62 +6 +27 +8 +31 The Warm scenario, with both increased CO2 and temperature, tended to remove some of the effects of the CO2 change alone resulting in smaller increases in transpiration efficiency (7%) and drainage. However, compared with the baseline results, simulated animal production increased markedly (19%) due to increases in the number of growing days in the cooler months (13%). Similarly, the 27% increase in mineralisation index suggests that additional nitrogen could become available under such conditions, reinforcing the need to better simulate processes affecting nitrogen availability. The two scenarios with altered rainfall in addition to the CO2 and temperature changes showed varying responses. The Warm/Dry scenario showed a 21% increase in seasonal transpiration efficiency and significantly reduced runoff and drainage compared with the baseline. Under this global change scenario, the positive effects of CO2 and temperature on animal production are negated. The Warm/Wet scenario shows small increases in seasonal transpiration efficiency (3%) but retains most of the positive response to the CO2 and temperature change. Drainage is increased substantially (50%) compared with the baseline conditions. 5. Discussion This simulation study has certain caveats which have important implications for interpretation. The major purpose of the study was to evaluate how an empirical operational pasture model such as GRASP could be modified to reasonably simulate the effects of CO2 and climate change scenarios. The CO2 effects have been derived from recent growth chamber and/or glasshouse studies on tropical grasses. Thus, this study represents our first attempt to link ecosystem modelling to physiological studies. Similarly, the studies with climate change scenarios represent our first attempt to evaluate alternative representations of climate change and their interaction with doubled CO2. However, it must be remembered that we have evaluated only an ‘average’ native pasture without trees and the climate change data are small samples (38 years) of possible climate change representations. Nevertheless, the simulation results using the above representation of CO2 effects highlight : 1) the beneficial effects on plant growth of doubling CO2 in dry years and the mean increase in soil moisture observed experimentally; 16 2) the effects on plant growth are likely to lead to increased animal production through increased pasture productivity, length of growing season, reduced variability and greater opportunities for pasture burning; 3) erosion may be reduced due to lower runoff and increased ground cover but increased drainage may pose problems where there are salinity and waterlogging risks; and 4) nitrogen limitations on plant growth may restrict the beneficial impacts of increased CO2. This study suggests that empirical models such as GRASP can be plausibly modified to include the effects of CO2 as measured in growth chambers and glasshouses once measurements for relevant species and appropriate conditions (low nutrient availability, high VPD, and frequent water stress) are available. However, the parameter changes made to represent the effects of doubled CO2 were derived from short term (i.e. seasonal) growth chamber experiments. Longer term studies using open-topped chambers or Free Air Carbondioxide Experiments are required to determine whether these effects are permanent responses to elevated CO2. Nevertheless, the above preliminary simulation analysis indicated that the potential beneficial effects are worthy of further research. The simulations are limited by lack of knowledge of the impact of CO2 on additional processes operating in complex semi-natural ecosystems such as grazed native pastures. For example, the effects of climate change and management on nutrient cycles, especially nitrogen and phosphorus, will play a large role in limiting or amplifying the effects of CO2 increase. Furthermore, the interactions of individual plant species or ecotype attributes with increased CO2 are yet to be considered. The simulated liveweight gain per hectare provides only an indication of change in animal production. Graziers have a large number of possible adaptation options given the available variation in animal type (sheep, cattle), product (wool, calves, steers, cows), breed (e.g. Bos indicus, Bos taurus) and pasture management. Furthermore, several important issues in animal production are yet to be considered: the nutritional effects of digestibility, leaf/stem ratio, species composition, availability of browse, landscape driven redistribution of runoff, soil mineral limitations such as phosphorus, and compensatory gain. In addition, Howden and Turnpenny (1997) suggest that under the global warming scenario used here, the frequency of heat stress days will increase markedly (by about 140%) in cattle in this location in south-east Queensland. Despite these caveats we consider that the most important impacts of CO2 and climate change will be on plant growth which drives the animal productivity of the grazing system (Day et al. 1997) through quantity, quality, seasonality and year-to-year variation. Explicit modelling of the whole soil-plant-animal production-management system is now required to examine to what extent changes in plant growth and variability translate into changes in economic performance (i.e. Campbell et al. 1997). In this study we have simulated a likely grazier response to changes in plant growth by burning a third of the pasture whenever yields exceeded 1200kg/ha; and adjusting animal numbers each year to eat a fixed proportion of the standing dry matter in June (i.e. end of the growing season) over the next 12 months. This type of approach allows calculation of the effects of the likely adaptation to climate change and CO2 increase in relation to these particular management decisions. When combined with an emissions budget approach (Moore et al. 1997) this will enable assessment of both mitigation options and the potential feedbacks of global change into atmospheric composition. 17 Global change impacts on production and economic performance of the grazing industry will not happen in isolation from other agricultural pursuits. In the Gayndah region of south-east Queensland, cropping is an alternative landuse on some land units. Reyenga et al. (1997) have assessed the impacts of the same global change scenarios used here on wheat cropping for this location thus enabling some preliminary assessment of the impacts of global change on landuse. However, choice of landuse will be affected by the relative profitability of different production systems such as dairying, extensive grazing, forage cropping for finishing cattle, and grain production. Prices received for products can be strongly affected by overseas demand and hence are subject to the impact of global climate and other changes. In the absence of a global economic analysis the results of Reyenga et al. and the current study can be compared only in terms of relative change from the current baseline. Such a comparison (Table 3) suggests greater relative advantages to wheat production for all scenarios with the exception of the Warm scenario. Full integration of these landuse change effects would require: 1) spatial modelling of the respective systems with a socio-economic component; 2) consideration of pests and diseases; and 3) calculation of fluxes of greenhouse gases to determine feedback effects on the global changes. 6. Conclusion The major results of this simulation study were the demonstration that: 1) a simple empirical and operational model of pasture growth was able to be modified, with stated caveats, to include current experimental results on the response of C4 native pastures to increased CO2 ; 2) the evaluation of the potential benefit of increased CO2 especially under warmer, wetter conditions will require careful attention to the effects on nitrogen availability; and 3) the impact on increased CO2 and climate change on grazing management strategies such as changes in annual stocking rate, animal production and frequency of pasture burning can be evaluated providing a basis for comparison with alternative land uses. 7. References Agren, G.I., (1994) The interaction between CO2 and plant nutrition: comments on a paper by Coleman, McConnaughay and Bazzaz. Oecologia, 98:239-240. Archer, S., Schimel, D.S. and Holland, E.A. (1995) Mechanisms of shrubland expansion: land use, climate or CO2? Climate Change, 29:91-100. Ash, A.J., Prinsen, J.H., Myles, D.J. and Hendricksen, R.E. (1982) Short-term effects of burning native pasture in spring on herbage and animal production in south-east Queensland. Proceedings of the Australian Society of Animal Production, 14:377-380,. Burrows, W.H., (1995) Greenhouse revisited - land use change from a Queensland perspective. Climate Change Newsletter, 7:6-7. Campbell, B.D., Stafford Smith, D.M. McKeon, G.M., (1997) Elevated CO2 and water supply interactions in grasslands: a pasture and rangelands management perspective. Global Change Biology 3:177-87. 18 Carter, D.R. and Peterson, K.M. (1983) Effects of a CO2-enriched atmosphere on the growth and competitive interaction of a C3 and a C4 grass. Oecologia 58:188-193. Carter, J.O., Flood, N., McKeon, G.M., Peacock, A. and Beswick, A. (1996) Model framework, parameter derivation, model calibration, model validation, model outputs, web technology; Development of a National Drought Alert Strategic Information System Volume 4. Final Report on QPI 20 to Land and Water Resources Research and Development Corporation (May 1996), pp. 42. Chen, D., Coughenour, M.B., Knapp, A.K. and Owensby, C.E. (1993) Mathematical simulation of C4 grass photosynthesis in ambient and elevated CO2. Ecological. Modelling, 73:63-80. Day, K.A., McKeon, G.M. and Carter, J.O. (1997) Evaluating the risks of pasture and land degradation in native pastures in Queensland. Final Project Report, DAQ124A, RIRDC,. Ghannoum, O., von Caemmerer, S., Barlow, E.W.R. and Conroy, J.P. (1997)The effect of CO2 enrichment and irradiance on the growth, morphology and gas exchange of a C3 (Panicum laxum) and a C4 (Panicum antidotale) grass. Australian Journal of Plant Physiology, 24:227-37. Gifford, R.M., (1988) Direct effects of higher carbon dioxide concentrations on vegetation. In Greenhouse: Planning for climate change, G.I. Pearman (ed), pp. 506-519, CSIRO Australia,. Ham, J.M., Owensby, C.E., Coyne P.I. and Brener, D.J. (1996) Fluxes of CO2 and water vapor from a prairie ecosystem exposed to ambient and elevated atmospheric CO2. Agricultural and Forest Meteorology,77:73-93. Hall, W.B. (1996) Near-real time financial assessment of the Queensland wool industry on a regional basis. PhD Thesis, University of Queensland, Brisbane. Hattersley, P.W. (1983) The distribution of C3 and C4 grasses in Australia in relation to climate. Oecologia, 57:113-128,. Hattersley, P.W. and Watson, L. (1992) Diversification of photosynthesis, In Grass evolution and domestication. G.P. Chapman (ed.), pp 38-116, Cambridge University Press, Cambridge. Henderson, S., Hattersley, P., von Caemmerer, S. and Osmond, C.B. (1994) Are C4 pathway plants threatened by global climate change, In Ecological Studies. Vol. 100: Ecophysiology of Photosynthesis, E.-D. Schulz and M. Caldwell (eds), pp. 529-549, Springer-Verlag, New York. Hendricksen, R.E., Rickert, K.G., Ash, A.J., and McKeon, G.M. (1982) Beef production model. Proceedings of the Australian Society of Animal Production, 14: 204-208. Howden, S.M. and Turnpenny, J. (1997) Modelling heat stress and water loss of beef cattle in subtropical Queensland under current climates and climate change. In Modsim ’97 International Congress on Modelling and Simulation Proceedings, 8-11 December, University of Tasmania, Hobart (Eds McDonald, D.A. and McAleer, M.). Modelling and Simulation Society of Australia, Canberra. pp1103-1108,. Hungate, B.A., Chapin, F.S., Zhong, H., Holland, E.A. and Field, C.B. (1997) Stimulation of grassland nitrogen cycling under carbon dioxide enrichment. Oecologia, 109:149-53. 19 Hunt, H.W., Elliot, E.T., Detling, J.K., Morgan, J.A. and Chen, D.X. (1996) Responses of a C3 and a C4 perennial grass to elevated CO2 and temperature under different water regimes, Global Change Biology 2:35-47. Houghton, J.T., Meira Filha, L.G., Callander, B.A., Harris, N., Kattenberg A. and Maskell, K. (eds), (1996) Climate Change 1995: The Science of Climate Change. Contributions of Working Group I to the Second Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge. Kimball, B.A., Mauney, J.R., Nakayama, F.S. and Idso, S.B. (1993) Effects of elevated CO2 and climate variables on plants. Journal of Soil and Water Conservation, 48:9-14. Kirkham, M.B., He, H., Bolger, T.P., Lawlor, D.L. and Kanemasu, E.T. (1991) Leaf photosynthesis and water use of Big Bluestem under elevated carbon dioxide. Crop Science 31:1589-1594. Knapp, A.K., Hamerlynck, E.P. and Owensby, C.E. (1993) Photosynthetic and waters relation responses in elevated CO2 in the C4 grass Andropogon geradii. International Journal of Plant Science, 154: 459-466. Littleboy, M. and McKeon, G.M. (1997) Appendix 2 - Subroutine GRASP: Grass Production Model. Documentation of the Marcoola Version of Subroutine GRASP, In Final Project Report, DAQ124A, for the Rural Industries Research and Development Corporation. McCown, R.L. (1980), The climatic potential for beef cattle production in tropical Australia: Part I. Simulating the annual cycle of liveweight change. Agricultural Systems, 6:303-318,. McKeon, G.M., Day, KA., Howden, S.M., Mott, J.J., Orr, D.M., Scattini, W.J. and Weston, E.J. (1990) Management of pastoral production in northern Australian savannas. Journal of Biogeography, 17:255-72. McKeon, G.M. and Howden, S.M. (1993) Adapting the management of Queensland’s grazing systems to climate change, In Climate Change: Implications for Natural Resource Conservation. S. Burgin (ed.), pp. 123-40, University of Western Sydney, Hawkesbury. Occasional Papers in Biological Sciences No.1. McKeon, G.M., Rickert, K.G., Ash, A.J., Cooksley, D.G. and Scattini, W.J. (1982) Pasture production model. Proceedings of the Australian Society of Animal Production, 14:202-204. Moore, J.L., Howden, S. M., McKeon, G.M., Carter, J.O. and Scanlan, J.C. (1997) A method to evaluate greenhouse gas emissions from sheep grazed rangelands in south west Queensland. In Modsim ’97 International Congress on Modelling and Simulation Proceedings, 8-11 December, University of Tasmania, Hobart (Eds McDonald, D.A. and McAleer, M.). Modelling and Simulation Society of Australia, Canberra. pp137-142. Morgan, J.A., Hunt, H.W., Monz C.A. and Lecain, D.R. (1994) Consequences of growth at two carbon dioxide concentrations and two temperatures for leaf gas exchanges in Pascopyrum smithii (C3) and Bouteloua gracilis (C4). Plant, Cell and Environment, 17:1023-1033. Morison, J.I.L. and Gifford, R.M. (1984) Plant growth and water use with limited water supply in high CO2 concentrations. I. leaf area, water use and transpiration. Australian Journal of Plant Physiology, 11:361-374. 20 Nie, D., He, H., Kirkham, M.B. and Kanemasu, E.T. (1992a) Photosynthesis of a C3 grass and a C4 grass under elevated CO2. Photosynthetica, 26: 189-198. Nie, D., He, H., Kirkham, M.B. and Kanemasu, E.T. (1992b) Canopy photosynthesis and evapotranspiration of rangeland plants under doubled carbon dioxide in closed-top chambers. Agricultural and Forest Meteorology, 61:205-217. Nie, D., Kirkham, M.B., Ballou, L.K., Lawlor, D.J. and Kanemasu, E.T. (1992c) Changes in prairie vegetation under elevated carbon dioxide levels and two soil moisture regimes. Journal of Vegetation Science, 3:673-678. Nie, D., He, H., Kirkham, M.B. and Kanemasu, E.T. (1993) Photosynthesis and water relations of a C4 and a C3 grass under doubled carbon dioxide. In Proceedings of the 17th International Grassland Congress, Vol. 2, pp 1139-1140, Palmerston North, New Zealand. Norman, M.J.T. (1963) Pattern of dry matter and nutrient content changes in native pastures at Katherine, N.T. Australian Journal of Experimental Agriculture and Animal Husbandry, 3:119124. Owensby, C.E., Coyne, P.I., Ham, J.M., Auen L.M. and Knapp, A.K. (1993) Biomass production in a tallgrass prairie ecosystem exposed to ambient and elevated CO2. Ecological Applications 3: 644-653. Parton, W.J., Stewart, J.W.B. and Cole, C.V. (1988) Dynamics of C, N, P and S in grassland soils: a model. Biogeochemistry 5:109-131. Polley, H.W., Johnson, H.B. and Mayeux, H.S. (1994) Increasing CO2: comparative responses of the C4 grass Schizachyrium and grassland invader Prosopis. Ecology, 75:976-988. Poorter, H. (1993) Interspecific variation in the growth response of plants to an elevated ambient CO2 concentration. Vegetatio, 104/105:77-97. Poppi, D.P., Minson. D.J. and Ternouth, J.H. (1981) Studies of cattle and sheep eating leaf and stem fractions of grasses. I. The voluntary intake, digestibility and retention time in the reticulorumen. Australian Journal of Agricultural Research 32:99-108. Read, J.J. and Morgan, J.A. (1996) Growth and partitioning on Pascopyrum smithii (C3) and Bouteloua gracilis (C4) as influenced by carbon dioxide and temperature. Annals of Botany 77:487-496. Reyenga,P.J., Howden, S.M., Meinke, H. and McKeon, G.M. (1997) Global change impacts on wheat cropping in the Burnett Region of Queensland: a simulation approach. In Modsim ’97 International Congress on Modelling and Simulation Proceedings, 8-11 December, University of Tasmania, Hobart (Eds McDonald, D.A. and McAleer, M.). Modelling and Simulation Society of Australia, Canberra. pp149-154. Rickert, K.G. and McKeon, G.M. (1982) Soil water balance model: WATSUP. Proceedings of the Australian Society for Animal Production 14:198-200. 21 Rogers, G.S., Milham, P.J., Gillings, M. and Conroy, J.P. (1996) Sink strength may be the key to growth and nitrogen responses in N-deficient wheat at elevated CO2, Australian Journal of Plant Physiology, 23:253-264. Rogers, H.H., Runion, G.B. and Krupa, S.V. (1994) Plant responses to atmospheric CO2 enrichment with emphasis on roots and the rhizosphere. Environmental Pollution 83:155-189. Sellers, P.J., Bounoua, L., Collatz, G.J., Randall, D.A., Dazlich, D.A., Los, S.O., Berry, J.A., Tucker, C.J., Field, C.B. and Jensen, T.G. (1996) Comparisons of radiative and physiological effects of doubled atmospheric CO2 on climate. Science 271:1402-1406. Seneweera, S.P, Ghannoum, O., and Conroy, J. (1998) High vapour pressure deficits and low soil water availability enhance shoot growth responses of a C4 grass (Panicum coloratum cv. bambatsi) to CO2 enrichment. Australian Journal of Plant Physiology, 25:287-292. Wilsey, B.J., McNaughton, S.J. and Coleman, J.S. (1994) Will increases in atmospheric CO2 affect regrowth following grazing in C4 grasses from tropical grasslands? A test with Sporobolus kentrophyllus. Oecologia, 99:141-144. Woodward, F.I., (1987) Stomatal numbers are sensitive to increases in CO2 from pre-industrial levels. Nature, 327:617-618. 22 3. Impacts on fire dynamics in the mulga woodlands of south-west Queensland. Howden, S.M., Moore, J.L., McKeon, G.M., Reyenga, P.J., Carter, J.O. and Scanlan, J.C. 1. Introduction Increases in the density of shrubs and trees (woody weeds) in grasslands in semi-arid Australia is currently a major threat to sustainable agricultural production due to the reductions in grass growth and hence livestock performance that occur (eg Burrows et al. 1990). However, whilst these invasions reduce grass production and hence wool cuts, they can also result in substantial increases in carbon stores which may become a new source of income for these regions in the eventuality that carbon emission trading arises from the Kyoto Protocol. This may require adaptation of grazing and burning regimes in these regions depending on the management goals. A further complication is that the dynamics of woody weed invasions may themselves be changed by increases in atmospheric CO2 levels and by possible changes in climate. We investigate responses of the mulga (Acacia aneura) woodland ecosystems in south-west Queensland under scenarios of CO2 and climate change in the context of adaptation for differing management goals. We use an existing model of the dynamics of these woodlands (Moore et al. 1997) adapted to simulate responses to varying atmospheric CO2 levels. 2. Methods The model of Moore et al. (1997) is an extension of GRASP (McKeon et al. 1990). GRASP is a model simulating the above-ground yield of a sward dominated by perennial native grasses. GRASP includes a four layer soil water balance and a plant growth model which calculates the processes of run-off, infiltration, drainage, soil evaporation, tree and grass transpiration, pasture growth, consumption and decay, nitrogen uptake, pasture management effects (i.e. stocking rate and pasture burning) and plant density (i.e. perennial grass basal cover). GRASP calculates pasture growth as a function of grass transpiration, radiation interception, temperature, VPD, nitrogen availability and regrowth potential. Evaluation, calibration and validation of the model are described in Day et al. (1997). The density of trees in GRASP, however, is set by the user within each simulation. The model of Moore et al. (1997) adds modules which simulate the population dynamics and growth of mulga in response to changes in environment and management (stocking rate, burning, tree clearing). It also has an emissions module which tracks the flow of carbon and other emissions through the plant and animal components of the ecosystem although it doesn’t currently model the soil carbon components. We have modified this model to simulate responses to high atmospheric CO2 concentrations within both the grass sward and the mulga components. The general impacts of increased CO2 on C3 grasses are understood in terms of photosynthetic and stomatal responses and changes in nitrogen nutrition (eg Wand et al. 1999). Mulga lands in good condition have a large component of C3 grasses and we assume for these simulations 23 that they dominate the sward, acknowledging that many pastures have substantial C4 components. The CO2 response of the grass component was modified following Howden (1999a,b) by changing the transpiration efficiency, radiation use efficiency and nitrogen efficiency of the grasses. In contrast, the responses of woody plants to increased CO2 appear to be more variable than the C3 grass responses with the meta-analysis by Curtis et al. (1998) showing relatively consistent photosynthetic responses (averaging +16% for woody plants in nutrient-stressed conditions) but quite variable stomatal conductance responses (mean -11% but not significant). We thus have used two sensitivity scenarios which vary the response of mulga to high CO2. These are a low response where daily transpiration per unit tree basal area decreases by 11% but where the water-use efficiency increases proportionately, and a high response where transpiration per unit basal area decreases as before but there is an additional increase in water-use efficiency of 16%. We apply this model to a range of feasible global change scenarios which reflect the different uncertainties. Increases in atmospheric CO2 concentration are highly likely due to existing economic and population development pathways and energy and landuse patterns. In the midrange IPCC scenarios, atmospheric CO2 concentrations are likely to double to about 700ppm around the year 2100 (Houghton et al. 1996). Consequent increases in global temperature are consistently forecast whilst changes in rainfall at regional level have considerable uncertainty. We thus construct a cascading hierarchy of scenarios (Table 1). The temperature and rainfall changes in the Hot, Wet and Dry scenarios are implemented by using the historical climate record for Charleville for the period 1885-1995 but adding 3oC equally to maximum, minimum and dewpoint temperature (Hennessy et al. 1999), whilst rainfall changes are implemented as + 10% multipliers on daily rainfall (Howden 1999a). To ensure consistency between all files, pan evaporation was recalculated using a regression derived from the period 1980 to 1995 when the Class A pan measurements were most reliable (Eq’n 1). VP and VPD were recalculated for the scenarios with changing temperature using the equations in McKeon et al. (1998). Table 1: Changes in carbon dioxide levels, temperature and rainfall for each global change scenario Scenario CO2 (ppm) Baseline CO2 Hot Wet Dry 350 700 700 700 700 Temperature (oC) 0 0 +3 +3 +3 Epan = -1.338 + (0.177 *Rad) + (0.238 * VPD) r2 = 0.920 Rainfall (%) 0 0 0 +10 -10 [Eq’n 1] The simulations were run using the site data and parameterisations from Moore et al. (1997). The initial mulga cohort was altered to give a more even age distribution (200stems/ha each at 0.25m and 1m height, 50 stems/ha at 2m, 30 stems/ha at 5m and 5 stems/ha at 10m) and a total basal area of 0.475m2/ha. Treatments were a factorial combination of grazing by fire with two grazing treatments [ungrazed (U) and grazed (G) with sheep stocking rate set for 24 20% utilisation based on pasture availability at 1 June: McKeon et al. 1990] and three fire treatments [never burnt (N), burnt every 6 years (F) and burnt annually (A). Burning occurs at the end of the dry season, if a threshold biomass of 1000kg DM/ha is present]. These treatments were simulated for both the global change and baseline scenarios. All scenarios apart from the Baseline were simulated with both the Low and High CO2 response by the mulga. Data are generally presented as means of the 110 year simulation. Net emissions are expressed in terms of carbon-dioxide equivalent emissions (CO2-e) by using Global Warming Potentials to adjust for the different radiative forcing and lifetimes of gases such as methane and nitrous oxide (Houghton et al. 1996). The difference between final and inital carbon stores was calculated to give the change in carbon store over the simulation. 3. Results There was a substantial trade-off between wool production and net emissions with four clusters of results being dominated by grazing and burning treatment (Fig 1). Variations in global change scenario and/or mulga CO2 response had lesser impacts than the management. The substantial variation of results within a treatment suggests that there are feasible global change scenarios which result in both high wool production and low net emissions/high average carbon store. 2.0 Grazed and burnt Wool (kg/ha) 1.5 Grazed and unburnt 1.0 0.5 Ungrazed and unburnt 0.0 -400 -300 -200 Ungrazed and burnt -100 0 100 Net Emissions (kg CO 2-e/ha/year) Figure 1: Wool production (kg/ha) and net emissions (kg CO2-e/ha/year) for all treatments and scenarios. Doubling CO2 and climate change increased average net emissions for all treatments except UN with GN remaining similar to the Baseline simulation (Table 2). Average net emissions were negative (ie a sink) for GN (-140kg CO2-e/ha/year) and not influenced much by global change scenario. The sink size was doubled in the UN treatments (-250 for the Dry scenario to -340 kg CO2-e/ha/year for the Wet scenario). Climate change in addition to CO2 increase had minor effects on net emissions for UA, UF, GA and GF treatments. All changes in net 25 Table 2: Average net emissions, average carbon store, change in carbon store, average wool production, number of burns and grass production for each management treatment and global change scenario. Treatment Average Net Emissions Average C Store Change in C Store (kg/ha) Wool Burns Grass Growth (kg CO2-e /ha/yr) (kg/ha) (kg/ha/yr) (no.) (kg DM/ha) Ungrazed - Annual Burn (UA) Baseline 11 3436 CO2 30/29 2854/2939 Hot 30/29 2555/2669 Wet 31/29 2735/2858 Dry 22/19 2726/2836 294 -525/-446 -644/-539 -504/-396 -336/-111 0 0 0 0 0 33 51/51 53/53 55/56 44/44 758 1086/1078 1068/1054 1163/1157 919/909 Ungrazed – Burnt every 6 years (UF) Baseline -36 5913 CO2 -12/-18 5405/5697 Hot -13/-18 5891/6132 Wet -13/-18 5672/5871 Dry -9/-13 5488/5775 3285 2138/2536 2228/2503 2289/2603 1798/2102 0 0 0 0 0 11 17/17 15/15 17/17 15/15 622 971/949 901/886 1028/1013 808/789 Ungrazed - Never Burnt (UN) Baseline -273 11726 CO2 -300/-332 12947/13807 Hot -286/-316 12434/13255 Wet -315/-343 13347/14101 Dry -249/-280 11258/12117 17314 19176/21074 18303/20097 20084/21758 16021/17857 0 0 0 0 0 0 0 0 0 0 428 601/574 583/556 626/602 544/518 Grazed - Annual Burn (GA) Baseline 39 CO2 56/56 Hot 53/52 Wet 59/52 Dry 49/49 3586 2939/3003 2658/2741 2698/2788 2693/2765 -56 -677/-685 -618/-614 -830/-315 -649/-648 1.18 1.39/1.38 0.99/0.98 1.13/1.12 0.89/0.88 23 43/44 45/46 51/48 39/39 719 1038/1032 1022/1016 1133/1150 891/886 Grazed - Burnt every 6 years Baseline 3 CO2 31/25 Hot 28/28 Wet 34/33 Dry 16/14 (GF) 5721 5450/5778 5489/5644 5604/5820 5452/5619 2233 1427/1805 1466/1508 1391/1472 1909/1988 1.1 1.66/1.62 1.16/1.15 1.38/1.36 0.98/0.97 10 15/13 14/14 15/15 11/11 588 885/864 848/839 946/941 731/721 10912 11247/11626 11157/11541 11499/11830 10618/11033 0.9 1.25/1.21 0.87/0.84 0.98/0.93 0.78/0.74 0 0 0 0 0 435 605/587 588/571 639/622 537/519 Grazed - Never Burnt (GN) Baseline -146 10079 CO2 -141/-148 10820/11278 Hot -141/-148 10543/10998 Wet -144/-150 11068/11468 Dry -135/-143 9863/10357 26 emissions were closely linked to differences in average carbon store with decreases in average carbon store from the Baseline simulations for all treatments except UN with marginal changes in GN depending on the climate change scenario. However, carbon store trajectories differed significantly between treatments (Figure 2) with some treatments (UA, GA) having a carbon store at the end of the 110 year simulation less than that at the start. Only the UN treatments showed continuing carbon storage at the end of the simulation (Table 2) with significant variation with scenario in the increase in storage from +17000 kgC/ha (Dry) to +21000 kgC/ha (Wet). The GN treatments also maintained carbon storage for a long period into the simulation. 22000 20000 UN 18000 Carbon Store (kg/ha) 16000 GN 14000 12000 10000 8000 GF 6000 4000 2000 GA 0 1885 1895 1905 1915 1925 1935 1945 1955 1965 1975 1985 1995 Year Figure 2: System carbon stores (kg/ha) over the 110-year historical simulations for selected treatments under the Baseline scenario. Grass growth was substantially stimulated by the doubling of CO2 (mean 44%) with the response being mediated by both grazing/burning treatment and the response of the mulga to CO2 (Table 3). The grass response was least in the treatments that were never burnt (+34 to +40%) and greatest in those frequently burnt (+47 to +56%). High mulga CO2 response decreased the grass CO2 response from 40 to 34% in the UN treatment. There was no effect in some treatments (UA and GA). Table 3: Response (% change from baseline) of grass production to CO2 under the different management scenarios Treatment Ungrazed – Annual Ungrazed – 6 yearly Ungrazed – Never Grazed – Annual Grazed – 6 yearly Grazed – Never Grass Response Low High 43 42 56 53 40 34 44 44 50 47 39 35 The number of burning events (Table 2) in the Baseline simulations were 10 (GF), 23 (GA), 11 (UF) and 33 (UA). Doubling CO2 increased these markedly (14, 43, 17 and 51 respectively). The Hot and the Dry climate change scenarios reduced the number of fire events but they always remained above baseline values. 27 Wool production varied by 30% for the Baseline scenario with values of 1.18, 1.1 and 0.9 kg/ha/year for the GA, GF and GN treatments (Table 2). Doubling of CO2 increased production markedly especially for the GF(50%) and GN (38%) treatments. However, when climate change was simulated in conjunction with CO2 increase, wool production was reduced in all scenarios sometimes to below the Baseline values (eg 30% reduction in the Dry scenario of GA). The differences between the two CO2 responses for the mulga component was small for the average carbon store, average net emissions and the other variables (eg wool production, burning events). 4. Discussion These simulations suggest that this system can be managed to act as either a net source or a net sink of greenhouse gases under current climate and CO2 and under a range of global change scenarios. The key component in determining source or sink status is the management of the woody mulga. Excluding both burning and grazing is the most effective means of increasing carbon stores and hence reducing net emissions although we note that permanent exclusion of fire is probably not a practical option. Under a zero-grazing scenario, this land would no longer be used for agricultural production. If grazing is undertaken with various fire management regimes, there remains a significant trade-off between wool production and increasing carbon stores although there are combinations of management regimes such as excluding fire with light grazing which, on average can provide both. However, it was only in the simulations where no fires or grazing occurred that there a continuing increment of carbon throughout the 110 year simulation. Under other management regimes, initial increases in carbon stores are followed by reductions as the initial mulga cohorts die out. The model is of course a simplification of real management practices and there are opportunities which generally exclude grazing but use the mulga and the associated chenopods as drought reserve (eg Lauder and Freudenberger 1999) which could make multiple uses feasible. The issue of whether carbon sequestration is an alternative landuse to grazing is currently hotly debated. The purchase cost of land, maintenance costs (eg firebreaks and fencing), income foregone, security of store, the costs, accuracy and precision of monitoring and verification, transaction costs and other possible associated benefits and costs (eg biodiversity) all need to be addressed. The low cost of land and current low returns from wool production from the rangelands in south-west Queensland suggests that this region may be suitable to develop a mosaic of landuses which incorporates carbon storage as one goal provided that some of the above cost issues can be addressed (eg reduce monitoring costs). The simulations in Moore et al. (1997) using this model suggest that both the dynamics of storage and total carbon storage are influenced by the initial mulga stand structure. This needs further investigation as it may have an influence on the viability of carbon storage as a landuse. The effects of increases in CO2 on ecosystem carbon stores were unexpected. Mean growth responses from the literature for doubling of CO2 are 44% for C3 grasses (Wand et al. 1999) and 16% for woody species in nutrient stressed conditions (Curtis and Wang 1998). In these simulations when ungrazed and unburnt, this resulted in a 10-17% increase in mean carbon storage with the variation due to the different mulga CO2 responses investigated. This is consistent with the physiological potential of 16%. When grazed but still unburnt, this 28 changed to 7-11% increase in mean carbon store: considerably lower than the potential. However, under all treatments in which burning occurred, doubling CO2 resulted in a smaller mean carbon store and less carbon stored over the simulation. This occurred because the substantial increases in grass growth with doubling of CO2 (34 to 56%) enabled more fire events to occur and this affected the mulga populations, killing off the establishing cohorts needed to ensure continued carbon accumulation. The mean effect of increased CO2 on grass growth across all treatments is 44%. This is identical with the mean effect of doubling CO2 for C3 grasses found in the meta-analysis of Wand et al. (1999) suggesting that this result may be applicable in other ecosystems where fire has a similar function. The effect of changing the CO2 response of the mulga had surprisingly little impact on the emissions dynamics or other aspects of the ecosystems simulated such as fire frequency or wool production. The high mulga CO2 response slightly reduced the grass CO2 response by increasing competition. Further investigation of the response of woody plants under increased CO2 is needed as we have not explored the full array of possible responses. For example, a scenario with no change in water use from stomatal conductance change but a growth increase due photosynthetic response would result in increased competition with grasses. There remain, however, several sources of uncertainty in these analyses apart from those we explored by using scenarios. For example, the potential burning frequency of 23% when grazed (baseline GA treatment) appears more frequent than seen on actual properties. This difference probably reflects maximum possible fire frequency with biological constraints as opposed to current cultural practice. The frequently burnt scenario (every 6 years) is probably more representative of current pasture management in the region with 10 burning events in the historical record. Burning frequency is very sensitive to woody plant density. In a study of the effects of Eremophilla gilesii on potential fire frequency at Charleville, Carter and Johnston (1986) showed that with grazing at 20% utilisation an increase in E. gilesii canopy cover from 7.5% to 10% reduced the frequency of 1000kg yields from 36% to 12%. When average grass fuel load is close to the threshold required for fire one could expect high model sensitivity. The ungrazed/annual burn treatment gave results consistent with those of Johnston and Carter (1986). In the simulations, we also retain domestic stock on the paddock at all times whereas graziers may spell their paddocks occasionally. Mulga is very sensitive to grazing (eg Brown 1985) and these spelling periods appear to be vital in allowing the saplings to grow to a height where they are no longer susceptible to grazing and burning. For these reasons, these simulations cannot be considered to provide a hindcast trajectory of mulga spread even though as discussed in Moore et al. (1997), they simulate the correct gross response observed in various management regimes. In addition, different plant communities are likely to provide very different interactions between grazing and fire management, particularly those with unpalatable shrub species. Further studies are needed on such systems. We do not simulate changes in soil carbon which may be important under some treatments and scenarios. These lands were likely to have been burnt very regularly as part of aboriginal land management practices resulting in the possibility of a relatively large pool of elemental carbon (‘charcoal’). This pool can be as large as 30% of the total soil carbon pool (Skjemstad et al. 1996). Removal of burning from the system as simulated in some of these treatments could result in long-term reduction in the ‘charcoal’ pool which may offset part of the increase in biomass carbon store. The global change scenarios could also alter both carbon inputs through increasing growth and carbon outputs through increasing decomposition rates with increased temperature and, in some scenarios, increased soil moisture. The balance 29 between these factors needs evaluating through a model such as CENTURY (Parton et al. 1988). 5. Summary • The possibility of trading greenhouse gas emission permits as a result of the Kyoto Protocol has spurred interest in developing land-based sinks for greenhouse gases. Extensive grazing lands which have the potential to develop substantial woody biomass are one obvious candidate for such activities. • However, any such activity needs to take into account the possible impacts on existing grazing and the possible impacts on fire frequency of continuing CO2 buildup in the atmosphere and resultant climate change. We use simulation models to investigate these issues in south-west Queensland. • These simulations suggest that this system can be managed to act as either a net source or a net sink of greenhouse gases under current climate and CO2 and under a range of global change scenarios. • The key component in determining source or sink status is the management of the woody mulga. Excluding both burning and grazing is the most effective means of permanently increasing carbon stores and hence reducing net emissions. There are combinations of management regimes such as excluding fire with light grazing which, on average, allow productive grazing but transient carbon storage. • The effects of increased CO2 on ecosystem carbon stores were unexpected. Carbon stores increased (7-17%) with doubling of CO2 only in those simulations where burning did not occur, but decreased when burnt. This occurred because the substantial increases in grass growth with doubling of CO2 (34 to 56%) enabled more fires, killing off the establishing cohorts needed to ensure continued carbon accumulation. • On average doubling CO2 increased grass growth by 44%, this is identical with mean literature values suggesting that this result may be applicable in other ecosystems where fire has a similar function. • A sensitivity analysis of the CO2 response of mulga showed only minor impacts. We discuss additional uncertainties and shortcomings. • Another report in this series will document for Australia’s forests the likely changes in fire regimes with global change, the impacts these changes may have and adaptation options that could be developed to counter these changes and impacts. 6. References Brown, R.F., (1985) The growth and survival of young mulga (Acacia aneura F. Muell) trees under different levels of grazing. Australian Rangeland Journal, 7:143-48. 30 Burrows, W.H., Carter, J.O., Scanlan J.C. and Anderson, E.R. (1990) Management of savannas for livestock production in north-east Australia: contrasts across the tree-grass continuum, Journal of Biogeography, 17:503-512. Carter, J.O. and Johnston, P.J. (1986) Modelling expected frequencies of fuel loads for fire at Charleville in Western Queensland. In: Proceedings Third Queensland Fire Research Workshop, Gatton, 53-67. Curtis, P.S., and Wang, X. (1998) A meta-analysis of elevated CO2 effects on woody plant mass, and physiology, Oecologia, 113(3):299-313. Day, K.A., McKeon G.M. and Carter, J.O. (1997) Evaluating the risk of pasture and land degradation in native pastures in Queensland. Final Project Report for RIRDC project DAQ124A. Hennessy, K.J., Whetton, P.H., Katzfey, J.J., McGregor, J.L., Jones, R.N., Page, C.M. and Nguyen, K.C. (1998) Fine Resolution Climate Change Scenarios for New South Wales. Annual Report 1997-98. CSIRO Atmospheric Research. Howden, S.M., McKeon, G.M., Walker, L., Carter, J.O., Conroy, J.P, Day, K.A., Hall, W.B., Ash A.J. and Ghannoum, O. (1999a) Global change impacts on native pastures in south-east Queensland, Australia, Environmental Modelling and Software, 14, 307-316. Howden, S.M., McKeon, G.M., Carter, J.O. and Beswick, A. (1999b) Potential global change impacts on C3-C4 grass distribution in eastern Australian rangelands. In Proceedings of the VI International Rangeland Congress, pp 41-43, Townsville, Australia. Houghton, J.T., Meira Filho, L.G., Callander, B.A., Harris, N., Kattenberg, A. and Maskell K.M. (eds), (1996) Climate Change 1995. The Science of Climate Change, IPCC. Johnston, P.J. and Carter, J.O. (1986) The role of fire in production systems in western Queensland: a simulation approach. In: Proceedings Third Queensland Fire Workshop, Gatton, 161-172. Lauder, A. and Freudenburger, D. (1999) Sustaining a grazing enterprise in south-western Queensland, Australia, In Proceedings of the VI International Rangeland Congress, pp 10371039, Townsville, Australia. McKeon, G.M., Day, K.A., Howden, S.M., Mott, J.J., Orr, D.M., Scattini, W.J. and Weston, E.J. (1990) Northern Australian savannas: management for pastoral production, Journal of Biogeography, 17:355-72. McKeon, G.M., Hall, W.B., Crimp, S.J., Howden, S.M., Stone, R.C., Jones, D.A. (1998) Climate change in Queensland’s grazing lands. I. Approaches and climatic trends, Rangeland Journal 20:151-176. Moore, J.L., Howden, S. M., McKeon, G.M., Carter, J.O. and Scanlan, J.C. (1997) A method to evaluate greenhouse gas emissions from sheep grazed rangelands in south west Queensland. In Modsim ’97 International Congress on Modelling and Simulation 31 Proceedings, 8-11 December, University of Tasmania, Hobart (Eds McDonald, D.A. and McAleer, M.). Modelling and Simulation Society of Australia, Canberra. pp137-142. Parton, W.J., Stewart, J.W.B. and Cole, C.V. (1988) Dynamics of C,N,P and S in grassland soils: a model. Biogeochemistry, 5:109-131. Skjemstad J.O., Clarke, P. Taylor, J.A., Oades, J.M. and McClure, S.G. (1996) The chemistry and nature of protected carbon in soil. Australian Journal of Soil Research, 34:251-271. Wand, S.J.E., Midgley, G.F., Jones, M.H. and Curtis, P.S. (1999) Response of wild C4 and C3 grass (Poaceae) species to elevated atmospheric CO2 concentration: a meta-analytic test of current theories and perceptions, Global Change Biology, 5:723-741. 32 4. Potential impacts on C3-C4 grass distributions in eastern Australian rangelands S.M. Howden, G.M. McKeon, J.O. Carter and A. Beswick 1. Introduction In the rangelands of eastern Australia, livestock productivity is influenced by the proportion of C3 and C4 grasses in the sward (Wilson and Minson 1980). The proportion of C4 grasses (measured as % of native grass species within regional floristic lists) declines strongly with declining spring and summer temperatures (Hattersley 1983). Temperature and other climatic elements are likely to change in the future due to ongoing human activities such as the burning of fossil fuels. Henderson et al. (1994) suggested that global warming of 4oC may result in the 50% C4 isoline moving southwards by about 250km. The latitude where the frequency of C4 species is 90% may move southwards by up to 500km in eastern Australia. Changes in CO2 concentrations affect water use efficiency of grasses and, in the case of C3 species, photosynthetic rates and nitrogen use efficiency (eg Lutze and Gifford 1999) and these could also be expected to impact on the relative distributions of C3 and C4 species. Ehleringer et al. (1997) have assessed implications of altering CO2 on the ‘temperature crossover point’ for quantum yields of C3 and C4 plants based on a simple physiological photosynthesis model and measurements. Their approach indicates a crossover point of about 21oC (mean growing season temperature) for current CO2 levels for NADP-ME C4 subtypes (the dominant subtype in eastern Australia) with this corresponding to observed change in biomass from C3 to C4 grasses in the US (Epstein et al. 1997). However, this approach suggests that this ‘crossover point’ will increase to about 35oC when CO2 rises to 700 ppm (eg about year 2100) implying that the majority of Australia would become dominated by C3 grasses even considering maximum likely temperature increases. Thus in assessing the likely impacts of global change on rangeland vegetation there is a need to reconcile these opposing hypotheses derived from expected temperature and CO2 change. Furthermore, the analyses to date for Australia have focussed on the relative frequency of C3 and C4 species not the likely pasture production which is both the basis of the grazing industries in these regions and ecosystem dominance. However, in Australia there is no comprehensive database of pasture production that would allow such an analysis. We use instead a pasture growth model validated for tropical C4 pastures (GRASP: Day et al. 1997) parameterised to represent three generic functional groups (cool climate C3, warm climate C3 and C4 grasses) to develop a relationship between floristic representation and C3 and C4 functional group biomass production for eastern Australian rangelands. We then evaluate global change scenarios. 2. Methods Growing season (Sept-April) temperatures were calculated from historical data (1958 to 1988) for 31 sites distributed across the eastern Australian rangelands (Fig 1). GRASP was parameterised using mean values from the predominantly C4 pastures across northern 33 Australia (160 site-by-year sets of pasture growth measurements). Whilst there are considerable data to support C4 parameterisations, there are few equivalent data for native C3 grasslands. The temperature response functions of the cool and warm C3 groups were altered to optima of 15-25oC and 18-30oC respectively. The minimum nitrogen content (ie the N% where nitrogen dilution stops growth) was assumed to be 0.88% compared with 0.68% for C4 grasses. We did not change transpiration efficiency nor potential regrowth rates of C3 groups due to lack of information. Average annual biomass for each functional group was simulated using climate data for 1958 to 1988. The C3 functional group with greatest simulated biomass at each site was then used to calculate a growth index (C4/C3 biomass) which was regressed against observed %C4 frequency interpolated from Hattersley (1983). N 0 km 500 Figure 1. The eastern Australian rangelands showing the line where growth index equals 1 for the current climate and CO2 levels (solid line), the mulga lands which are the northernmost extent of grasslands with a significant C3 component in eastern Australia (shaded area) and the existing 50% C4 isoline from Hattersley (1983) (dashed line). The transect in Table 1 is represented by the dotted line with included sites marked (fi) and other sites marked ( ). 34 The simulation studies were a factorial combination of current CO2/doubled CO2 (700ppm) and temperature change of 0 or 3oC applied to both maximum, minimum and dewpoint temperatures. Modifications to GRASP to simulate doubled CO2 response for C4 grasses and for C3 grass transpiration followed Howden et al. (1999) and change in radiation use efficiency and leaf critical nitrogen content for C3 species based on Reyenga et al. (1999). The impact of CO2 increase on minimum nitrogen contents has been rarely studied and hence uncertain.. Wheat and some cultivated grasses show no discernible change in this parameter (eg Conroy and Hocking 1993) but a recent study of a native C3 grass indicated a substantial decline with doubled CO2 (Lutze and Gifford 1999). We thus ran the doubled CO2 scenarios with this value both set to current levels (0.88%) and reduced by 25% so that it was the same as the C4 grasses. 3. Results Mean growing season (Sept-April) temperature was 20.5oC at the sites where 50% of grasses were C4 which is similar to the crossover point based on quantum yields. The location of the isoline where the growth index equals 1 corresponds closely with the northernmost extent in Queensland of the rangelands with a significant C3 component (Fig 1). However, the position of this line is likely to change with C3 parameterisation as more data become available. The percent C4 species in the native flora was related curvilinearly with the growth index (G) C4%=95.59(1-e-2.918G), R2=0.84, P<0.0001). Southwards movements of the two isolines for growth index=1 and 50% C4 frequency occurred under the 2CO2 (~150km), 2CO2+3oC (~250km) and 3oC (~100km) scenarios and there was a corresponding increase in C4% in the southern regions especially under the scenarios with temperature increase (Table 1). In contrast, in the scenarios in which increased CO2 was assumed to reduce the minimum nitrogen content, there was a northwards movement of the growth index isoline (~250-300km) but the movement of the 50% C4 isoline varied with scenario, being slightly (~100km) northward under the 2CO2 scenario but substantially southward under the 2CO2+3oC scenario. Table 1 Growth index values for seven sites across the transect (Figure 1) for scenarios with factorial combination of +- CO2 increase and + 3oC increase. The two scenarios with CO2 change are repeated but with the C3 functional types minimum N content reduced from 0.88% to 0.68% (LN). Simulated % frequency of C4 values are in parentheses. Site CO2 2CO2 Horsham Balranald Bourke Cunnamulla Charleville Roma Emerald 0.23 (46) 0.33 (59) 0.50 (73) 0.54 (76) 0.87 (88) 1.07 (91) 1.27 (93) 0.27 (52) 0.35 (61) 0.54 (76) 0.60 (79) 0.96 (90) 1.16 (92) 1.27 (93) 2CO2 LN 0.21 (44) 0.27 (52) 0.42 (67) 0.47 (72) 0.77 (86) 0.90 (89) 0.99 (90) +3oC 0.41 (67) 0.43 (69) 0.68 (82) 0.66 (82) 0.94 (89) 1.10 (92) 1.28 (93) 2CO2 +3oC 2CO2 +3oC LN 0.48 (72) 0.37 (63) 0.48 (72) 0.37 (63) 0.67 (82) 0.55 (76) 0.72 (84) 0.57 (78) 1.01 (91) 0.81 (87) 1.18 (93) 0.91 (89) 1.28 (93) 1.01 (91) 35 4. Discussion The 50% C4 isoline occurs close to where it is predicted on the basis of relative quantum yields for current CO2 concentrations. However, the quantum yield approach does not incorporate effects caused by seasonality of rainfall nor nitrogen dynamics which we hypothesise result in significant C3 representation in communities occurring far north of this point. We used a simulation approach to incorporate these factors in calculating a growth index. The observed northern boundary of where C3 grasses contribute substantially to native pasture production corresponds to a line with a growth index of 1, suggesting a bioclimatic component to current distributions. Major differences in soil-species-grazing management associations occur in this region and hence possible bioclimatic effects may be overridden by these non-climatic factors. Improvement of C3 parameterisation is required before further direct comparison of growth of functional types can be made. Global change scenarios suggest possible changes to this simulated ‘boundary’ and to frequency of C3/C4 functional group representation south of this. Temperature increases of 3oC, particularly under the doubled CO2 scenario, generally moved southward both the line where the growth index equals 1 and the 50% C4 line consistent with the changes presented in Henderson et al. (1994). The effects of CO2 increases alone were dependent upon the parameterisation of nitrogen dynamics in the grasses. If no change was made to the minimum nitrogen content (eg Conroy and Hocking 1993), then these lines moved southwards whilst if minimum nitrogen content was reduced (eg Lutze and Gifford 1999) then the lines moved northwards substantially. This occurred because increased N dilution enabled expression of the enhanced photosynthetic and water use efficiency by C3 groups under higher levels of CO2. Under the doubled CO2 and temperature increase scenario with reduced minimum nitrogen content, there was both a small movement north of the growth index line but a southwards movement of the 50% C4 isoline. These uncertainties suggest improved understanding is needed of the nitrogen dynamics of native grasses under global change, particularly in quantifying the minimum nitrogen content which is critical to biomass accumulation in these infertile semi-arid rangelands. However, none of the doubled CO2 scenarios studies resulted in the major changes in distributions suggested by the quantum yield analysis (Ehleringer et al. 1996). 5. Acknowledgements We would like to thank Dr Peter Johnston for discussions on the distribution of grasses in Queensland 6. References Conroy, J. and Hocking, P.J. (1993) Nitrogen nutrition of C3 plants at elevated atmospheric CO2 concentrations. Physiologia Plantarum, 89:570-576. Day, K.A., McKeon, G.M. and Carter, J.O. (1997) Evaluating the risks of pasture and land degradation in native pasture in Queensland. Final report for Rural Industries Research and Development Corporation. Project DAQ124A. 36 Ehleringer, J.R., Cerling, T.E. & Helliker, B.R. (1997) C4 photosynthesis, atmospheric CO2, and climate. Oecologia, 112, 285-299. Epstein, H.E., Lauenroth, W.K., Burke, I.C. and Coffin, D.P. (1997) Productivity patterns of C3 and C4 functional types in the US Great Plains. Ecology 78, 722-731. Hattersley, P.W. (1983) The distribution of C3 and C4 grasses in Australia in relation to climate. Oecologia, 57, 113-128. Henderson, S., Hattersley, P., von Caemmerer, S. & Osmond, C.B. (1994) Are C4 pathway plants threatened by global climatic change ? In: Schultz, E-D., Caldwell, M. (eds) Ecophysiology of photosynthesis. Ecological studies, Vol 100, pp 529-549. Springer-Verlag, New York. Howden, S.M., McKeon, G.M., Walker, L., Carter, J.O., Conroy, J.P., Day, K.A., Hall W.B., Ash, A.J.,& Ghannoum, 0. (1999). Global change impacts on native pastures in south-east Queensland, Australia. Environmental Modelling and Software, 14:307-316. Lutze, J.L. & Gifford, R.M. (1999) Nitrogen accumulation and distribution in Danthonia richardsonii swards in response to CO2 and nitrogen supply over four years of growth. Global Change Biology, (in press) Reyenga, P.J., Howden, S.M., Meinke, H. and McKeon, G.M. (1999) Modelling global change impacts on wheat cropping in south-east Queensland, Australia. Environmental Modelling and Software, 14:297-306. Wilson, J.R. & Minson, D.J. (1980) Prospects for improving the digestibility and intake of tropical grasses. Tropical Grasslands, 14: 253-259. 37 5. Climate change impacts on heat stress and water requirements of cattle in Australia S.M. Howden, J.R. Turnpenny W.B. Hall and D. Bruget 1. Introduction Heat stress is a significant issue for the productivity of livestock grazing in tropical areas of Australia (e.g. Petty et al. 1998) and these stresses are likely to increase with prospects of global warming due to the accelerating emission of greenhouse gases (e.g. Houghton et al. 1996). Heat stress in livestock has been reported to decrease conception rates and increase foetal and postnatal mortalities (Finch 1983; Berman 1991), impair spermatogenesis (Entwhistle 1992), increase urinary nitrogen loss (O’Kelly 1988) and increase susceptibility to a range of parasitic and non-parasitic diseases (e.g. Finch 1983). Heat stress has also been found to decrease growth rates (e.g. Frisch 1981, Hahn 1985) although there appears to be strong compensatory growth responses when animals are returned to less stressful conditions following drought (G. McKeon unpublished data). High heat loads also result in greater water requirements for livestock (e.g. King 1983). Lack of availability of water due to the large distances from water sources that tropical animals sometimes have to forage, can reduce production directly through reductions in metabolic rates and feed intake (e.g. Utley 1970), and indirectly by reducing the area grazed through restriction of distance travelled from watering points (e.g. Noble 1975). Heat stress in cattle has been analysed using two main approaches. Firstly, through the use of relationships such as the Temperature-Humidity Index (THI; Johnson et al. 1963) which relates stress to both daily maximum temperature and dewpoint temperature. This relationship, originally derived for dairy cattle with high intake and metabolic rates, has been shown to be a robust predictor of heat stress in cattle, being related to reduced liveweight gain in beef cattle (Petty et al. 1998), milk production in dairy cattle (Hahn and Oosburn 1969), conception rates (Hahn 1981) and mortality rates (Hahn 1985) and distribution of beef cattle varieties in Africa (King 1983). The THI has also been used operationally for heat stress assessment in dairy cattle in South Africa (Du Preez et al. 1990) and the USA (Hahn and Mader 1997). The THI has minimal input requirements and has been used in a variety of environments, suggesting it is suitable for the broad scale assessment of issues such as climate change impacts. However, King (1993) and Finch (1983) suggest that effective evaluation of the implications of heat stress for beef cattle in extensive grazing systems requires additional measurements of net radiation load and convection. Furthermore, the THI approach does not explicitly incorporate important factors such as coat colour which influence heat loads and thus provides little information on how to direct selection programs to enhance heat loss mechanisms (Finch 1983). An alternative approach is to explicitly model fluxes of water and energy from animals using a physically-based approach such as that in Turnpenny et al. (1997). However, this approach requires considerable parameterisation information and intensive data input (i.e. hourly meteorological variables) which are often unavailable. The second approach uses physically-based models to simulate evaporation from the skin surface of livestock using an energy balance model of the animal. This provides the 38 possibility of modelling water requirements of livestock by using a mass balance approach which accounts for other water gains (e.g. in feed) and losses (e.g. urine). The aim of this study was to compare the two approaches to analysing heat stress on livestock, to develop an approach for modelling water requirements of livestock in the Australian tropics, and to investigate how both heat stress and water requirements are likely to be affected under plausible climate change scenarios. 2. Heat stress model descriptions The THI is calculated as: THI = Tmax + 0.36Tdewpoint + 41.2 where Tmax is daily maximum dry bulb temperature (oC) and Tdewpoint is dewpoint temperature (oC) A series of mathematical models have been developed to predict the metabolic rate and occurrence of thermal stress in different livestock species (Turnpenny 1997). These models calculate heat loss from model animals using hourly meteorological data as input and assess the degree of thermal stress under different combinations of weather conditions. The models were designed to be applicable to as wide a range of conditions as possible, and as such have been based on animal physiology and the physics of heat transfer rather than empirical relationships. The model for beef cattle was developed as a system of cylinders with rounded ends, and incorporates three layers - the underlying tissue, the coat and the external environment. By specifying the thermal environment, metabolic rate and weight of the animal, the temperatures of the layer interfaces are calculated by solving the energy balance at each interface. This allows the heat loss from the animal to be calculated. Physiological responses to heat and cold stress, including sweating and varying blood flow to the peripheries have been parameterised. Further details of the general energy balance model are in Turnpenny et al. (1997), and a more in-depth discussion of considerations for outdoor animals, such as shade and shelter requirements can be found in Turnpenny (1997). The beef cattle model is described briefly in Parsons et al. (submitted). The model requires hourly data for the environmental inputs which we generated by downscaling daily climate data using the approach of Turnpenny (1997). Briefly, hourly values of the following variables: cloud cover fraction, direct and diffuse components of solar radiation, air temperature, radiant temperature of the sky, ground temperature and precipitation were calculated from daily observations of temperature, rainfall, vapour pressure and incident solar radiation. Solar radiation is generated by comparing the top of the atmosphere total radiation with the measured total, and deducing an atmospheric transmittance which is related to cloud cover fraction. The direct and diffuse components for each hour are then calculated empirically. Air temperature is a sine function between the maximum and minimum, with the maximum occurring at 2 pm, and the minimum at 2 am. Sky temperature is an empirical function of cloud cover and air temperature while ground temperature is found by solving a simple energy balance. Vapour pressure is assumed constant over the day. Wind speed data were not available, so a typical 39 value of 2 m/s was used for all times. Precipitation was distributed using a triangular function centred on midday. Brahman (Bos indicus) cattle are now widely used throughout the Australian tropics. Their efficient sweating responses providing superior thermoregulatory ability over those of Bos taurus breeds (Finch et al. 1982) for which the heat loss model was originally parameterised. The model was re-parameterised for Brahman cattle using information in Finch et al. (1982,1984). The simulations reported here were for a 425 kg animal. Key parameter changes for these simulations were thermoneutral heat production of 600W, coat albedo 0.5 and 15 mm coat depth on the body and 3 mm on head and legs. 3. Water requirement model Cattle have a very efficient sweating mechanism which increases heat loss when the conditions are too hot for the metabolic heat production and heat gain from the environment to be dissipated by sensible heat loss alone. Under hot conditions, a 500 kg cow can lose up to one litre per hour through sweating alone (Thompson 1973, Webster 1974). The thermal balance model described above calculates the evaporative water flux (E) from the skin given the environmental conditions. This flux can then be combined with estimates of water gain from metabolic water production (M), water in the feed (F) and water in the urine (U) and faeces (S) in a mass balance approach to calculate water intake: Water intake (l/day) = E + U + S - M - F F depends on the feed intake per unit time, and the water content of the forage which can vary from 0% to more than 80% of dry matter (DM) depending on the growing conditions and species (Wood et al. 1996). In this study, the water content of the forage was assumed to be a constant 35% with an intake of 12 kg DM/day (for the 425 kg animal modelled here). M was modelled as a linear function of the total metabolic heat production (600 W for a 425 kg animal) following Brown and Lynch (1972). The fraction of water intake lost as urine and faeces remained constant at 32% and 35% respectively following the results of Colditz and Kellaway (1972) for Brahman cattle in tropical Australia under a range of temperature conditions. Additional water is needed if there is a high salt concentration in the diet, however, this is generally not a major issue in tropical Australia unlike the saltbush (Atriplex spp.) shrublands of temperate Australia. 4. Single site modelling study The two approaches for calculating heat stress were tested for one site (Gayndah, Qld.,25.7oS, 151.8oE) in subtropical Australia for the forty years 1957-96. Daily climate data for maximum and minimum air temperatures, rainfall, evaporation, total solar radiation on a horizontal surface and vapour pressure (HPa) were retrieved from climate surfaces (Carter et al. 1996) and then downscaled to hourly data for the physically-based approach. Dewpoint was calculated following McKeon et al. (1998) as: 40 Tdewpoint = (237.3*ln(VP/6.107))/(17.269-ln(VP/6.107)) To enable comparison with the THI, a daily heat stress index (HSI) was constructed from the physically-based model by averaging the calculated ratio between hourly evaporative flux density needed to maintain homeostasis and the maximum evaporative flux for the five hours from 11am to 3pm. Daily modelled water intake was calculated using the mass-balance approach described earlier for the years 1957 (driest year of the record - 220mm) and 1959 (wettest year - 945mm). These years were chosen as they represent the extremes available in this 40 year record. The THI was compared with the stress index from the physically-based model for all years and the calculated water intake was compared with both indices. Thermal stress is likely to occur in beef cattle when THI exceeds a threshold value of 79-80 (Hahn and Mader 1997, D. Mayer pers. comm.) and days where this occurred are subsequently termed ‘heat stress days’. Regression analysis was used to determine if there was any trend of change in the frequency of heat stress days during the past 40 years. A climate change scenario (2xCO2) was constructed based on the Australian 1996 CSIRO scenarios (http:/www.dar.csiro.au/pub/programs/climod/cm4.htm) for a doubling of atmospheric CO2 concentration for the mid-range emissions scenarios and mid-range climate sensitivities. This suggested a 2.76oC increase in temperatures in this location happening in around the year 2100. The historical temperature record was modified by increasing both the daily maximum and minimum temperatures and then recalculating the hourly data. The climate change scenarios also suggest rainfall changes (summer -24% to 0% and winter -12% to +12%), however these were not included in these simulations as they are likely to have minor secondary impacts compared with the temperature and associated changes. 5. National modelling study Daily climate data for maximum air temperatures and vapour pressure (VP; HPa) were retrieved from climate surfaces (Carter et al. 1996) to give values on a 5km grid across Australia for the period 1957 to 1997. Dewpoint was calculated as before. We recorded the frequency of days (%) above the THI threshold of 80 for each year to produce a map of mean frequencies across the historical period. For each grid cell, the frequency of heat stress days was regressed against year from 1957 to 1997 to determine if there was a trend over time. A climate change scenario was constructed as before and this suggested a general 2.7oC increase in temperatures for the year 2100. We have not varied this by region as this would have resulted in geographic discontinuities in model response. The historical temperature record was modified by increasing both the daily maximum and dewpoint temperatures by 2.7oC. The frequency of heat stress days using the climate change scenario record was then recalculated and mapped. 6. Results 41 Site-based analysis Simulated daily water requirements were highly correlated with daily mean temperatures for both the wettest and driest years on the climate record used for Gayndah (e.g. Figure 1). There was no significant difference between the regressions (p <0.001) for the two years with a pooled regression being: Water requirement (l/day) = 1.30 Tmean - 1.59 Simulated water requirements using this regression were 20.5, 29.6, 40.0 and 47.8 litres/day for 17oC, 24oC, 32oC and 38oC respectively. Measured values of water requirements for Brahman cattle in controlled temperature rooms are 30, 36, 38 and 44-49 litres/day for the above temperatures (Colditz and Kellaway 1972, O’Kelly and Reich 1981) indicating good agreement at temperatures of 32oC or higher. Water requirements (l/day) 40 35 y = 1.2895x - 1.4125 R2 = 0.9812 30 25 20 15 10 5 0 0 10 20 o Temperature ( C) 30 40 Figure 1. Variation of simulated water requirements (l/day) with daily mean temperature (oC) for Gayndah for the year 1957. THI There were strong linear relationships between the daily THI values and the HSI (Figure 2: r2= 0.84) and between simulated water requirements and THI (Figure 3: r2 = 0.96). The relationship between water requirements and the HSI (Figure 4) was less strong (r2 = 0.74) with significant variation due to other factors such as solar radiation, evaporation and vapour pressure. 100 90 80 70 60 50 40 30 20 10 0 y = 53.696x + 34.958 R2 = 0.8369 0 0.2 0.4 0.6 0.8 Stress Index 1 1.2 Figure 2: Relationship between daily THI and the daily stress index from the physicallybased model. 42 Water requirements (l/day) 45 40 35 30 25 20 y = 50.583x - 11.37 R2 = 0.7382 15 10 5 0 0 0.5 1 Stress Index 1.5 Water requirements (l/day) Figure 3: Relationship between simulated daily water requirements and the daily stress index from the physically-based model. 45 40 y = 0.9803x - 47.116 R2 = 0.9553 35 30 25 20 15 10 5 0 0 20 40 60 80 100 THI Figure 4: Relationship between simulated daily water requirements and daily THI. The incidence of days that exceeded a THI threshold value of 80 has increased significantly (P < 0.01) during the past 40 years (Figure 5) with the regression being: Frequency (days/year) = 0.7028*year - 1331 (r2= 0.17) Inspection of the data showed that high frequencies of days exceeding the threshold occurred in drought years. Frequency of THI stress (days/year) 120 100 y = 0.7028x - 1330.6 R2 = 0.169 80 60 40 20 0 1950 1960 1970 1980 1990 2000 Year Figure 5: Annual frequency of days when THI greater than 80 for the years 1957-1996. Median water requirements were increased by about 13% under the climate change scenario compared with current conditions (Figure 6) whilst this increase was only 7% for the HSI 43 (Figure 7) and 5% for the THI (Figure 8). However, in the climate change scenario there were 92 days where the HSI exceeded a value of 1 whereas there were no days which this occurred in the current climate scenario. Heat stress index values greater than unity mean that the cattle can no longer thermoregulate via sweating alone and that they need to either start to pant or adopt behavioural changes (i.e. seek shade or stand head-on to the sun) to avoid hyperthermia. In contrast the highest HSI value for the climate record 1957-1997 was 0.92. Probability of exceedence (%) The THI threshold value of 80 was exceeded on only 16% of days under the current climate. Under the climate schange scenario this occurred on 38% of days. 100 90 80 70 60 50 40 30 20 10 0 1xCO2 2xCO2 0 10 20 30 40 Water requirements (l/day) 50 Probability of exceedence (%) Figure 6: Probabilities of exceedence of water requirements (l/day) under the current climate and with the climate change scenario. 100 90 80 70 60 50 40 30 20 10 0 1xCO2 2xCO2 0 0.5 1 1.5 Heat Stress Index Probability of exceedence (%) Figure 7: Probabilities of exceedence of the heat stress index under the current climate and with the climate change scenario. 100 90 80 70 60 50 40 30 20 10 0 1xCO2 2xCO2 0 20 40 60 80 100 THI Figure 8: Probabilities of exceedence of the THI under the current climate and with the climate change scenario. National analysis 44 The current mean frequency of heat stress days decreases substantially north (70-80% of days) to south (0-10%; Figure 9) with a broad band of low heat stress incidence along the southern and eastern coastlines. Under the climate change scenario, there is a marked increase in mean frequency of heat stress days in most locations with an increase of 10-20 percentage points occurring across most of the Australian rangelands and with greater increases (about 30 percentage points) in the Northern Territory and northern Queensland (Figure 10). Figure 9: Average frequency of heat stress days 1957-1998 Figure 10: Average frequency of heat stress days: 2.7oC climate change scenario 45 The frequency of days in which THI exceeded the threshold value of 80 has increased significantly since 1957 for large areas of northern, central and eastern Australia (Figure 11). However, in a large part of West Australia and southeast Australia there has been little or no increase with declines in some regions. Figure 11: Change in frequency of heat stress days (%/year) 1957-1998 7. Discussion Mean frequencies of heat stress days currently vary markedly across Australia with frequencies of 70-80% occurring at the top of the Northern Territory and 0-10% in a broad band across coastal southern and southeast Australia. Under the climate change scenario used here, the pattern of heat stress shifts generally southwards with frequencies of 90-100% experienced in the top of the Northern Territory and western Cape York and the band where frequencies are 0-10% shrinks markedly to encompass small areas in southern Western Australia and southeast Australia. Generally there is an increase of about 10 to 20 percentage points in heat stress frequencies although in some locations, such as Arnhem Land in the Northern Territory, the increase is greater. The incidence of heat stress as measured by the frequency of days with THI greater than 80 has increased significantly over the 40 year record at both the Gayndah site and more generally over large areas of Australia although there are also regions where there has been no change through to a small decline. This increase is consistent with recorded increases in maximum temperatures (e.g. Wright et al. 1996) and relative humidity (McKeon et al. 1998) and also the incidence of drought as drought years appear to be also associated with high stress frequencies. The changes experienced at Gayndah over the past 40 years (approximately 60% increase over the period) are proportionately similar to those suggested over the next 100 years in the climate change scenario (138% increase compared with the mean of the 1957-97 period). We found a strong correspondence between the empirical THI and the stress index constructed from the physically-based model using the ratio of energy loss from evaporative 46 transfer needed to maintain homeostasis against the maximum possible loss under those environmental conditions. This suggests that studies of heat stress in cattle using the more empirical THI model remain useful indicators of animal stress even though they use much lower levels of data input. However, the physically-based model used here provides an additional capability to investigate specific situations where the data is available. The physically-based model simulates water requirements of Brahman cattle effectively at temperatures at or above 32oC but tends to underpredict them at lower temperatures when compared with measured water needs in controlled environment rooms (Colditz and Kellaway 1972, O’Kelly and Reich 1981). However, total heat loads in such rooms are not necessarily equivalent to those experienced in the field at the same nominal temperature (e.g. Finch 1983). Finch suggests that the heat exchanges are approximately equivalent in the two environments in the range of 35 to 45oC, suggesting that the model is performing well but that further testing is required at lower temperatures. Strong linear relationships were found between modelled water requirements and both temperature and THI as we would expect. This suggests that relationships similar to those of King (1983) between water requirements and readily available climate data could be developed for broad scale studies. In contrast, the relationship between water requirements and the stress index derived from the physically-based model was less robust with factors such as solar radiation, precipitation and vapour pressure adding variability. Simulated water requirements at Gayndah increased significantly (~13%) under the climate change scenario used here when compared with current conditions suggesting that any overgrazing near watering points is likely to be exacerbated under global change. Hence, the smaller median increases in the two indices (5% for THI and 7% for the stress index) understates the potential significance of the climate change on animal stress. Additionally, in the current climate, the animal temperature regulation was achieved by sweating alone on all days, with the maximum ratio of evaporative flux density being 0.92. However, under climate change, the value of this index exceeded unity on 92 days in the record suggesting that physiological and behavioral change would be needed by animals to avoid hyperthermia. As found more generally, climate change at Gayndah is likely to increase frequency of heat stress days with the current frequency of 16% of days more than doubling (to 38%) under the climate change scenario used here. These results suggest that past selection for cattle lines with effective thermoregulatory control (e.g. Hammond et al. 1996) are likely to need to be continued into the future if livestock productivity is to be maintained across northern Australia. However, Finch et al. (1982) found that thermoregulatory control was negatively correlated with metabolic rate suggesting that it may be difficult to combine the desirable traits of adaptation to high temperature environments with high production potential in cattle, although there are possibilities for selection of coat colour and characteristics that may provide some opportunities for selection of heat tolerance Finch et al. 1984). A physically-based model such as that used here could be useful in determining the significance of different sets of thermoregulatory mechanisms for selection programs. There may also be a need on a continuing basis to proactively research and implement management strategies to offset increased heat stress (e.g. Hahn and Mader 1997, Blackshaw and Blackshaw 1994) The current work provides a simple assessment of the drinking requirements of cattle in tropical environments. However, some of the assumptions could be improved, for example, 47 the parameterisations of metabolic water production and water from feed intake. The inclusion of wind run data is also needed to determine the effect this variable may have. Variations in feed intake and feed quality which occur in the field could be accounted for by linking the model to existing or new diet selection or feed intake models. The hourly weather data could be improved, either by using measured hourly data or using more complicated statistical techniques to downscale variables like rain. At present, the rainfall scheme may lessen the impact of high solar radiation in the middle of the day or the combination of high winds and rainfall at night. Finally, more data on the physiology of tropical animals such as properties of the coat and tissue insulation are needed to allow a full model of tropical cattle to be built. 8. Acknowledgements We would like to thank the Queensland Department of Natural Resources for supplying the daily weather data and Dr Bob Hunter (CSIRO Animal Production) for background reading materials on the topic. Thanks also to Prof. Ian Noble for discussions and ideas on the water balance models, to Penny Reyenga and Drs Mikhail Entel and Barney Foran for comments on the manuscript and to the UK Ministry of Agriculture, Nottingham University and the Royal Meteorological Society for funding for travel by JT. This work was partly funded by the climate research program of Environment Australia and the Climate Variability in Agriculture Program administered by the Land and Water Resources Research and Development Corporation. 9. REFERENCES Berman, A., (1991) Reproductive responses of ruminants under high temperature conditions, In Animal Husbandry in Warm Climates, Ronchi, B., Nardone, A. and Boyazoglu, J.G. (eds), EAAP Pulbication No. 55, pp 31-38,. Blackshaw, J.K. and Blackshaw, A.W. (1994) Heat stress in cattle and the effect of shade on production and behaviour: a review. Australian Journal of Experimental Agriculture, 34: 285295. Brown, G.D. and Lynch, J.J. (1972) Some aspects of the water balance of sheep at pasture when deprived of drinking water. Australian Journal of Agricultural Research, 23: 669-684,. Carter, J.O., Flood, N.F., Danaher, T., Hugman, P. & Young, R. (1996) Development of data rasters for model inputs, in Development of a National Drought Alert Strategic Information System, Vol. 4, Final Report on QPI 20 to LWRRDC. Colditz, P.J. and Kellaway, R.C. (1972) The effect of diet and heat stress on feed intake, growth, and nitrogen metabolism in Friesian, F1 Brahman x Friesian, and Brahman heifers, Australian Journal of Agricultural Research, 23: 717-725,. Du Preez, J.H., Hattingh, P.J., Giesecke, W.H. and Eisenberg, B.E. (1990) Heat stress in dairy cattle and other livestock under southern African conditions. III. Monthly temperature- 48 humidity index mean values and their significance in the performance of dairy cattle, Onderstepoort Journal of Veterinary Research, 57: 243-248. Entwistle, K. (1992) Effects of heat stress on reproductive functions in bulls, In Proceedings of a workshop on bull fertility, Queensland Department of Primary Industry, Brisbane. Finch, V.A. (1983) Heat as a stress factor in herbivores under tropical conditions, In Proceedings of the American Society of Animal Science, 89-105. Finch, V.A., Benentt, I.L. and Holmes, C.R. (1984) Coat colour in cattle: effect on thermal balance, behaviour and growth, and relationship with coat types, Journal of Agricultural Science, Cambridge, 102: 141-147. Finch, V.A., Benentt, I.L. and Holmes, C.R. (1982) Sweating response in cattle and its relation to rectal temperature, tolerance of sun and metabolic rate, Journal of Agricultural Science, Cambridge, 99: 479-487. Frisch, J.E., (1981) Changes occurring in cattle as a consequence of selection for growth rate in a stressful environment. Journal of Agricultural Science, Cambridge, 96, 23-38. Hahn, G.L. and Oosburn, D.D. (1969) Feasibility of summer environmental control for dairy cattle based on expected production losses. American Society Agricultural Engineers, 12, 448-451. Hahn, G.L. and Mader, T.L. (1997) Heat waves in relation to thermoregulation, feeding behavior and mortality of feedlot cattle. Proceedings, 5th International Livestock Environment Symposium, American Society of Agricultural Engineers, St. Joseph, MI. pp 563-571. Hahn, G.L., (1981)Housing and management to reduce climatic impacts on livestock, Journal of Animal Science, 52: 175-186. Hahn, G.L., (1985) Management and housing of farm animals in hot environments, in Stress Physiology in Livestock. Ungulates, Vol. 2, edited by M.K. Yousef, pp 151-176, Boca Raton, Florida: CRC Press. Hammond, A.C., Olson, T.A, Chase, C.C., Bowers, E.J., Randel, R.D., Murphy, C.N., Vogt, D.W. and Tewolde, A. (1996) Heat tolerance in two tropically adapted Bos taurus breeds, Senepol and Romosinuano, compared with Brahman, Angus and hereford cattle in Florida. Journal Animal Science, 74: 295-303. Houghton, J.T., L.G. Meira Filho, B.A. Callander, N. Harris, A. Kattenberg and K. Maskell (eds), Climate Change 1995. The Science of Climate Change, IPCC 1996. Johnson, H.D., Ragsdale, A.C., Berry L.L. and Shanklin, M.D. (1963) Temperature-humidity effects including influence of acclimation in feed and water consumption of Brahman, Santa Gertrudis and Shorthorn calves during growth. Missouri Agricultural Experimental Station Research Bulletin No. 846. 49 King, J.M. (1983) Livestock water needs in pastoral Africa in relation to climate and forage. Addis Ababa, ILCA Research Report No. 7. McKeon, G.M., Hall, W.B., Crimp S.J., Howden, S.M., Stone, R.C. and Jones, D.A. (1998) Climate Change in Queensland’s grazing lands: . Approaches and Climatic Trends. The Rangeland Journal, 20: 151-176. McKeon, G.M., Howden, S.M., Abel, N.O.J. and King, J.M. (1993) Climate change: adapting tropical and subtropical grasslands, in Proceedings of the XVII International Grassland Congress, Palmerston NZ, 13-16 February 1993, Vol 2, pp 1181-1190. Noble, I.R. (1975) Computer simulations of sheep grazing in the arid zone. PhD Thesis, University of Adelaide. O’Kelly, J.C. and Reich, H.P. (1981) Sebum output and water metabolism in different genotypes of cattle in hot environments, Journal of Thermal Biology, 6: 97-101. O’Kelly, J.C. (1988) Effects of heat on cattle, In Animal Production in Australia, Proceeding of the Australian Society of Animal Production, Vol 17, Sydney, May 1988, Permagon Press. Parsons, D.J., Cooper, K., Armstrong, A.C., Matthews, A.M., Turnpenny, J.R., Clark, J.A. and McArthur, A.J. (submitted) Integrated models of livestock systems for climate change studies. 1. Grazing Systems. Global Change Biology. Petty, S.R., Poppi, D.P. and Triglone, T. (1998) Effect of maize supplementation, seasonal temperature and humidity on the liveweight gain of steers grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia. Journal Agricultural Science, 130: 95-105. Thompson, G.E. (1973) Review of the progress of dairy science - climatic physiology of cattle. Journal of Dairy Research, 40: 441-473. Turnpenny, J.R. (1997) The impact of climate change on the thermal balance of UK livestock. PhD Thesis, University of Nottingham. Turnpenny, J.R., Clark, J.A. and McArthur, A.J. (1997) Modelling the effect of environmental conditions on the thermal balance of the ewe. In Proceedings of the Fifth International Livestock Environment Symposium, Minneapolis, May 29-31 1997, edited by R.W. Bottcher and S.J.Hoff, pp 234-241. ASAE Publications. Utley, P.R., Bradley, N.W. and Boling, J.A. (1970) Effect of restricting water intake on feed intake, nutrient digestibility and nitrogen metabolism in steers, Journal of Animal Science, 31: 130. Webster, A.J.F. (1974) Heat loss from cattle with particular emphasis on the effects of cold. in Heat Loss from Animals and Man. In Proceedings of the Twentieth Easter School in agricultural science, University of Nottingham, 1973, eds J.L. Monteith and L.E.Mount, pp 205-231, Butterworths, London. 50 Wood, H., Hassett, R., Carter, J., Danaher, T. et al. (1996) Development of a national drought alert strategic information system. Volume 2: Field validation of pasture biomass and tree cover. Final report on QPI 20 to LRRDC. Wright, W. J., de Hoedt, G., Plummer, N., Jones, D. A., and Chitty, S. (1996) Low frequency climate variability over Australia. In Proceedings Second Australian Conference on Agricultural Meterology; October 1996; Uni. of Queensland, Brisbane. pp 102-106. 51 6. Past and future competitiveness of wheat and beef cattle production in Emerald, NE Queensland. Howden, S.M., McKeon, G.M., Reyenga, P.J., Entel, M. Meinke, H. and Flood, N. 1. Introduction Managing climate variability is a key requirement for Australian livestock and cropping industries (eg Meinke and Hammer 1995). Farm management and government policies require analyses based on probabilistic information (eg Stafford Smith and McKeon 1999). These analyses typically use climate information from a 40 to 100 year record. However, recent evaluations suggest that there is long-term variability in our climate record (eg Isdale et al. 1986; Power et al. 1999) and this along with the possible emergence of trends related to climate change and increased atmospheric concentrations of carbon dioxide (CO2) raise issues about the applicability of such evaluations (McKeon et al. 1998). Furthermore, such evaluations typically focus on one industry in isolation whereas shifts between industries can be a rational response to climate variations. For example, the peanut industry further south in Queensland expanded considerably in response to favourable climatic conditions followed by retraction when these conditions reverted (Meinke and Hammer 1995). There is thus a general expectation that agricultural industries at the climatic margin will be most affected by changes in climate means and variability and are thus appropriate locations for study of long-term industry viability. Emerald, north-east Queensland (23o 34’ S, 148o 11’ E) is at the northern margin of the wheat cropping region of Australia. The Emerald region was previously used predominantly for grazing beef cattle, however, cropping has developed in importance over the past 30 years (Figure 1). The trends in climate documented by McKeon et al. (1998) suggest that it is possible that the relative suitability of cropping versus grazing in Emerald is an artifact of recent climate. In addition, increasing concentrations of CO2 in the atmosphere may have had an impact on yields. Expectations of further changes in climate and CO2 concentration suggest that relative productivity of the grazing and cropping landuses will change in the future (Howden et al. 1999b). The implications for industry and policy will be different if change in cropping area is due to long-term variability or to climate change. We use simulation models of grazing systems (GRASP) and wheat cropping systems (I_WHEAT) to assess the relative biological productivities of these two landuses over the last 108 years using a daily climate record and recorded CO2 concentrations. We compare these results with a set of global change scenarios consisting of combinations of CO2 and climate change. 2. Methods 2.1 GRASP description GRASP (McKeon et al. 1990) is a model simulating the above-ground yield of a sward dominated by perennial native grasses. A full description of each equation is given in Littleboy and McKeon (1997). Evaluation, calibration and validation are described in Day et al. (1997) including for the Dichanthium pastures where cropping occurs in Emerald. GRASP 52 Area (ha) was run with a responsive stocking strategy aiming to use 20% of the standing biomass present at the 1st June. 45000 40000 35000 30000 25000 20000 15000 10000 5000 0 1960 1970 1980 1990 2000 Year Figure 1: Area of wheat cropping in the Emerald Statistical Local Area The changes to GRASP to incorporate CO2 effects are extensions of those described in Howden et al. (1999a). Parameterisations of GRASP were largely made through experimentation around 1990 when CO2 levels were about 355ppm and all parameter changes were scaled against this CO2 concentration. Parameters relating to transpiration were altered linearly with CO2 concentration between values of 280ppm (pre-industrial levels) and 700ppm (doubled current levels) using the relationships in Howden et al. (1999a). Similarly radiation use efficiency (RUE) was varied linearly to increase by 5% with CO2 concentration of 710ppm and reduced linearly by 8% at 280ppm. 2.2 I_WHEAT description I_WHEAT is a wheat crop module of the APSIM modelling system which simulates soil moisture, soil carbon:nitrogen dynamics and residue management. A full description of I_WHEAT and model testing is given in Meinke et al. (1998). Changes to I_WHEAT to simulate variable CO2 environments are documented in Reyenga et al. (1999a). Management and soil parameterisations are as in Howden et al. (1999b). I_WHEAT adequately simulates yield from statistical areas (Howden et al. 1999b). Regressions of dewpoint as a function of maximum and minimum temperature (Eq’n 1) were calculated from the post-1957 Emerald record, and pan evaporation was calculated as a function of maximum and minimum temperature and solar radiation (Eq’n 2) from the 19751994 period when Emerald pan values were more likely to be accurate. These relationships were then used to calculate pan evaporation and dewpoint consistently over the whole climate file. GRASP uses dewpoint, assumed to be reached at minimum daily temperature, to calculate vapour pressure deficit. Simulations with measured humidity and pan evaporation against the reconstructed values showed strong correspondence (within 5%) and no difference in average values over 40 years (data not shown). Dewpoint (oC) = 1.45 - 0.0448Tmax + 0.884Tmin (r2=0.727, n=13000) (Eq’n 1) Pan evaporation (mm/day) = -3.60 + 0.126Tmax + 0.101Tmin + 0.219Srad (r2=0.77, n=7304) (Eq’n 2) 53 The long-term CO2 record was constructed from the ice-core data of Etheridge et al. (1998) and more recently from the direct atmospheric measurements at Mauna Loa (Keeling and Whorf 1998). Linear interpolation was used for values for years in which no measurement occurred. 2.3 Climate change and CO2 scenarios Mid-range scenarios of atmospheric CO2 concentrations suggest an increase from the current levels of 364ppm to about 700ppm by the year 2100 (Houghton et al. 1996). This is expected to result in global warming as well as uncertain changes to rainfall characteristics. Global change scenarios were constructed with 1) 700ppm CO2 but with historical climate, 2) 700ppm but with temperature increases of 3oC, 3) both CO2 and temperature increase but with 10% rainfall decrease and 4) both 700ppm CO2 and temperature increase but with 10% rainfall increase. The climate change scenarios were implemented as in Howden et al. (1999a) with pan evaporation recalculated as described previously. 2.4 Data analysis Wheat yields, grass yields and LWG and their coefficients of variation (CV) are presented as running means to allow ready comparison over periods of different lengths. In all cases the mean presented is that of the preceding number of years (eg a running mean of 20 years for 1950 is the mean for the period of 1931 to 1950). To compare relative biological productivities of the cropping and grazing industries we calculated two annual indices by dividing wheat yield by grass production and wheat yield by LWG/ha. We also compare El Niño events and values of the Interdecadal Pacific Oscillation (IPO), an index of decadal and multi-decadal climatic variability derived from the principal component score of the third empirical orthogonal function (EOF3) of a near-global sea-surface temperature analysis, (Power et al. 1999) with the above results. 3. Results The 5-year running mean of wheat yield has been above the mean since the 1960s and was the highest on record during the 1980s but declined towards the overall mean over the past few years (Figure 2). The lowest 5-year running mean yield occurred in 1905 but was similar to several values in the 1930s. The lowest 20-year mean occurred in the late 1930s to 1940s with the greatest value occurring over the last few decades. For grass yield, the lowest 10 and 20-year means were found in the 1930s to 1940s, whilst the highest on record were experienced over the last few decades declining to the mean over the past few years (Figure 3). The lowest 5-year mean value was in 1996 although this was again similar to values in the 1930s to 1940s. 54 3500 4000 (a) 3000 3500 (a) 2500 3000 2000 2500 1500 1000 2000 3500 500 (b) 4000 1500 3000 (b) 3500 2500 3000 2000 2500 1500 1000 500 3500 (c) 3000 2000 1500 4000 3500 (c) 2500 3000 2000 2500 1500 1000 2000 500 (d) 1900 1920 1940 1960 1980 0 .7 5 0 .7 0 0 .6 5 0 .6 0 0 .5 5 0 .5 0 0 .4 5 0 .4 0 0 .3 5 2000 Figure 2: Wheat yield (kg/ha) running means for (a) 5, (b) 10, (c) 20 year periods and (d) 20 year running mean coefficient of variation for Emerald. The bar on the top right indicates the range of mean yields for the year 2100 global change scenarios. 0 .6 1500 (d) 0 .5 0 .4 0 .3 0 .2 1900 1920 1940 1960 1980 0 .1 2000 Figure 3: Grass production (kg/ha) running means for (a) 5, (b) 10, (c) 20 year periods and (d) 20 year running mean coefficient of variation for Emerald. The bar on the top right indicates the range of mean production for the year 2100 global change scenarios. 40 35 (a) 30 25 20 15 40 (b) 40 35 0 .9 35 0 .8 30 0 .7 25 0 .6 20 0 .5 15 0 .4 (c) (a) 100 0 .3 (b) 90 30 80 25 70 20 60 50 0 .5 15 (d) 0 .4 1900 1920 1940 1960 1980 40 2000 0 .3 0 .2 1900 1920 1940 1960 1980 0 .1 2000 Figure 4: LWG (kg/ha/year) running means for (a) 5, (b) 10, (c) 20 year periods and (d) 20 year running mean coefficient of variation for Emerald. The bar indicate the range of mean LWG for the year 2100 global change scenarios. Figure 5: Ratio of (a) wheat yield (kg/ha) to grass production (kg/ha) and (b) wheat yields to LWG/ha for Emerald. Bars indicate the range of mean ratio for the global change scenarios 55 For LWG, the lowest 10 and 20-year running mean values were found in the 1930-40s and highest values over the past decades, although they have been declining over the last several years with the 5-year mean in 1997 being the lowest on record (Figure 4). The 10-year running mean of the ratio of grain:grass production has been above the overall mean since the 1970’s and over the past few years has been the equal highest on record. The ratio of grain:LWG has been above the overall mean over the past two decades but was below the overall mean from the 1950s to the 70s (Figure 5). The coefficient of variation (CV; 20 year running mean) varied by a factor of 1.8 over the period for wheat and 3 for grass yields but a factor of 4 for LWG. The CV for wheat declined sharply following 1976 and has remained the lowest on record since then. For grass production and LWG, there was a marked decline in CV from the mid-1950s and has remained low except for LWG which has increased to the mean over the last decade. The periods around the 1940s had high variability for all production systems. El Niño and La Niña events had a marked effect on wheat and grass production but smaller effects on LWG (Table 1). EOF3 values show peaks around 1900, the 1930-1940 period and 1980 onwards. Table 1. Effect of average SOI values for June-November on annual wheat yield (kg/ha), grass growth (kg/ha) and LWG (kg/ha) for 1890-1998. Wheat yield Grass growth LWG SOI<-5 1130 2313 25.8 -5<SOI<5 1680 3172 29.4 SOI>5 2100 3573 29.5 Increases in CO2 levels over the past 108 years were simulated to increase wheat production by about 8% and grass production by about 5% with most of this change occurring since the 1960s (data not shown). The CO2 and climate change scenarios gave a range of wheat yields from +12 to +29%, grass yields of +2 to +17% and LWG of +5 to +57% above the means of those experienced since the 1890s using recorded climate and CO2 concentrations. These values straddle the 20-year mean for the most recent decades for wheat and grass production but are higher for LWG (Figure 2,3,4). Doubling of CO2 concentration without climate changes increased the ratios of wheat:grass production and wheat:LWG by 27 and 17% respectively. Changes in CO2 in conjunction with climate changes resulted in little change in this ratio against the Baseline for grass production (except for the Wet scenario with +9%) but around 20% decreases for LWG. 4. Discussion The recent expansion of wheat cropping in the Emerald region appears to have been a response to a changing environment (both climate and CO2 changes). The period of major expansion from 1980 to the early 1990s had by far the greatest mean wheat yields and the 56 lowest variability of yield in the past 108-years. Furthermore, increased yields associated with progressive reductions in frost frequency over this period may underestimate this simulated change (Stone et al. 1996). The relativity of wheat production against grass growth was also the highest on record whilst the ratio of wheat production against LWG was also above average suggesting that the timing of change from grazing to cropping was not just a result of historical development paths or technological change but rather a rational response to a change in the relative productivities and risks of the respective industries. Meinke and Hammer (1995) also have suggested that the peanut industry expansion in south-east Queensland in the 1960s and 1970s was driven by similar climate variations and there are other examples in Australia’s history (eg those relating to the Goyder Line in South Australia; Reyenga et al 1999b). Increases in CO2 probably had a small but positive effect (less than 8%) impact on wheat yields. This suggested response of the cropping industry to growing conditions raises the issue of future stability of the industry in this location. If this favourable ‘window’ for cropping is a result of long-term variability, then return to more average conditions is inevitable and cropping will decline. If the ‘window’ is due to climate change, then some continuation of wheat cropping appears likely provided the incidence of El Niño events doesn’t increase. Simulations of future potential yields under climate change and CO2 increase suggest wheat yields that are 12 to 29% above the 108-year average but similar to those experienced in the high-yielding period in the 1980s. In contrast, in most other wheat cropping areas in Australia, increases over recent yields are likely (Howden et al. 1999b) the relative lack of response being due the likelihood of supra-optimal temperature conditions at Emerald. The historical correspondence of periods of low yield with El Niño events is cause for some concern as there has been a change in the frequency of El Niño events over past decades that may be related to climate change (Cai and Whetton 1999) and the incidence of El Niño events may increase with future global warming (eg Timmerman et al. 1999). The likely expansion or contraction of the wheat industry compared with grazing landuses depended on the future scenario used. Under doubled CO2 without climate changes, the relative productivity of wheat compared with grazing may increase by around 20% suggesting expansion may be possible. Whereas if warmer temperatures occur as well, there may be a 20% reduction of the ratio of wheat yields against liveweight gain but little change against grass production. The increase in LWG was due to increases in the growing season of the native grasses with increases in minimum temperatures and rainfall (Hall et al. 1998). However, GRASP doesn’t simulate the effects of increased heat stress on livestock and this may restrict LWG in these regions leading to lower animal productivities than indicated (Howden et al. 1999c). Nevertheless, even small reductions in relative productivities compared with the 108-year mean suggests contraction of the wheat industry may be likely. Decadal and interdecadal climatic variability, such as evident in the IPO record (Power et al. 1999) manifests itself in corresponding variability of production. In the early part of the record (1890 to the 1950’s) there was a correspondence between peaks in the index values and periods when productivity was low. However, over the past four decades, the relationship appears to be breaking down as the IPO values are high but productivity is also high. Further statistical analyses of the relationship between El Niño, the IPO and wheat yields is currently being undertaken (Meinke unpub. data). Inspection of the simulated production data show two distinct points of change: the 1950s when variability in grazing system productivity declined and the mid 1970s when variability in wheat yields declined. The latter change appears to be related to changes in ocean circulations and temperatures from 1976 onwards 57 (eg Zhang et al. 1998) which appear to have resulted in changes in autumn minimum temperatures and other climate variables in Queensland (McKeon et al. 1998). These results have considerable implications for policymakers dealing with drought issues. The low variability and high productivity of the past few decades are unique in the 108-year record for both grazing and cropping systems. This is likely to have biased the expectations of producers. Return to more normal levels of productivity and variability over the last six years has resulted in claims for ‘Exceptional Circumstances’ drought support. This study suggests that this has not been an unusually poor period if a decadal view is taken. If a 5-year view is adopted, then on average, the last several years have been the poorest on record for the grazing industry. The worst production periods simulated in the record differ depending on the production element being addressed (ie wheat yield, grass production or LWG ) and with the duration being assessed (ie 5, 10 and 20-year windows) as noted by Stafford Smith and McKeon (1999). There exists a challenge to ensure that industry and government policymaking effectively uses such information. 5. References Cai, W. and Whetton, P.H.. (1999) Evidence for a time-varying pattern of greenhouse warming in the Pacific Ocean. Nature, (submitted), Day, K.A., G.M. McKeon and J.O. Carter, (1997)Evaluating the risk of pasture and land degradation in native pastures in Queensland. Final Project Report for RIRDC project DAQ124A. Etheridge, D.M., Steele, L.P., Langenfelds, R.L., Francey, R.J., Barnola, J.-M. and Morgan, V.I. (1996) Natural and anthropogenic changes in atmospheric CO2 over the last 1000 years from air in Antarctic ice and firn. Journal of Geophysical Research 101:4115-4128, Hall, W.B., McKeon, G.M., Carter, J.O., Day, K.A., Howden, S.M., Scanlan, J.C., Johnston, P.W. and Burrows, W.H. (1998) Climate change in Queensland’s grazing lands. II. An assessment of the impact on animal production from native pastures. Rangeland Journal, 20:177-205,. Houghton, J.T., Meira Filho, L.G., Callander, B.A., Harris, N., Kattenberg, A. and Maskell, K. (eds), (1996) Climate Change 1995. The Science of Climate Change, IPCC. Howden, S.M., McKeon, G.M., Walker, L., Carter, J.O., Conroy, J.P, Day, K.A., Hall, W.B., Ash, A.J. and Ghannoum, O. (1999a) Global change impacts on native pastures in south-east Queensland, Australia, Environmental Modelling and Software, 14:307-316. Howden, S.M., Reyenga, P.J. andMeinke, H. (1999b) Global Change Impacts on Australian Wheat Cropping. Report to the Australia Greenhouse Office Howden S.M., Hall, W.B. and Bruget, D. (1999c) Heat stress and beef cattle in Australain rangelands: recent trends and climate change. In People and Rangelands Building the Future. Proceedings of the VI International Rangeland Congress, D.Eldridge and D.Freudenberger (eds.) July 19-23, Townsville, pp 43-45. 58 Isdale, P.J., Stewart, B.J., Tickle, K.S. and Lough, J.M. (1998) Palaeohydrological variation on a tropical river catchment: a reconstruction using fluorescent bands in corals of the Great Barrier Reef, Australia, The Holocene, 8(1):1-8. Keeling, C.D. and Whorf, T.P. (1998) Atmospheric CO2 records from sites in the SIO air sampling network. In: Trends: A Compendium of Data on Global Change. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, Oak Ridge, Tenn., U.S.A. Littleboy, M. and McKeon, G.M. (1997) Subroutine GRASP: Grass production model. Documentation of the Marcoola version of subroutine GRASP. Appendix 2 of Evaluating the risk of pasture and land degradation in native pastures in Queensland. Final Project Report for RIRDC project DAQ124A. McKeon, G.M., Day, K.A., Howden, S.M., Mott, J.J., Orr, D.M., Scattini, W.J. and Weston, E.J. (1990) Northern Australian savannas: management for pastoral production, Journal of Biogeography, 17:355-72. McKeon, G.M., Hall, W.B., Crimp, S.J., Howden, S.M., Stone, R.C., Jones, D.A. (1998) Climate change in Queensland’s grazing lands. I. Approaches and climatic trends, Rangeland Journal 20:151-176. Meinke, H. and Hammer, G.L. (1995) Climatic risk to peanute production: a simulation study for Northern Australia, Australian Journal of Experimental Agriculture, 35:777-780. Meinke, H., Hammer, G.L., Van Keulen, H. and Rabbinge, R. (1998) Improving wheat simulation capabilities in Australia from a cropping systems perspective III. The integrated wheat model (I_WHEAT). European Journal Agronomy 8:101-116. Power, S., Casey, T., Folland, C., Colman, A. and Mehta, V. (1999) Decadal modulation of the impact of ENSO on Australia. Climate Dynamics (submitted). Reyenga, P.J., Howden, S.M., Meinke, H. and McKeon, G.M. (1999) Impacts of global change on cropping in SW Queensland. Environmental Modelling and Software, 14: 297-306. Reyenga, P.J., Howden, S.M., Meinke, H. and Hall, W.B. (in press) Global Change Impacts on Wheat Production along an Environmental Gradient in South Australia. In Modsim’99 International Congress on Modelling and Simulation Proc, December 6-9th, Hamilton, New Zealand. Stafford Smith, D.M and McKeon, G.M. (1999) Assessing the historical frequency of drought events on grazing properties in Australian rangelands, Agricultural Systems, 57:271-299. Stone, R., Nicholls, N., and Hammer, G. (1996) Frost in Northeast Australia: trends and influences of phases of the Southern Oscillation. Journal of Climate, 9:1896-1909. Timmermann, A., Oberhuber, J., Bacher, A., Esch, M., Latif, M. and Roeckner, E. (1999) Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature, 398:694-697. 59 Zhang, R.H., Rothstein, L.M. and Busalacchi, A.J. (1998) Origin of upper-ocean warming and El Niño change on decadal scales in the tropical Pacific Ocean, Nature, 391:879-883. 60