Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Epigenetics wikipedia , lookup
Histone acetyltransferase wikipedia , lookup
Point mutation wikipedia , lookup
Genome (book) wikipedia , lookup
Minimal genome wikipedia , lookup
X-inactivation wikipedia , lookup
Microevolution wikipedia , lookup
Protein moonlighting wikipedia , lookup
Genomic imprinting wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Cancer epigenetics wikipedia , lookup
Designer baby wikipedia , lookup
Long non-coding RNA wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
Gene expression programming wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Gene therapy of the human retina wikipedia , lookup
Epigenetics of neurodegenerative diseases wikipedia , lookup
Epigenetics in stem-cell differentiation wikipedia , lookup
Epigenetics in learning and memory wikipedia , lookup
Epigenetics of diabetes Type 2 wikipedia , lookup
Gene expression profiling wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Mir-92 microRNA precursor family wikipedia , lookup
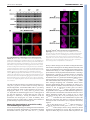
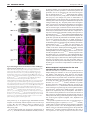
3478 RESEARCH ARTICLE Development 140, 3478-3485 (2013) doi:10.1242/dev.097204 © 2013. Published by The Company of Biologists Ltd The histone H3-K27 demethylase Utx regulates HOX gene expression in Drosophila in a temporally restricted manner Ömer Copur and Jürg Müller* SUMMARY Trimethylation of histone H3 at lysine 27 (H3-K27me3) by Polycomb repressive complex 2 (PRC2) is a key step for transcriptional repression by the Polycomb system. Demethylation of H3-K27me3 by Utx and/or its paralogs has consequently been proposed to be important for counteracting Polycomb repression. To study the phenotype of Drosophila mutants that lack H3-K27me3 demethylase activity, we created UtxΔ, a deletion allele of the single Drosophila Utx gene. UtxΔ homozygotes that contain maternally deposited wild-type Utx protein develop into adults with normal epidermal morphology but die shortly after hatching. By contrast, UtxΔ homozygotes that are derived from Utx mutant germ cells and therefore lack both maternal and zygotic Utx protein, die as larvae and show partial loss of expression of HOX genes in tissues in which these genes are normally active. This phenotype classifies Utx as a trithorax group regulator. We propose that Utx is needed in the early embryo to prevent inappropriate instalment of long-term Polycomb repression at HOX genes in cells in which these genes must be kept active. In contrast to PRC2, which is essential for, and continuously required during, germ cell, embryonic and larval development, Utx therefore appears to have a more limited and specific function during development. This argues against a continuous interplay between H3-K27me3 methylation and demethylation in the control of gene transcription in Drosophila. Furthermore, our analyses do not support the recent proposal that Utx would regulate cell proliferation in Drosophila as Utx mutant cells generated in wild-type animals proliferate like wild-type cells. INTRODUCTION Enzymes that add or remove post-translational modifications at specific amino acid residues of histone proteins have emerged as an important class of transcriptional regulators. Among these regulators are the Polycomb group (PcG) proteins, which form several distinct multiprotein complexes with specific histonemodifying activities (reviewed by Schwartz and Pirrotta, 2008; Müller and Verrijzer, 2009; Beisel and Paro, 2011; Lanzuolo and Orlando, 2012; O’Meara and Simon, 2012; Scheuermann et al., 2012). PcG proteins were first identified through genetic studies in Drosophila, where they are required for the repression of HOX and other developmental regulator genes. In particular, Polycomb complexes are essential for keeping HOX genes inactive in the progeny of cells in which these genes were initially repressed by transcriptional regulators that were transiently present in the early Drosophila embryo. Repression by PcG complexes is therefore thought to comprise an epigenetic memory mechanism that permits the heritable propagation of this repression throughout development (reviewed by Ringrose and Paro, 2004). The discovery that Polycomb repressive complex 2 (PRC2) is a histone methyltransferase that specifically methylates lysine 27 of histone H3 (H3-K27) and that trimethylation of H3-K27 (H3-K27me3) is present across extended stretches of chromatin at PcG-repressed genes, has made this modification an attractive candidate for a chromatin mark that might be inherited through replication and MPI of Biochemistry, Chromatin and Chromosome Biology, Am Klopferspitz 18, 82152 Martinsried, Germany. *Author for correspondence ([email protected]) This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed. Accepted 11 June 2013 mitosis (Cao et al., 2002; Czermin et al., 2002; Kuzmichev et al., 2002; Müller et al., 2002; Bernstein et al., 2006; Papp and Müller, 2006; Schwartz et al., 2006; Trojer and Reinberg, 2006; Mikkelsen et al., 2007; Schuettengruber et al., 2009; Hansen et al., 2008; Margueron et al., 2009; Filion et al., 2010). Recent studies in Drosophila showed that cells in which wild-type histone H3 has been replaced by the nonmethylatable mutant H3K27R fail to maintain Polycomb repression (Pengelly et al., 2013). This provides strong evidence that methylation of H3-K27 is indeed crucial for the repression of Polycomb target genes in Drosophila. Earlier studies identified H3-K27-specific demethylases. In particular, Utx and the related JmjD3 protein were found to demethylate H3-K27me3 in vitro and in vivo (Agger et al., 2007; De Santa et al., 2007; Hong et al., 2007; Lan et al., 2007; Lee et al., 2007; Swigut and Wysocka, 2007). The discovery of these enzymes suggested that H3-K27me3 in chromatin might be added and removed in a dynamic fashion. This challenged the view of H3K27me3 as a stable epigenetic mark that, once installed on an array of nucleosomes, could only be removed by eviction of the modified octamer or by dilution due to deposition of unmodified octamers during DNA replication. To understand the role of H3-K27me3 demethylation, it is essential to determine the phenotype of animals that lack H3K27me3 demethylase activity and to investigate how the expression of PRC2-regulated genes is affected in such animals. Mouse, worm and flies contain different numbers of H3-K27me3 demethylase paralogs that are all characterised by a catalytic JmjC domain with an adjacent zinc-binding domain that is important for substrate specificity (Sengoku and Yokoyama, 2011; Kim and Song, 2011). The mouse genome encodes three H3-K27me3 demethylase family members: the Utx (Kdm6a – Mouse Genome Informatics) gene on the X chromosome, the Uty gene on the Y chromosome and JmjD3 (Kdm6b – Mouse Genome Informatics). Recent studies reported that Utx homozygous mutant female mice die during DEVELOPMENT KEY WORDS: Polycomb, PcG, Trithorax, trxG, Utx, H3-K27me3 Utx function in Drosophila MATERIALS AND METHODS Fly strains The following Drosophila strains were used in this study: y w;Utx∆/CyO, twi:GAL4, UAS:GFP; y w;Utx∆ FRT40A/CyO; y w hs-flp, tubP:GAL4, UAS:nGFP; tubP:GAL80 FRT40A/CyO; y w hs-flp; ovoD1 FRT40A/CyO, hs:hid; w; Utx1 FRT40A/CyO, twi:GAL4, UAS:GFP; w; Df(2L)BSC143/CyO, twi:GAL4, UAS:GFP; y w hs-flp; hs-nGFP FRT40A; y w hs-flp; hs-nGFP F2A; y w; ash122 F2A/TM6C; y w hs-flp; [Utx∆; hs-nGFP F2A]/SM5^TM6; w; [Utx∆; ash122 F2A]/SM5^TM6; and w; FRT40A. Generation of the UtxΔ allele The ends-out recombination strategy was used to generate Utx∆ by replacing the Utx JmjC coding region (Utx503-952) with a miniwhite marker gene following the strategy described (Gong and Golic, 2003). In brief, for the Utx disruption construct, 3.5 kb of Utx 3⬘ coding and downstream sequences (FlyBase 2L:10269938..10273485) and 3.5 kb of 5⬘ coding and upstream sequences (FlyBase 2L:10275043..10278555) were cloned into pw35 (Gong and Golic, 2003). In this construct, the coding region of Utx503-952 [FlyBase 2L:10273487..10275042 (on the minus strand)] was thus replaced by the miniwhite gene from pw35; the AGC codon for Ser503 was replaced with an in-frame TAA termination codon after the His502 codon of Utx. Analysis of survival into adults To determine the fraction of animals developing into adults, three independent batches of 100 first instar larvae of genotype wild type, Utx∆/Utx∆, Utx∆/Df(2L)BSC143, Utx1/Utx1 and Utx1/Df(2L)BSC143 were collected, transferred into a vial with food, and larvae were reared at 25°C. Flies that eclosed from the pupal case were counted. Western blot analysis To analyse Utx protein in wild-type or mutant larvae, total extracts from imaginal discs and CNS tissues of 50 third instar wild-type, Utx∆/Utx∆ or Utx1/Utx1 larvae were prepared by resuspending the tissues in 100 μl 1×LDS buffer (Life Technologies), sonication and then heating at 75°C for 5 minutes. The tissue suspension was centrifuged and 20 μl supernatant of each preparation was run on an 8% SDS polyacrylamide gel. The membrane was probed with anti-Utx420-633 antibody (Tie et al., 2012) or with anti-Utx1153 antibody (Herz et al., 2010). For the analysis of histone modifications, total extracts from imaginal discs and CNS tissues were prepared the same way, using 50 third instar larvae of genotype wild type, Utx∆/Df(2L)BSC143 or Utx1/Df(2L)BSC143. Serial dilutions using 6, 3 and 1.5 μl of each extract were loaded on a 15% SDS polyacrylamide gel. Membranes were probed with anti-H3-K27me3 (1:5000) (Peters et al., 2003), anti-H3-K27ac (1:5000; Active Motif), anti-H3-K4me1 (1:1000; Upstate) or anti-H4 (1:5000; Abcam) antibodies. Immunostaining of embryos and imaginal discs and preparation of adult cuticles Preparation of embryonic cuticles, staining of embryos and larval imaginal discs and clonal analysis were performed following standard protocols. The following antibodies were used: anti-H3-K27me3 (1:300) (Peters et al., 2003), anti-H3-K27ac (1:300; Active Motif), anti-Ubx (1:30) (White and Wilcox, 1984) and anti-Abd-B (1:100) (Celniker et al., 1990). DNA was stained with Hoechst. RESULTS Zygotic Utx function is dispensable for morphogenesis but essential for adult viability To investigate the function of Utx in Drosophila development, we used a homologous recombination strategy (Gong and Golic, 2003) to generate Utx∆, a deletion allele that lacks most of the JmjC catalytic domain and additional portions of the Utx coding region DEVELOPMENT embryonic development with defects in heart and neural tube morphogenesis and that males lacking both Utx and Uty show the same phenotype but that males only lacking Utx develop into normal and fertile adults (Lee et al., 2012; Shpargel et al., 2012; Welstead et al., 2012; Wang et al., 2012; Thieme et al., 2013). Uty thus evidently compensates for the lack of Utx in male Utx mutants (Shpargel et al., 2012). However, there is currently no evidence that Uty has H3-K27me3 demethylase activity, even though the catalytic JmjC domain and the adjacent zinc finger are highly conserved (Hong et al., 2007; Shpargel et al., 2012). Moreover, in a differentiation assay in vitro, knock-in male embryonic stem cells (ESCs) that express full-length but catalytically inactive Utx instead of the wild-type protein show normal induction of mesodermal marker genes, whereas knockout cells lacking Utx protein fail to induce expression of these genes (Wang et al., 2012). Together, these studies led to the suggestion that the Utx/Uty proteins, but not their H3-K27me3 demethylase activity, are crucial for normal embryonic development (Shpargel et al., 2012; Wang et al., 2012). Jmjd3 mutant mice have been reported to be perinatal lethal (Satoh et al., 2010). At present, it is unclear whether the H3-K27me3 demethylase activities of JmjD3 and Utx compensate for each other (Lee et al., 2012; Shpargel et al., 2012; Welstead et al., 2012; Wang et al., 2012). Studies in tissue culture cell models suggest that Utx demethylates H3-K27me3 in target gene chromatin through a mechanism that requires elongating polymerase (Seenundun et al., 2010), whereas JmjD3 has been implicated to be required for the release of poised RNA polymerase II into productive transcriptional elongation (Chen et al., 2012). C. elegans contains utx-1 and three JmjD3 genes (Vandamme et al., 2012). utx-1 mutant worms that are derived from heterozygous mothers and therefore contain maternally deposited wild-type UTX1 protein are viable and show no morphological abnormalities. However, the mutant progeny from utx-1 homozygotes (i.e. lacking both zygotic and maternal UTX-1 protein) arrest development at the late embryonic stage (Vandamme et al., 2012). Intriguingly, this phenotype can be rescued not only by wild-type but also by a catalytically inactive UTX-1 protein, suggesting that UTX-1 protein, but not its H3-K27me3 demethylase activity, is essential for C. elegans development (Vandamme et al., 2012). Worms lacking all three JmjD3 genes are viable and fertile and there is no evidence for a major functional redundancy between UTX-1 and the three JmjD3 proteins (Vandamme et al., 2012). Drosophila contains a single Utx gene, also called dUtx (Smith et al., 2008). Recent studies analysed the phenotype of Drosophila Utx mutants containing a premature termination codon in the C-terminal portion of the JmjC domain (Herz et al., 2010). A large proportion of the animals homozygous for this allele died during the pupal stages, but some were found to develop into adults that showed a spectrum of phenotypes, including rough eyes and mortality shortly after eclosion (Herz et al., 2010). Based on studies with this allele in mosaic animals, it was proposed that Utx acts as a tumour suppressor in Drosophila (Herz et al., 2010). In this study, we generated a Drosophila Utx knockout allele carrying a deletion of the catalytic JmjC domain. Our analyses of Utx mutants provide no evidence for a tumour suppressor function of Utx and show that the zygotic Utx product is dispensable for the normal development of epidermal structures but is essential for adult viability. However, we found that maternally deposited Utx protein in the early embryo is crucial for the normal expression of multiple HOX genes. This work establishes Utx as a trithorax group (trxG) regulator that is specifically required early in embryogenesis to set up long-term stable HOX gene expression patterns. RESEARCH ARTICLE 3479 Fig. 1. Analysis of Utx protein in Utx mutants and requirement for Utx in Drosophila development. (A) Domain architecture of the Drosophila Utx protein showing the tetratricopeptide repeats (TPR) and the catalytic JmjC domain. The lesions of the previously described Utx1 allele (Herz et al., 2010) and of the UtxΔ deletion allele are indicated; in UtxΔ, a Ser503>stop mutation was introduced and a large portion of the JmjC coding region was deleted. (B) Total extracts from imaginal disc and CNS tissues from wild-type (wt, lane 1), homozygous Utx1 (lane 2) and homozygous UtxΔ (lane 3) larvae probed with an antibody against Utx420-633. Note the lack of the 145 and 80 kDa bands (arrowheads) in the Utx1 and UtxΔ mutant extracts. Each lane contained the extract from ten third instar larvae and non-specific cross-reacting bands detected by the antibody provide an additional control for loading of comparable amounts of extract. (C) Survival of wild type and Utx mutants of the indicated genotypes to adults; Df Utx indicates Df(2L)BSC143 and mat+ zyg– indicates that these animals were obtained from Utx1 and UtxΔ heterozygous mothers. mat– zyg– animals were obtained from mothers with UtxΔ germline clones; they arrest development before the pupal stage (no adults were obtained, asterisk) (see main text). For each genotype, three independent batches with 100 first instar larvae each were collected, transferred into vials and the number of eclosed adults was determined. Error bars indicate s.d. (Fig. 1A; supplementary material Fig. S1). Utx∆ is thus a molecularly defined H3-K27 demethylase deficiency allele. We generated extracts from wild-type larvae and from larvae that were homozygous for Utx∆ or Utx1, a previously described point mutation (Fig. 1A) (Herz et al., 2010), and probed them with antibodies that had been raised against a central portion [anti-Utx420-633 (Tie et al., 2012)] or the N-terminus [anti-dUtx1-153 (Herz et al., 2010), referred to here as anti-Utx1-153] of the Utx protein. In wild-type extracts, the anti-Utx420-633 antibody detected two bands with apparent molecular weights of 145 and 80 kDa that were both absent in Utx∆ or Utx1 mutant extracts (Fig. 1B). This suggests that both the 145 and the 80 kDa proteins are Utx products. Although the 80 kDa band appears more intense than the 145 kDa band and could thus represent an alternate, shorter Utx product, it is also possible that the 80 kDa band represents a degradation product of the 145 kDa protein. The anti-Utx1-153 antibody detected the 145 kDa but not the 80 kDa band in wild-type extracts, suggesting that the 80 kDa Utx product lacks the N-terminus (data not shown). The anti-Utx1-153 antibody also no longer detected the 145 kDa band in Utx∆ extracts, but instead detected an extra band of ~55 kDa that is absent in wildtype extracts and is likely to correspond to the Utx1-502 protein encoded by Utx∆ (data not shown). In summary, the molecularly Development 140 (16) defined deletion of the JmjC domain in Utx∆ and these western blot analyses demonstrate that Utx∆ is a demethylase null mutation. In a first set of experiments, we examined the phenotype of Utx∆/Df(2L)BSC143 transheterozygotes that were derived from females with a Utx∆ heterozygous germ line. Although these mutants do not express wild-type Utx protein from their genome, they still contain wild-type Utx product during the earliest stages of development, deposited in the egg by the mother. We refer to these as Utx∆ mat+ zyg– mutants to distinguish them from Utx∆ mat– zyg– mutants that are derived from females with Utx∆ germ cells and which therefore completely lack wild-type Utx protein (see below). The majority of Utx∆ mat+ zyg– animals develop into adults that are morphologically indistinguishable from wild-type flies, but die within a day after eclosion from the pupal case (Fig. 1C). Previous studies reported that animals homozygous for Utx1 only rarely survive to adults and that surviving individuals show a rough eye phenotype and die shortly after eclosion (Herz et al., 2010). We found that Utx1/Utx1 homozygotes indeed only rarely survive to adults (Fig. 1C) but that Utx1/Df(2L)BSC143 transheterozygotes develop into adults at a frequency comparable to Utx∆/Df(2L)BSC143 transheterozygotes or Utx∆ homozygotes and that these animals show no detectable morphological defects (Fig. 1C; data not shown). As in the case of Utx∆ adults, Utx1/Df(2L)BSC143 transheterozygous adults also die shortly after eclosion. We conclude that zygotic Utx function is not required for the development of morphologically normal epidermal structures but, for unknown reasons, it is essential for adult viability. Global H3-K27me3 and H3-K27ac levels are unaffected in Utx mutants We next investigated how loss of Utx demethylase activity affects the global levels of H3-K27me3. Previous studies reported that bulk H3-K27me3 levels in Utx1 homozygous larvae or in Utx RNAitreated adults are increased compared with wild-type animals (Herz et al., 2010; Tie et al., 2012) and that bulk levels of histone H3 acetylation at lysine 27 (H3-K27ac) are decreased (Tie et al., 2012). We performed western blot analyses on serially diluted extracts from Utx∆/Df(2L)BSC143 or Utx1/Df(2L)BSC143 larvae with antiH3-K27me3 and anti-H3-K27ac antibodies. We were unable to detect a change in the bulk levels of these modifications in Utx mutants compared with wild-type larvae (Fig. 2A). Probing the same material with an antibody against H3-K4me1, we found a very small but reproducible decrease in bulk H3-K4me1 levels in Utx mutants (Fig. 2A), consistent with a previous report (Herz et al., 2010). We also analysed H3-K27me3, H3-K27ac and H3-K4me1 levels by immunofluorescence labelling of imaginal wing discs with clones of Utx∆ or Utx1 cells. We found that H3-K27me3 and H3K27ac, but also H3-K4me1, immunofluorescence signals in Utx∆ or Utx1 cells were comparable to those in neighbouring wild-type cells in the same tissue (Fig. 2B; data not shown). We conclude that lack of Utx has no major effect on global H3-K27me3 and H3K27ac levels but results in a very small reduction in global H3K4me1 levels. Utx mutant cells proliferate like wild-type cells Previous studies reported that Utx1 cell clones in imaginal discs show a growth advantage relative to neighbouring wild-type cells and proposed that Utx acts as a tumour suppressor (Herz et al., 2010). We used the exact same experimental conditions as Herz and co-workers (Herz et al., 2010) to analyse the size and growth of Utx∆ or Utx1 cell clones in the eye-antenna imaginal disc but found that both Utx∆ and Utx1 clones grow like control clones of wild-type DEVELOPMENT 3480 RESEARCH ARTICLE Fig. 2. Bulk H3-K27me3 and H3-K27ac levels are unchanged in Utx mutants. (A) Western blots of serial dilutions (4:2:1) of total extracts from imaginal disc and CNS tissues of wild-type (wt, lanes 1-3), Utx1/Df(2L)BSC143 (lanes 4-6) and UtxΔ/Df(2L)BSC143 (lanes 7-9) larvae, probed with antibodies against H3-K27me3, H3-K27ac and H3-K4me1. In each case, the membrane was simultaneously probed with an antibody against histone H4 to control for extract loading and western blot processing but was exposed to film for a shorter period of time in order not to saturate the H4 signal. Note that H3K27me3 and H3-K27ac levels are unchanged in Utx mutant extracts but that H3-K4me1 levels are very slightly (less than one-third) decreased in the mutants. (B) Wing imaginal discs with MARCM clones of UtxΔ homozygous cells that are marked by the presence of GFP, stained with H3-K27me3 or H3K27ac antibodies and Hoechst (for DNA). Note that the H3-K27me3 and H3K27ac immunofluorescence signals in UtxΔ/UtxΔ clone cells are comparable to those in the neighbouring UtxΔ/+ cells. Discs were analysed 96 hours after clone induction. cells that we induced as reference in a parallel experiment (Fig. 3). Using the ey-flp driver to induce clones across the entire eyeantenna imaginal disc, we found that adults with Utx1 eyes show a rough eye phenotype similar to that reported for surviving Utx1 homozygotes (Herz et al., 2010) (data not shown). Since Utx1/Df(2L)BSC143 transheterozygotes and Utx∆ homozygotes do not show this phenotype, it seems likely that this rough eye phenotype is caused by a second site mutation on the 2L chromosome arm carrying the Utx1 allele. In summary, these analyses provide no evidence that Utx mutant cells would have a growth advantage compared with wild-type cells. Maternally deposited Utx is essential for stably activated HOX gene expression We next analysed the phenotype of mutants that lack both maternal and zygotic Utx product. We generated Utx∆/Df(2L)BSC143 animals from females with Utx∆ germ cells. These Utx∆ mat– zyg– animals RESEARCH ARTICLE 3481 Fig. 3. Utx1 and UtxΔ clones do not show an overproliferation phenotype. Eye-antenna imaginal discs with MARCM clones of Utx1 or UtxΔ homozygous cells marked by the presence of GFP and stained with Hoechst. As reference, clones of a wild-type 2L chromosome arm (wt) were generated the same way and are also positively marked by GFP (top). In all cases, discs were analysed 96 hours after clone induction. Note that the clone sizes are comparable in all three genotypes and that the clone cells populate a comparable area of disc tissue. develop to the larval stages; 85% die before reaching the third larval instar and the remaining larvae die during the third larval instar, with only a very small fraction (0.6%) developing to form white prepupae, which then also die (Fig. 1C). Thus, complete absence of Utx does not permit animals to develop beyond the third larval instar or to undergo metamorphosis, suggesting an essential function of maternally supplied Utx products. Nevertheless, if Utx∆ oocytes (i.e. derived from females with a Utx∆ germ line) are fertilised by wild-type sperm, zygotic expression of Utx protein rescues these Utx∆ mat– zyg+ animals into viable and fertile adults. The majority of such Utx∆ mat– zyg+ adults show wild-type morphology but a substantial fraction shows homeotic transformations that are reminiscent of trithorax (trx) mutants (Ingham and Whittle, 1980) and are likely to be caused by a reduction or loss of expression of HOX genes (see below). Specifically, 30% of Utx∆ mat– zyg+ males (n=382) show a patchy loss of pigmentation in the tergite of the fifth abdominal segment (A5), which represents a transformation of the affected tissue into a more anterior abdominal segment and is caused by loss of Abd-B expression in A5 (Fig. 4A). At very low frequency (2.8%, n=758), Utx∆ mat– zyg+ individuals also show transformation of parts of the haltere into wing tissue (not shown), a homeotic transformation caused by the loss of Ultrabithorax (Ubx) expression in this body segment. The trx-like phenotypes in Utx∆ mat– zyg+ adults prompted us to analyse the expression of HOX genes in Utx∆ mat– zyg– animals. Utx∆ mat– zyg– mutant embryos show a strong reduction of Abd-B expression in parasegment (ps) 10 (Fig. 4B), from which the A5 segment of adults (Fig. 4A) is derived. Abd-B expression in the DEVELOPMENT Utx function in Drosophila 3482 RESEARCH ARTICLE Development 140 (16) Fig. 4. Trithorax-like homeotic transformations and loss of HOX gene expression in UtxΔ mutants. (A) Portions of the dorsal abdomen of adult males of the indicated genotype, showing the tergites of abdominal segments A4, A5 and A6. Note the patchy loss of pigmentation in the anterior part of A5 of the UtxΔ mat– zyg+ male (arrowheads), suggesting a transformation of this tissue to A4 segment identity. (B) Ventral views of stage 16 embryos stained with antibodies against the HOX proteins Ubx and Abd-B. The UtxΔ mat– zyg– genotype refers to UtxΔ/Df(2L)BSC143 embryos derived from females with a UtxΔ germ line. Ubx expression is indistinguishable from wild type (wt). Abd-B expression is strongly reduced in parasegment (ps) 10 (arrowhead) but appears largely undiminished in more posterior parasegments; a vertical bar marks the anterior margin of the normal Abd-B expression domain in ps 10 in the CNS. (C) Haltere imaginal discs from third instar larvae stained with Ubx antibody and Hoechst. Mutant genotypes are UtxΔ/Df(2L)BSC143 derived from a UtxΔ heterozygous mother (UtxΔ mat+ zyg–, middle) and from a female with a UtxΔ germ line (UtxΔ mat– zyg–, bottom). Note the complete loss of Ubx expression in a large fraction of cells in the UtxΔ mat– zyg– disc (asterisk) and that Ubx expression in the UtxΔ mat+ zyg– disc is indistinguishable from the wild type. The loss of Ubx expression is not compartment specific and always occurs in a clone-like pattern with large patches of Ubx-negative cells. more posterior ps 11-14 appears largely undiminished with the exception of occasionally reduced expression in a few cells in ps 11 (Fig. 4B). We were unable to detect a reduction of Ubx expression in Utx∆ mat– zyg– embryos compared with wild-type embryos (Fig. 4B). We next analysed HOX gene expression in imaginal discs of Utx∆ mat– zyg– larvae that survived to the third larval instar (see above). Distinct temporal requirements for the trxG regulators Utx, Trx and Ash1 The observation that Utx is required for the long-term expression of multiple HOX genes classifies Utx as a trxG regulator (Kennison and Tamkun, 1988; Kennison, 1995). However, the temporal requirement for Utx during the Drosophila life cycle is very distinct from that of other trxG proteins such as Trx and Ash1. Specifically, maternally deposited Utx product is sufficient to permit Drosophila to develop into adults with morphologically normal epidermal structures (i.e. in Utx∆ mat+ zyg– animals). Maternal Utx product becomes extensively diluted during embryonic and larval cell divisions and Utx∆ mat+ zyg– third instar larvae indeed lack detectable levels of wild-type Utx protein (Fig. 1B). For the regulation of HOX gene expression – though not for viability of adults – Utx therefore appears to be required only early in development. This stands in clear contrast to the requirement for Trx and Ash1; the function of these proteins is needed throughout development and removal of either protein from wild-type cells during the larval stages results in loss of HOX gene expression (Ingham, 1981; Klymenko and Müller, 2004). One possible explanation for the lack of a trxG phenotype in animals lacking zygotic Utx protein (i.e. in Utx∆ mat+ zyg– mutants) could be that Utx function during larval development is redundant in the presence of Ash1 or Trx. We investigated this possibility as follows. Clones of cells that are homozygous for the protein-null mutation ash122 in haltere or third leg imaginal discs show loss of Ubx expression in the majority of cells but a fraction of cells DEVELOPMENT In wild-type animals, Ubx is expressed in all cells of the haltere and third leg discs, Abd-B is expressed in a characteristic pattern in the genital disc, and Scr is expressed in all cells of the first leg disc (Fig. 4C; data not shown). Utx∆ mat– zyg– mutant larvae show complete loss of Ubx expression in a patchy pattern in haltere and third leg discs (Fig. 4C). Even though Ubx protein is undetectable in a substantial fraction of cells in these discs, its expression level in the remaining cells in these tissues appears undiminished compared with wild-type animals (Fig. 4C). The patchy nature of the Ubx-negative tissue, as opposed to a salt-and-pepper distribution of Ubx-negative and -positive cells across the tissue, suggests that the Ubx-negative cells are of clonal origin, a notion that we have not been able to test. This Ubx loss of expression phenotype was observed in all third instar larvae analysed, but the fraction of cells that show complete loss of expression varied between individuals. Abd-B protein expression in the genital disc of Utx∆ mat– zyg– larvae appeared indistinguishable from that of wild-type animals (not shown). It should be noted that the genital disc represents the primordium for abdominal segment A8, A9 and A10 structures and that loss of Abd-B expression in Utx∆ mat– zyg– embryos was restricted to ps 10, which is the primordium of abdominal segment A5 in adults (Fig. 4B). Similarly, the homeotic transformations in Utx∆ mat– zyg+ adults were also limited to A5 (Fig. 4A). In Utx mutants, Abd-B expression is thus only affected in a subset of cells of the Abd-B expression domain. Finally, we found Scr protein expression in first leg discs of Utx∆ mat– zyg– larvae to be indistinguishable from that of wild type (data not shown). We also note that, in imaginal discs of Utx∆ mat+ zyg– animals, expression of all three HOX genes (Ubx, Abd-B and Scr) was indistinguishable from wild type, consistent with the lack of homeotic phenotypes in adults of this genotype (Fig. 4C; data not shown). Taken together, these data show that Utx function in the very early Drosophila embryo is required for the stable expression of the HOX genes Ubx and Abd-B, but that zygotic Utx function is dispensable for normal HOX gene expression. Utx function in Drosophila RESEARCH ARTICLE 3483 dispensable for normal morphogenesis of epidermal structures, Utx∆ mat+ zyg– adults nevertheless die shortly after eclosion from the pupal case. It is possible that this lethality is caused by defective morphogenesis of internal tissues or organs. Alternatively, Utx might be needed for some other vital function in adult Drosophila. Below, we discuss in turn the conclusions that can be drawn from these results and consider possible mechanisms as to how Utx might regulate HOX gene expression. nevertheless maintains high or moderate levels of Ubx expression (Fig. 5) (see Klymenko and Müller, 2004). This all-or-none loss of expression phenotype is also observed in ash122 homozygotes, and it correlates with the accumulation of H3-K27me3 at the Ubx promoter and coding regions in haltere and third leg imaginal disc cells (Papp and Müller, 2006). If zygotically expressed Utx protein synergised with Ash1 in imaginal disc cells to maintain HOX gene expression, one might expect a more extensive loss of Ubx expression in clones of Utx∆; ash122 double-mutant cells. We therefore induced clones of ash122 homozygous cells in Utx∆ mat+ zyg– animals (Fig. 5). However, we found that the fraction of cells that maintain high levels of Ubx expression in these ash122 Utx∆ double-mutant clones was comparable to that in ash122 singlemutant clones (Fig. 5). Thus, even in a genetic background in which another trxG regulator is removed, there is no evidence for a role of zygotic Utx protein in HOX gene regulation. DISCUSSION In this study, we used a Utx allele with a deletion of the catalytic domain to investigate the role of Utx and its H3-K27 demethylase activity in Drosophila development. Analyses of animals that lack maternal and zygotic Utx product uncovered that Utx protein is crucial for the long-term stable expression of multiple HOX genes. By contrast, animals that lack only zygotic expression of Utx develop into adults with morphologically normal epidermal structures, suggesting that zygotic Utx function is not needed for HOX gene regulation. Moreover, in animals lacking maternally deposited Utx protein, zygotic Utx product alone is unable to fully rescue normal HOX gene expression and these animals show homeotic transformations (i.e. in Utx∆ mat– zyg+ animals). Together, these data suggest that Utx function is crucial for HOX gene regulation right at the onset of zygotic transcription in the early embryo. Even though zygotic Utx expression is apparently Possible molecular mechanisms of HOX gene regulation by Utx We envisage two possible mechanisms of how Utx could function to activate the expression of HOX genes, one of which depends on the H3-K27me3 demethylase activity of Utx and another that acts independently of this enzymatic activity. A first scenario would be that H3-K27me3 demethylation by Utx in the early embryo removes ‘ectopic’ H3-K27 trimethylation by PRC2 at HOX genes in cells in which these genes need to be expressed. According to this view, the loss of HOX gene expression in Utx∆ mat– zyg– animals would be due to ectopic placement of H3K27me3 and, as a consequence, PcG silencing of HOX genes within their normal expression domains. H3-K27me3 demethylation by Utx would thus act to antagonise the establishment of PcG repression. Recent studies reported the genome-wide binding profile of Utx in Drosophila tissue culture cells and found that Utx binding colocalises tightly with the binding of PcG protein complexes at Polycomb response elements (PREs) of HOX genes (Tie et al., 2012). This supports the idea that the function of Utx in HOX gene DEVELOPMENT Fig. 5. Lack of Utx function does not exacerbate loss of HOX gene expression in ash1 mutants. Haltere imaginal discs from third instar larvae with clones of ash122 homozygous cells were stained with Ubx antibody and Hoechst. ash122 mutant cells are marked by the absence of GFP and were induced in a wild-type genetic background (ash122, top) or in the background of UtxΔ mat+ zyg– homozygous larvae (UtxΔ, ash122, bottom). Discs were analysed 96 hours after clone induction. In each sample, the boundary of one clone is outlined. Note that in both genotypes a fraction of cells in the clone shows complete loss of Ubx expression (open arrowheads), whereas other cells maintain high to moderate levels of Ubx expression (white arrowheads). The requirement for Utx function is more limited than for PRC2 Considering that Utx is the only known H3-K27 demethylase in Drosophila, it is interesting to compare the requirement for Utx with that for PRC2 during Drosophila development. First, germ cells with null mutations in different PRC2 subunits fail to develop (Phillips and Shearn, 1990; Birve et al., 2001), but Utx∆ mutant germ cells develop into oocytes that support embryonic development (this study). PRC2 is thus essential for germ cell development whereas Utx is not. Second, removal of PRC2 subunits in somatic cells at any time point during development (e.g. in imaginal disc cells in larvae) results in misexpression of PRC2 target genes and cell fate changes (Birve et al., 2001; Müller et al., 2002), but animals lacking zygotic Utx expression develop into adults with normal epidermal morphology. PRC2 is thus continuously required during embryonic and larval development. By contrast, Utx is crucial during embryogenesis and in adults but development from the third larval instar onwards, through pupal development and metamorphosis, appears to occur largely normally in the absence of detectable Utx protein (Fig. 1B). Taken together, this argues against a view that transcriptional regulation of PRC2 target genes during development involves a constant interplay between H3-K27 methylation and demethylation. In this context, it is interesting to note that, in second instar larvae that lack zygotic PRC2 function but had survived up to this stage because of maternally supplied PRC2, H3-K27me3 levels are comparable to those in wild-type larvae at the same stage (Nekrasov et al., 2007). This suggests that H3-K27me3 that was generated by maternally supplied PRC2 remains stable when the supply of maternal PRC2 disappears and de novo deposition of this modification ceases. This further supports the notion that H3K27me3 is a stable modification in Drosophila that is apparently not subject to constant turnover. chromatin is intimately linked to that of PRC2. Below, we discuss the possible relationship between this proposed role of Utx in antagonising H3-K27me3 deposition by PRC2 and the role of Ash1 and Trx in the same process. As discussed in the Introduction, previous studies reported that Utx protein, but not its H3-K27me3 demethylase activity, is essential for normal C. elegans development (Vandamme et al., 2012) and for the correct differentiation of murine ESCs in vitro (Wang et al., 2012). An alternative scenario would thus be that Drosophila Utx promotes HOX gene transcription by a demethylase-independent mechanism. How could this work? As in worms and mammals, Drosophila Utx is a component of an MLL3/4-like complex that includes the H3-K4 methyltransferase Trithorax-related (Trr) (Cho et al., 2007; Issaeva et al., 2007; Lee et al., 2007; Patel et al., 2007; Mohan et al., 2011; Vandamme et al., 2012). Recent studies in Drosophila proposed that Trr is required for genome-wide H3-K4 monomethylation at enhancers and that H3K27me3 demethylation of enhancers by Trr-associated Utx would be a general mechanism to convert inactive to active enhancers (Herz et al., 2012). Although we have been unable to detect changes in bulk H3-K27me3 and H3-K27ac levels in Utx mutants, we found a slight decrease in bulk H3-K4me1 levels (Fig. 2A), consistent with earlier studies (Herz et al., 2010). This decrease in H3-K4me1 is not simply due to a reduction in Trr protein levels because these were found to be undiminished after RNAi-mediated knockdown of Utx (Herz et al., 2012). One possibility would thus be that Utx is needed for H3-K4 monomethylation by Trr and thereby contributes to the activation of HOX gene expression in the early embryo. trr zygotic mutants die as embryos without any obvious trxG mutant phenotypes (Sedkov et al., 1999), but the phenotype of embryos lacking maternal and zygotic Trr protein is not known and so it is currently unclear whether Trr plays a role in HOX gene regulation in the early embryo. In the context of this scenario, we would emphasize that Utx∆ mat– zyg– mutants develop to the larval stages and, with some notable exceptions (i.e. the regulation of Abd-B expression in ps 10 in the embryo), the enhancer-mediated activation of genes required for embryonic patterning must therefore by and large occur normally in the absence of Utx. Similarly, Utx∆ mat+ zyg– larvae develop into morphologically normal adults and the reduction of H3-K4me1 levels in these animals does not therefore seem to compromise enhancer activity in any drastic manner. Relationship between Utx, Ash1 and Trx in maintaining HOX gene expression Utx, Ash1 and Trx are all required for the normal stable expression of multiple HOX genes. Previous studies provided several independent lines of genetic and biochemical evidence that Trx and Ash1 keep HOX genes active by antagonising PRC2 activity. First, HOX gene expression is lost in ash1 mutants but is restored in ash1 mutants that also lack E(z), the catalytic subunit of PRC2, suggesting that Ash1 is not needed for HOX gene transcription but rather to prevent PRC2 from shutting down HOX gene expression (Klymenko and Müller, 2004). Second, chromatin immunoprecipitation studies of the HOX gene Ubx in imaginal discs have shown that although PRC2 is also bound at Ubx in cells in which it is transcribed, Ash1 inhibits deposition of H3-K27me3 at the promoter and coding region in these cells (Papp and Müller, 2006). Third, on recombinant nucleosomes, the histone modifications H3-K36me2/3 and H3-K4me3 that are generated by Ash1 and Trx, respectively, strongly inhibit PRC2 from methylating H3-K27 on the same histone tail (Schmitges et al., 2011; Yuan et al., 2011). Development 140 (16) Here, we found that removal of Utx in imaginal disc cells has no effect on HOX gene expression. Moreover, in ash1 mutant haltere and third leg imaginal disc cells where PRC2 deposits faulty H3K27 trimethylation in the Ubx promoter and coding region (Papp and Müller, 2006), and where Ubx expression is lost in many although not all cells, removal of Utx does not exacerbate this loss of Ubx expression (Fig. 5). This suggests that, at least in larvae, H3K27me3 demethylation is not crucial for antagonising the formation of repressive H3-K27me3 chromatin on active HOX genes. During larval development, H3-K4 and H3-K36 trimethylation by Trx, Ash1 and possibly other H3-K4 and H3-K36 histone methyltransferases therefore appears to be the predominant mechanism that prevents HOX gene-bound PRC2 from depositing H3-K27me3. This might, however, be different in the early embryo. It is possible that during the establishment of HOX gene expression domains at the blastoderm stage, H3-K4me3 and H3-K36me2/3 marks are not yet fully installed on HOX gene chromatin and that it is during this critical time window that demethylation by Utx is needed to remove faulty H3-K27me3 deposition from HOX genes in cells in which they need to be expressed. Acknowledgements We thank A. Bergmann, P. Harte and Ali Shilatifard for the generous gift of fly strains and antibodies; Andres Gaytan de Ayala and Tony Chen for help with generating the UtxΔ disruption construct; and Sandra Müller for generation of transgenic flies. Funding This work was supported by funds from the MPI of Biochemistry. Deposited in PMC for immediate release. Competing interests statement The authors declare no competing financial interests. Author contributions O.C. and J.M. designed the experiments; O.C. carried out the experiments; J.M. supervised the work and wrote the paper. Supplementary material Supplementary material available online at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.097204/-/DC1 References Agger, K., Cloos, P. A., Christensen, J., Pasini, D., Rose, S., Rappsilber, J., Issaeva, I., Canaani, E., Salcini, A. E. and Helin, K. (2007). UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 449, 731-734. Beisel, C. and Paro, R. (2011). Silencing chromatin: comparing modes and mechanisms. Nat. Rev. Genet. 12, 123-135. Bernstein, B. E., Mikkelsen, T. S., Xie, X., Kamal, M., Huebert, D. J., Cuff, J., Fry, B., Meissner, A., Wernig, M., Plath, K. et al. (2006). A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 125, 315-326. Birve, A., Sengupta, A. K., Beuchle, D., Larsson, J., Kennison, J. A., Rasmuson-Lestander A, and Müller, J. (2001). Su(z)12, a novel Drosophila Polycomb group gene that is conserved in vertebrates and plants. Development 128, 3371-3379. Cao, R., Wang, L., Wang, H., Xia, L., Erdjument-Bromage, H., Tempst, P., Jones, R. S. and Zhang, Y. (2002). Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science 298, 1039-1043. Celniker, S. E., Sharma, S., Keelan, D. J. and Lewis, E. B. (1990). The molecular genetics of the bithorax complex of Drosophila: cis-regulation in the Abdominal-B domain. EMBO J. 9, 4277-4286. Chen, S., Ma, J., Wu, F., Xiong, L. J., Ma, H., Xu, W., Lv, R., Li, X., Villen, J., Gygi, S. P. et al. (2012). The histone H3 Lys 27 demethylase JMJD3 regulates gene expression by impacting transcriptional elongation. Genes Dev. 26, 13641375. Cho, Y. W., Hong, T., Hong, S., Guo, H., Yu, H., Kim, D., Guszczynski, T., Dressler, G. R., Copeland, T. D., Kalkum, M. et al. (2007). PTIP associates with MLL3- and MLL4-containing histone H3 lysine 4 methyltransferase complex. J. Biol. Chem. 282, 20395-20406. Czermin, B., Melfi, R., McCabe, D., Seitz, V., Imhof, A. and Pirrotta, V. (2002). Drosophila enhancer of Zeste/ESC complexes have a histone H3 DEVELOPMENT 3484 RESEARCH ARTICLE methyltransferase activity that marks chromosomal Polycomb sites. Cell 111, 185196. De Santa, F., Totaro, M. G., Prosperini, E., Notarbartolo, S., Testa, G. and Natoli, G. (2007). The histone H3 lysine-27 demethylase Jmjd3 links inflammation to inhibition of polycomb-mediated gene silencing. Cell 130, 1083-1094. Filion, G. J., van Bemmel, J. G., Braunschweig, U., Talhout, W., Kind, J., Ward, L. D., Brugman, W., de Castro, I. J., Kerkhoven, R. M., Bussemaker, H. J. et al. (2010). Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. Cell 143, 212-224. Gong, W. J. and Golic, K. G. (2003). Ends-out, or replacement, gene targeting in Drosophila. Proc. Natl. Acad. Sci. USA 100, 2556-2561. Hansen, K. H., Bracken, A. P., Pasini, D., Dietrich, N., Gehani, S. S., Monrad, A., Rappsilber, J., Lerdrup, M. and Helin, K. (2008). A model for transmission of the H3K27me3 epigenetic mark. Nat. Cell Biol. 10, 1291-1300. Herz, H. M., Madden, L. D., Chen, Z., Bolduc, C., Buff, E., Gupta, R., Davuluri, R., Shilatifard, A., Hariharan, I. K. and Bergmann, A. (2010). The H3K27me3 demethylase dUTX is a suppressor of Notch- and Rb-dependent tumors in Drosophila. Mol. Cell. Biol. 30, 2485-2497. Herz, H. M., Mohan, M., Garruss, A. S., Liang, K., Takahashi, Y. H., Mickey, K., Voets, O., Verrijzer, C. P. and Shilatifard, A. (2012). Enhancer-associated H3K4 monomethylation by Trithorax-related, the Drosophila homolog of mammalian Mll3/Mll4. Genes Dev. 26, 2604-2620. Hong, S., Cho, Y. W., Yu, L. R., Yu, H., Veenstra, T. D. and Ge, K. (2007). Identification of JmjC domain-containing UTX and JMJD3 as histone H3 lysine 27 demethylases. Proc. Natl. Acad. Sci. USA 104, 18439-18444. Ingham, P. (1981). Trithorax: a new homoeotic mutation of Drosophila melanogaster. Rouxs Arch. Dev. Biol. 190, 365-369. Ingham, P. and Whittle, R. (1980). Trithorax: a new homeotic mutation of Drosophila melanogaster causing transformations of abdominal and thoracic imaginal segments. Mol. Gen. Genet. 179, 607-614. Issaeva, I., Zonis, Y., Rozovskaia, T., Orlovsky, K., Croce, C. M., Nakamura, T., Mazo, A., Eisenbach, L. and Canaani, E. (2007). Knockdown of ALR (MLL2) reveals ALR target genes and leads to alterations in cell adhesion and growth. Mol. Cell. Biol. 27, 1889-1903. Kennison, J. A. (1995). The Polycomb and trithorax group proteins of Drosophila: trans-regulators of homeotic gene function. Annu. Rev. Genet. 29, 289-303. Kennison, J. A. and Tamkun, J. W. (1988). Dosage-dependent modifiers of polycomb and antennapedia mutations in Drosophila. Proc. Natl. Acad. Sci. USA 85, 8136-8140. Kim, E. and Song, J. J. (2011). Diverse ways to be specific: a novel Zn-binding domain confers substrate specificity to UTX/KDM6A histone H3 Lys 27 demethylase. Genes Dev. 25, 2223-2226. Klymenko, T. and Müller, J. (2004). The histone methyltransferases Trithorax and Ash1 prevent transcriptional silencing by Polycomb group proteins. EMBO Rep. 5, 373-377. Kuzmichev, A., Nishioka, K., Erdjument-Bromage, H., Tempst, P. and Reinberg, D. (2002). Histone methyltransferase activity associated with a human multiprotein complex containing the Enhancer of Zeste protein. Genes Dev. 16, 2893-2905. Lan, F., Bayliss, P. E., Rinn, J. L., Whetstine, J. R., Wang, J. K., Chen, S., Iwase, S., Alpatov, R., Issaeva, I., Canaani, E. et al. (2007). A histone H3 lysine 27 demethylase regulates animal posterior development. Nature 449, 689-694. Lanzuolo, C. and Orlando, V. (2012). Memories from the polycomb group proteins. Annu. Rev. Genet. 46, 561-589. Lee, M. G., Villa, R., Trojer, P., Norman, J., Yan, K. P., Reinberg, D., Di Croce, L. and Shiekhattar, R. (2007). Demethylation of H3K27 regulates polycomb recruitment and H2A ubiquitination. Science 318, 447-450. Lee, S., Lee, J. W. and Lee, S. K. (2012). UTX, a histone H3-lysine 27 demethylase, acts as a critical switch to activate the cardiac developmental program. Dev. Cell 22, 25-37. Margueron, R., Justin, N., Ohno, K., Sharpe, M. L., Son, J., Drury, W. J., III, Voigt, P., Martin, S. R., Taylor, W. R., De Marco, V. et al. (2009). Role of the polycomb protein EED in the propagation of repressive histone marks. Nature 461, 762-767. Mikkelsen, T. S., Ku, M., Jaffe, D. B., Issac, B., Lieberman, E., Giannoukos, G., Alvarez, P., Brockman, W., Kim, T. K., Koche, R. P. et al. (2007). Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448, 553-560. Mohan, M., Herz, H. M., Smith, E. R., Zhang, Y., Jackson, J., Washburn, M. P., Florens, L., Eissenberg, J. C. and Shilatifard, A. (2011). The COMPASS family of H3K4 methylases in Drosophila. Mol. Cell. Biol. 31, 4310-4318. Müller, J. and Verrijzer, P. (2009). Biochemical mechanisms of gene regulation by polycomb group protein complexes. Curr. Opin. Genet. Dev. 19, 150-158. Müller, J., Hart, C. M., Francis, N. J., Vargas, M. L., Sengupta, A., Wild, B., Miller, E. L., O’Connor, M. B., Kingston, R. E. and Simon, J. A. (2002). Histone methyltransferase activity of a Drosophila Polycomb group repressor complex. Cell 111, 197-208. Nekrasov, M., Klymenko, T., Fraterman, S., Papp, B., Oktaba, K., Köcher, T., Cohen, A., Stunnenberg, H. G., Wilm, M. and Müller, J. (2007). Pcl-PRC2 is needed to generate high levels of H3-K27 trimethylation at Polycomb target genes. EMBO J. 26, 4078-4088. RESEARCH ARTICLE 3485 O’Meara, M. M. and Simon, J. A. (2012). Inner workings and regulatory inputs that control Polycomb repressive complex 2. Chromosoma 121, 221-234. Papp, B. and Müller, J. (2006). Histone trimethylation and the maintenance of transcriptional ON and OFF states by trxG and PcG proteins. Genes Dev. 20, 20412054. Patel, S. R., Kim, D., Levitan, I. and Dressler, G. R. (2007). The BRCT-domain containing protein PTIP links PAX2 to a histone H3, lysine 4 methyltransferase complex. Dev. Cell 13, 580-592. Pengelly, A. R., Copur, O., Jäckle, H., Herzig, A. and Müller, J. (2013). A histone mutant reproduces the phenotype caused by loss of histone-modifying factor Polycomb. Science 339, 698-699. Peters, A. H., Kubicek, S., Mechtler, K., O’Sullivan, R. J., Derijck, A. A., PerezBurgos, L., Kohlmaier, A., Opravil, S., Tachibana, M., Shinkai, Y. et al. (2003). Partitioning and plasticity of repressive histone methylation states in mammalian chromatin. Mol. Cell 12, 1577-1589. Phillips, M. D. and Shearn, A. (1990). Mutations in polycombeotic, a Drosophila polycomb-group gene, cause a wide range of maternal and zygotic phenotypes. Genetics 125, 91-101. Ringrose, L. and Paro, R. (2004). Epigenetic regulation of cellular memory by the Polycomb and Trithorax group proteins. Annu. Rev. Genet. 38, 413-443. Satoh, T., Takeuchi, O., Vandenbon, A., Yasuda, K., Tanaka, Y., Kumagai, Y., Miyake, T., Matsushita, K., Okazaki, T., Saitoh, T. et al. (2010). The Jmjd3-Irf4 axis regulates M2 macrophage polarization and host responses against helminth infection. Nat. Immunol. 11, 936-944. Scheuermann, J. C., Gutiérrez, L. and Müller, J. (2012). Histone H2A monoubiquitination and Polycomb repression: The missing pieces of the puzzle. Fly (Austin) 6, 162-168. Schmitges, F. W., Prusty, A. B., Faty, M., Stützer, A., Lingaraju, G. M., Aiwazian, J., Sack, R., Hess, D., Li, L., Zhou, S. et al. (2011). Histone methylation by PRC2 is inhibited by active chromatin marks. Mol. Cell 42, 330-341. Schuettengruber, B., Ganapathi, M., Leblanc, B., Portoso, M., Jaschek, R., Tolhuis, B., van Lohuizen, M., Tanay, A. and Cavalli, G. (2009). Functional anatomy of polycomb and trithorax chromatin landscapes in Drosophila embryos. PLoS Biol. 7, e13. Schwartz, Y. B. and Pirrotta, V. (2008). Polycomb complexes and epigenetic states. Curr. Opin. Cell Biol. 20, 266-273. Schwartz, Y. B., Kahn, T. G., Nix, D. A., Li, X. Y., Bourgon, R., Biggin, M. and Pirrotta, V. (2006). Genome-wide analysis of Polycomb targets in Drosophila melanogaster. Nat. Genet. 38, 700-705. Sedkov, Y., Benes, J. J., Berger, J. R., Riker, K. M., Tillib, S., Jones, R. S. and Mazo, A. (1999). Molecular genetic analysis of the Drosophila trithorax-related gene which encodes a novel SET domain protein. Mech. Dev. 82, 171-179. Seenundun, S., Rampalli, S., Liu, Q. C., Aziz, A., Palii, C., Hong, S., Blais, A., Brand, M., Ge, K. and Dilworth, F. J. (2010). UTX mediates demethylation of H3K27me3 at muscle-specific genes during myogenesis. EMBO J. 29, 1401-1411. Sengoku, T. and Yokoyama, S. (2011). Structural basis for histone H3 Lys 27 demethylation by UTX/KDM6A. Genes Dev. 25, 2266-2277. Shpargel, K. B., Sengoku, T., Yokoyama, S. and Magnuson, T. (2012). UTX and UTY demonstrate histone demethylase-independent function in mouse embryonic development. PLoS Genet. 8, e1002964. Smith, E. R., Lee, M. G., Winter, B., Droz, N. M., Eissenberg, J. C., Shiekhattar, R. and Shilatifard, A. (2008). Drosophila UTX is a histone H3 Lys27 demethylase that colocalizes with the elongating form of RNA polymerase II. Mol. Cell. Biol. 28, 1041-1046. Swigut, T. and Wysocka, J. (2007). H3K27 demethylases, at long last. Cell 131, 2932. Thieme, S., Gyárfás, T., Richter, C., Özhan, G., Fu, J., Alexopoulou, D., Muders, M. H., Michalk, I., Jakob, C., Dahl, A. et al. (2013). The histone demethylase UTX regulates stem cell migration and hematopoiesis. Blood 121, 2462-2473. Tie, F., Banerjee, R., Conrad, P. A., Scacheri, P. C. and Harte, P. J. (2012). Histone demethylase UTX and chromatin remodeler BRM bind directly to CBP and modulate acetylation of histone H3 lysine 27. Mol. Cell. Biol. 32, 2323-2334. Trojer, P. and Reinberg, D. (2006). Histone lysine demethylases and their impact on epigenetics. Cell 125, 213-217. Vandamme, J., Lettier, G., Sidoli, S., Di Schiavi, E., Nørregaard Jensen, O. and Salcini, A. E. (2012). The C. elegans H3K27 demethylase UTX-1 is essential for normal development, independent of its enzymatic activity. PLoS Genet. 8, e1002647. Wang, C., Lee, J. E., Cho, Y. W., Xiao, Y., Jin, Q., Liu, C. and Ge, K. (2012). UTX regulates mesoderm differentiation of embryonic stem cells independent of H3K27 demethylase activity. Proc. Natl. Acad. Sci. USA 109, 15324-15329. Welstead, G. G., Creyghton, M. P., Bilodeau, S., Cheng, A. W., Markoulaki, S., Young, R. A. and Jaenisch, R. (2012). X-linked H3K27me3 demethylase Utx is required for embryonic development in a sex-specific manner. Proc. Natl. Acad. Sci. USA 109, 13004-13009. White, R. A. and Wilcox, M. (1984). Protein products of the bithorax complex in Drosophila. Cell 39, 163-171. Yuan, W., Xu, M., Huang, C., Liu, N., Chen, S. and Zhu, B. (2011). H3K36 methylation antagonizes PRC2-mediated H3K27 methylation. J. Biol. Chem. 286, 7983-7989. DEVELOPMENT Utx function in Drosophila