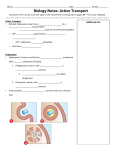
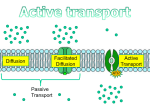
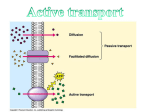
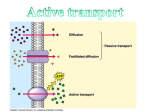
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
G protein–coupled receptor wikipedia , lookup
Cell membrane wikipedia , lookup
Cytokinesis wikipedia , lookup
Green fluorescent protein wikipedia , lookup
Hedgehog signaling pathway wikipedia , lookup
SNARE (protein) wikipedia , lookup
Protein phosphorylation wikipedia , lookup
Signal transduction wikipedia , lookup
Protein moonlighting wikipedia , lookup
Endomembrane system wikipedia , lookup
Protein–protein interaction wikipedia , lookup
606 Biochemical Society Transactions (2005) Volume 33, part 4 A role of SAND-family proteins in endocytosis D. Poteryaev1 and A. Spang Friedrich Miescher Laboratory of the Max Planck Society, Spemannstrasse 39, Tuebingen D-72076, Germany Abstract Caenorhabditis elegans has recently been used as an attractive model system to gain insight into mechanisms of endocytosis in multicellular organisms. A combination of forward and reverse genetics has identified a number of new membrane trafficking factors. Most of them have mammalian homologues which function in the same transport events. We describe a novel C. elegans gene sand-1, whose loss of function causes profound endocytic defects in many tissues. SAND-1 belongs to a conserved family of proteins present in all eukaryotic species, whose genome is sequenced. However, SAND family has not been previously characterized in metazoa. Our comparison of C. elegans SAND-1 and its yeast homologue, Mon1p, showed a conserved role of the SAND-family proteins in late steps of endocytic transport. Identification of new endocytosis genes in multicellular organisms Endocytosis is a key process conserved in all eukaryotes. It is essential for the uptake of extracellular nutrients and the regulation of membrane dynamics. Furthermore, endocytosis participates in the cell’s reaction to extracellular stimuli by desensitizing, down-regulating or recycling receptors and membrane proteins [1]. Genetic screens for effectors of endocytosis have been carried out successfully in the yeast Saccharomyces cerevisiae, yielding a number of genes which participate in this process [1]. Many of them have homologues in multicellular organisms, whose functions are similar to or identical with their yeast counterparts. However, the greater complexity of higher eukaryotes, compared with yeast, suggests the existence of many variations in endocytic transport in different cells and tissues. This can be exemplified by the Ypt/Rab family of small GTPases. The yeast genome contains 11 members of the Ypt family, nine of which were shown to participate in different steps of vesicular transport pathways, including endocytosis. The human Rab family comprises about 60 members, of which only ten are ubiquitous and have clear functional homologues in yeast. The other Rabs are believed to be required for either specialized cell functions or only at certain developmental stages [2]. Forward genetics in higher eukaryotes has been most successful in the two model organisms Drosophila melanogaster and Caenorhabditis elegans. Over the past few years, two screens for endocytosis-defective mutants in C. elegans have been performed [3]. The screens employed two endocytic models: receptor-mediated endocytosis of yolk Key words: Caenorhabditis elegans, endocytosis, Rab7, receptor-mediated endocytosis defective (rme) screen, SAND family, vacuole. Abbreviations used: CPY, carboxypeptidase Y; cup, coelomocyte uptake defective; GFP, green fluorescent protein; HOPS, homotypic fusion and vacuolar protein sorting complex; MLIV, mucolipidosis type IV; rme, receptor-mediated endocytosis defective; SNARE, soluble Nethylmaleimide-sensitive fusion protein attachment protein receptor; ssGFP, secreted soluble GFP. 1 To whom correspondence should be addressed (email dmitry.poteryaev@tuebingen. mpg.de). C 2005 Biochemical Society in oocytes [rme (receptor-mediated endocytosis defective) screen] and fluid-phase endocytosis by specialized scavenger cells residing in the worm’s body cavity, called coelomocytes [cup (coelomocyte uptake defective) screen]. Yolk uptake by maturing oocytes is an example of the receptor-mediated endocytosis pathway in many species. Yolk storage vesicles are thought to be the oocyte equivalent of late endosomes or lysosomes but with low proteolytic activity. A C. elegans strain carrying a yolk protein YP170 fusion to GFP (YP170–GFP, where GFP stands for green fluorescent protein) has been a useful tool to study receptor-mediated endocytosis and helped to characterize a number of mutants affecting this pathway [4]. Among the novel conserved genes identified in the rme screen were rme-1 and rme-8 (Figure 1). RME-1 is a member of a conserved class of EH (Eps15-homology)-domain proteins and is associated with the periphery of endocytic organelles, suggesting a direct role in endocytic transport. Evidence from studies of nematode and mammalian RME-1 established a function for RME-1 in recycling, specifically in the exit of membrane proteins from recycling endosomes. These studies have shown a conserved function in endocytic recycling for the RME-1 family of EH proteins [5]. Another gene, rme-8, whose loss of function causes the RME phenotype in C. elegans [6], has been later studied in Drosophila. Analysis of Drosophila Rme-8 mutants showed that internalization of receptors and the uptake of tracers were blocked. Rme-8 has been then shown to participate in clathrin uncoating, by interaction with Hsc704 through its J-domain. Thus Rme-8 appears to function as an unexpected but critical co-chaperone with Hsc70 in endocytosis [7]. The C. elegans cup screen was based upon monitoring the uptake of ssGFP (secreted soluble GFP) from the body cavity by coelomocytes. The most remarkable gene identified so far in the cup screen is cup-5. Although cup-5 mutants are not defective in the initial uptake, the endocytic tracers like ssGFP are not degraded in lysosomes. Instead, they accumulate in large vacuoles that are positive for both late endosomal and lysosomal markers. CUP-5 is the C. elegans functional orthologue of human mucolipin-1 Localization and Activation of Ras-like GTPases Figure 1 A model of endocytic traffic and functions of endocytic effectors discovered in C. elegans RAB-5, RAB-11 and RAB-7 participate in early endosome (EE) formation, recycling from endocytic recycling compartment and EE to late endosome (LE) transport respectively. The major endocytic traffic route to lysosomes from plasma membrane-coated pits is shown, incorporating present views [9] of lysosome–LE fusion and re-formation of lysosomes from the resulting hybrid organelles. CUP-5 might control the process of lysosome re-formation by regulating the calcium flux. SAND-1 is thought to function at the EE to LE step. CCV, clathrin-coated vesicle. [8]. Mutations in mucolipin-1 cause MLIV (mucolipidosis type IV), a disease characterized by psychomotor retardation and ophthalmological abnormalities. In MLIV patients, cells of several tissues accumulate large vacuoles that are presumed to be abnormal lysosomes. CUP-5 is a calcium channel and functions in the biogenesis of lysosomes originating from the so-called hybrid organelles (Figure 1) [8,9]. Late endosomes dock and fuse with lysosomes in a SNARE (soluble Nethylmaleimide-sensitive fusion protein attachment protein receptor)-dependent reaction that requires the efflux of calcium from lysosomes or endosomes. The fusion might occur either in a restricted manner (kiss-and-run) or in full, resulting in an endosome–lysosome hybrid organelle. According to the present view, late-endosome components are sorted into discrete subdomains and separated from hybrid organelles to enable the re-formation of lysosomes and the retrieval of late-endosome components. Calcium- and pHdependent condensation of contents facilitates lysosome reformation [9]. Remarkably, most of the genes identified in these studies, while having functional homologues in vertebrates, are not present in yeast. This stresses the importance of using metazoan model organisms, such as C. elegans, for which powerful genetic methods are available, in deciphering endocytosis. Clearly, there are many unanswered questions, and new effectors of endocytosis are awaiting discovery. SAND family and endocytosis The C. elegans mutant allele or552 was identified in a screen for temperature-sensitive embryonic lethal mutations [15]. Detailed analysis of the or552 phenotype led us to speculate that the affected gene is involved in late steps of endocytic transport and probably acts on the same level as rab-7. We found that or552 is a nonsense mutation in a gene that belongs to the SAND family. According to the C. elegans nomenclature it was named sand-1. SAND was first identified as an open reading frame in the yeast S. cerevisiae genome. An orthologous gene was found later in Fugu rubripes and was called SAND. By now, it is clear that SAND genes are present in every major eukaryotic taxon. Fungi, plasmodia, slime mould, nematodes, sea squirt and plants all have one SAND protein encoded in their genome. Interestingly, in vertebrate species, including humans, two SAND genes are always present. Sequence analysis of the SAND family has failed to detect any significant similarities with known protein domains. Therefore the SAND proteins are ancient and belong to an absolutely distinct protein family. The most conspicuous feature of sand-1 (or552) mutants is the presence of large intracellular membrane-bounded granules in the early embryo. These granules are a result of defective endocytic transport of yolk protein in maturing oocytes (Figure 2A). The fact that at permissive temperature the mutant animals survive, although still having defects in endocytosis, allowed us to investigate the role of sand-1 also in somatic tissues. In coelomocytes we observed a similar defect: accumulation of extremely large vacuoles of endocytic origin (Figure 2B). The only member of the SAND family whose biological function has been investigated is the budding yeast gene MON1 [10]. Mon1p is involved in autophagy, vacuolar morphology, cytoplasm-to-vacuole trafficking, and is required for homotypic vacuole fusion. In other words, Mon1p is required in nearly all membrane trafficking pathways, where the vacuole represents the terminal acceptor compartment. Biochemical experiments in budding yeast have identified four distinct stages in homotypic vacuole fusion: priming of the vacuoles, where cis-SNARE complex disassembles and the C-Vps/HOPS (homotypic fusion and vacuolar protein sorting complex) is released from the SNAREs; tethering, which requires the interaction of the C-Vps/HOPS complex with the rab GTPase Ypt7, and, finally, docking and fusion. Mon1p forms a stable complex with another protein, Ccz1p. This complex is required for homotypic vacuole fusion at tethering/docking stage; however, only Mon1p plays a direct role, as determined by antibody inhibition assays [11]. The complex of Ccz1p and Mon1p is essential for SNARE pair association during homotypic vacuole fusion and, probably, in other trafficking pathways to the vacuole [11,12]. Apparently, Ypt7p, Mon1p and Ccz1p act on the same level. The deletion phenotypes of these genes are highly similar, regarding the vacuole integrity and membrane transport. Moreover, overexpression of YPT7 completely suppressed the effect of a CCZ1 deletion [13]. Whether Ypt7p and either Mon1p or Ccz1p interact physically remains an open C 2005 Biochemical Society 607 608 Biochemical Society Transactions (2005) Volume 33, part 4 Figure 2 Conserved role of sand-1/MON1 in endocytosis (A) Abnormal receptor-mediated endocytosis in sand-1 (or552) mutants. Cytoplasm of an early embryo contains enlarged yolk granules. (B) Coelomocytes in sand-1 (or552) accumulate large vacuoles of endocytic origin. Uptake of BSA–Texas Red injected into the body cavity is shown. (C) Expression of C. elegans sand-1 in a yeast strain deficient for MON1 can restore the vacuole targeting. CPY maturation assay is shown. Arrows indicate: p1, CPY precursor in ER; p2, glycosylated CPY precursor in Golgi or pre-vacuolar compartment; m, mature form in the vacuole. The time of chase after the pulse is shown in min. question. Kucharczyk et al. [13] reported the interaction of Ccz1p with Ypt7p, whereas Wang et al. [11,12] were not able to reproduce these results. They also failed to detect an interaction between Mon1p and Ypt7p [11,12]. Still, it may be possible that these interactions are transient, whereas the Ccz1p–Mon1p complex is very abundant and stable. It is therefore tempting to suggest that Ccz1p and Mon1p are part of an Ypt7p effector complex. It should be noted that Ccz1p has not been found in genomes of any other eukaryotic species. Therefore this association is not evolutionarily conserved. Moreover, it is conceivable that in distant eukaryotic taxons, SAND-interacting proteins evolved independently and are different. To date, only one metazoan SAND has been experimentally studied. Human HSRG1 has been identified as a protein whose expression is induced at high levels in fibroblasts by HSV-1 (herpes simplex virus 1) infection [14]. Although these experiments did not go any further, one is tempted to speculate about an involvement of HSRG1 in endocytosis and a possible link of endocytic transport and HSV-1 infection. We propose that sand-1 may be a functional orthologue of yeast MON1. To test this hypothesis, we tried to complement the protein transport defect of ∆mon1. CPY (carboxypeptidase Y) is transported to the vacuole through the endoplasmic reticulum, Golgi complex and endosome. Because CPY is post-translationally modified, its transport through the secretory pathway is easily followed. In the wildtype strain, CPY matured with a half-time of 5–10 min. In ∆mon1, the majority of the protein was found in the precursor forms even after 40 min of chase, which is in agreement with previous findings [11]. The delay in CPY maturation resulted from a defective cargo transport from the pre-vacuolar compartment to the vacuole. The ∆mon1 strain transformed with C 2005 Biochemical Society sand-1 showed a marked increase in the kinetics of CPY maturation. After a 30-min chase, more than half of the CPY was in the mature form (i.e. reached the vacuole) (Figure 2C). Thus overexpression of sand-1 can at least partially rescue the function of its yeast orthologue. We have shown here that the C. elegans SAND-1 is required for late steps of endocytosis in oocytes, embryos, coelomocytes and, possibly, in many other tissues. Furthermore, the C. elegans SAND-1 is the functional orthologue of the S. cerevisiae Mon1p, suggesting a conserved role of these proteins in different systems. References 1 D’Hondt, K., Heese-Peck, A. and Riezman, H. (2000) Annu. Rev. Genet. 34, 255–295 2 Segev, N. (2001) Science STKE 100, RE11 3 Fares, H. and Grant, B. (2002) Traffic 3, 11–19 4 Grant, B. and Hirsh, D. (1999) Mol. Biol. Cell 10, 4311–4326 5 Grant, B., Zhang, Y., Paupard, M.C., Lin, S.X., Hall, D.H. and Hirsh, D. (2001) Nat. Cell Biol. 3, 573–579 6 Zhang, Y., Grant, B. and Hirsh, D. (2001) Mol. Biol. Cell 12, 2011–2021 7 Chang, H.C., Hull, M. and Mellman, I. (2004) J. Cell Biol. 164, 1055–1064 8 Treusch, S., Knuth, S., Slaugenhaupt, S.A., Goldin, E., Grant, B.D. and Fares, H. (2004) Proc. Natl. Acad. Sci. U.S.A. 101, 4483–4488 9 Pipera, R.C. and Luzio, J.P. (2004) Trends Cell Biol. 14, 471–473 10 Muren, E., Oyen, M., Barmark, G. and Ronne, H. (2001) Yeast 18, 163–172 11 Wang, C.W., Stromhaug, P.E., Kauffman, E.J., Weisman, L.S. and Klionsky, D.J. (2003) J. Cell Biol. 163, 973–985 12 Wang, C.W., Stromhaug, P.E., Shima, J. and Klionsky, D.J. (2002) J. Biol. Chem. 277, 47917–47927 13 Kucharczyk, R., Kierzek, A.M., Slonimski, P.P. and Rytka, J. (2001) J. Cell Sci. 114, 3137–3145 14 Dong, S., Dong, C., Liu, L., Che, Y., Sun, M., Hu, F., Li, J. and Li, Q. (2003) Acta Virol. 47, 27–32 15 Swan, K.A., Severson, A.F., Carter, J.C., Martin, P.R., Schnabel, H., Schnabel, R. and Bowerman, B. (1998) J. Cell Sci. 111, 2017–2027 Received 9 March 2005