Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
X-inactivation wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Epigenetics in stem-cell differentiation wikipedia , lookup
Genomic imprinting wikipedia , lookup
Epigenetics in learning and memory wikipedia , lookup
Epigenetics of diabetes Type 2 wikipedia , lookup
Designer baby wikipedia , lookup
Genome (book) wikipedia , lookup
Microevolution wikipedia , lookup
Gene therapy of the human retina wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Long non-coding RNA wikipedia , lookup
Primary transcript wikipedia , lookup
Gene expression profiling wikipedia , lookup
Genomic library wikipedia , lookup
Gene expression programming wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Mir-92 microRNA precursor family wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
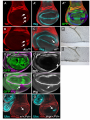
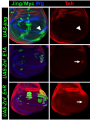
Genetics: Published Articles Ahead of Print, published on March 1, 2006 as 10.1534/genetics.106.056341 Culi et al. jing is required for wing development and to establish the proximo-distal axis of the leg in Drosophila melanogaster Joaquim Culi*,†, Pilar Aroca†,1, Juan Modolell† and Richard S. Mann* * Department of Biochemistry and Molecular Biophysics, Columbia University, New York, NY, USA, † Centro de Biología Molecular Severo Ochoa, CSIC and Universidad Autónoma de Madrid, 28049 Madrid, Spain. 1 Current address: Dpto. Anatomía Humana y Psicobiología; Facultad de Medicina; Universidad de Murcia; 30100 Murcia, Spain. 1 Culi et al. Running title: Role of jing in appendage development Key words: proximo-distal axis, leg development, alula development, Zn-finger repressor, homothorax, teashirt Corresponding author: Richard S. Mann [email protected] 212-305-7731 (phone) 212-305-7924 (fax) 2 Culi et al. ABSTRACT The establishment of the proximo-distal (PD) axis in the legs of Drosophila melanogaster requires the expression of a nested set of transcription factors that are activated in discreet domains by secreted signaling molecules. The precise regulation of these transcription factor domains is critical for generating the stereotyped morphological characteristics that exist along the PD axis, such as the positioning of specific bristle types and leg joints. Here we provide evidence that the Zn-finger protein encoded by the gene jing is critical for PD axis formation in the Drosophila legs. Our data suggest that jing represses transcription and that it is necessary to keep the proximal gene homothorax (hth) repressed in the medial domain of the PD axis. We further show that jing is also required for alula and vein development in the adult wing. In the wing, Jing is required to repress another proximal gene, teashirt (tsh), in a small domain that will give rise to the alula. Interestingly, we also demonstrate that two other genes affecting alula development, Alula and elbow, also exhibit tsh derepression in the same region of the wing disc as jing– clones. Finally, we show that jing genetically interacts with several members of the Polycomb (Pc) group of genes during development. Together, our data suggest that jing encodes a transcriptional repressor that may participate in a subset of Pc-dependent activities during Drosophila appendage development. INTRODUCTION The development of animal appendages requires the establishment of a proximo-distal (PD) axis, which extends orthogonally from the trunk. Unlike the antero-posterior (AP) and dorso-ventral (DV) axes, which exist at all stages of 3 Culi et al. development, the PD axis must be created de novo for each appendage during embryogenesis. Many insights into PD axis formation have been obtained by studying its establishment in Drosophila melanogaster (reviewed by (COUSO and BISHOP 1998; MORATA 2001). Cells that will give rise to the leg can be first visualized as a small circular ventral domain in each of the three thoracic hemisegments during stage 9 of embryogenesis. A marker for the leg primordia at this early stage is the homeobox gene Distal-less (Dll), which is also required for distal leg development (CAMPBELL and TOMLINSON 1998; COHEN 1990; COHEN et al. 1989). Interestingly, a subset of the cells that initially express Dll at this stage will migrate dorsally and contribute to the wing and haltere primordia, two dorsal appendages that grow from the second and third thoracic segments, respectively. Early expression of Dll in the leg primordia is activated by Wingless (Wg) signaling and constrained by two other pathways, Decapentaplegic (Dpp) signaling dorsally and Epidermal growth factor receptor (DER) signaling ventrally (COHEN et al. 1991; COHEN 1990; GOTO and HAYASHI 1997). In addition, Dll is repressed by the Hox genes in the abdomen, thus limiting its expression in the ectoderm to the thoracic segments (GEBELEIN et al. 2002; VACHON et al. 1992). Although there is no clear morphological PD axis at this stage, a subset of the Dll-expressing cells will contribute to the distal leg segments while more proximal leg segments appear to arise from cells immediately ventral to the Dllexpression domain (BOLINGER and BOEKHOFF-FALK 2005; GOTO and HAYASHI 1997). These proximally-fated cells express the genes homothorax (hth), teashirt (tsh), and escargot (esg). Late in embryogenesis and extending into larval development, the leg PD axis is elaborated by Wg and Dpp, which induce the expression of downstream transcription factors at different positions along the PD axis in the leg imaginal discs 4 Culi et al. (ABU-SHAAR and MANN 1998; LECUIT and COHEN 1997). During this stage, Wg and Dpp are both expressed adjacent to the AP compartment boundary, but in mutually exclusive ventral and dorsal stripes, respectively (JIANG and STRUHL 1996; PENTON and HOFFMANN 1996; THEISEN et al. 1996). By mechanisms that are still not well understood, high levels of both Dpp and Wg are required to activate Dll expression while lower levels of the same two signals activate another transcription factor, dachshund (dac) (LECUIT and COHEN 1997). The result of this Wg+Dpp signaling is the formation of three domains of gene expression along the PD axis: Dll in distal cells, dac in medial cells, and hth/tsh in proximal cells (ABU-SHAAR and MANN 1998; LECUIT and COHEN 1997). By approximately 48 hrs of development, both Dll and dac become independent of Wg and Dpp. This change in regulation, together with mutually antagonistic interactions between these transcription factors, results in a more complex pattern of gene expression along the PD axis that includes cells that express combinations of these transcription factors in addition to cells that express only Dll or only Dac. Finally, a secondary patterning event is triggered by the Wg+Dpp-dependent expression of ligands of the EGFR pathway in a small number of distal-most cells (CAMPBELL 2002; GALINDO et al. 2005; GALINDO et al. 2002). The activation of this pathway in a graded manner induces another set of transcription factors that subdivide and pattern the distal region of the leg. Thus, the leg PD axis is established by at least two sets of secreted signals that each induce the expression of a nested set of downstream transcription factors. A combinatorial code of these transcription factors establishes specific cell fate domains along the PD axis. One of the outputs of this code that has been examined is the formation of the leg joints in a process that depends on the activation of the Notch pathway (BISHOP et al. 1999; RAUSKOLB 2001; RAUSKOLB and IRVINE 1999). 5 Culi et al. Like most transcription factors, those that are required for PD axis formation in the Drosophila leg have additional and very distinct functions at other stages and tissues during development. For example, dac plays an important role in the development of the eye, central nervous system (CNS), and trachea (MARDON et al. 1994; MARTINI et al. 2000; SHEN and MARDON 1997). In addition to its role in ventral appendage formation, Dll is also expressed and likely required in the CNS and plays an important role in antennal development (CASARES and MANN 1998; DONG et al. 2000; DUNCAN et al. 1998; PANGANIBAN 2000). hth, in addition to specifying proximal identities in the leg, is also required for proximal wing (hinge) development, antennal and eye development, and, together with its partner gene extradenticle (exd), encodes a cofactor for the Hox proteins (ALDAZ et al. 2005; AZPIAZU and MORATA 2000; CASARES and MANN 1998; CASARES and MANN 2000; DONG et al. 2000; MANN and AFFOLTER 1998; PICHAUD and CASARES 2000). tsh also has roles in multiple tissues, including the eye, wing, and leg (BESSA and CASARES 2005; WU and COHEN 2000; WU and COHEN 2002). These observations raise the general question of how transcription factors execute their specific functions in vivo. The prevailing model to account for specificity is that transcription factors collaborate with each other to create unique combinations that have distinct properties that are not simply the sum of their individual properties. Although there is already evidence to support this idea in the leg, the number of combinations of factors currently known in the leg is still not sufficient to account for the many unique identities along the PD axis. Accordingly, we would expect there to be additional factors that contribute to PD axis specification. Here, we identify the gene jing as a factor that is required for forming the PD axis of the leg. We identified jing in a genetic screen to find modifiers of a hth haploinsufficiency phenotype. jing encodes a Zn finger transcription factor that has 6 Culi et al. previously been implicated in playing a role in oogenesis and CNS and trachea development (LIU and MONTELL 2001; SEDAGHAT et al. 2002; SEDAGHAT and SONNENFELD 2002; SONNENFELD et al. 2004). However, a role in adult leg development has not been previously described. Using clonal analysis, we show that jing is an essential factor for specifying medial fates in the leg. We further show that jing also plays a highly specific role in wing development. Together, these results suggest that jing may function together with previously characterized transcription factors involved in these processes to help them execute their functions in vivo. MATERIALS AND METHODS Drosophila genetics Most of the Drosophila stocks used in this work, including the deficiency set for the second chromosome, are described in FlyBase and Bloomington Stock Center web sites. jing22F3 and jing47H6 alleles were obtained from Denise Montell's lab (LIU and MONTELL 2001). Since these two mutations were not molecularly characterized, we sequenced the jing genomic region from homozygote single embryos of jing22F3 genotype. Our analysis showed that in the jing22F3 allele, the codon coding for K664 (amino acids according to Jing isoform with Gene bank accession number AAK49526.1) was mutated to a premature stop codon. Since K664 is located 5' of the DNA binding Zn finger motifs, this allele probably codes for a non-functional protein. To generate clones of jing- cells, we used the FLP/FRT system (XU and RUBIN 1993). jing22F3 and jing47H6 alleles were generated over an FRTG13 chromosome (LIU and MONTELL 2001). Thus, to analyze the clones in imaginal discs, we crossed hsFlp; FRTG13, ubi-GFP flies with the jing stocks. The resulting larvae were heatshocked for one hour at 37 C at 24-48 hr after egg laying and allowed to developed at 7 Culi et al. 25 C. 3th instar larval imaginal discs were dissected and analyzed by immunostaining. jing- cells can be identified by the absence of GFP expression. To analyze adult phenotypes, jing- clones were similarly generated in a f36a background. The mutant cells were marked by the loss of f rescue provided by two f+ transgenes (P{f+13}44C, P{f+13}47B , a gift from P. Martín, Antonio García-Bellido's lab) which were recombined with FRTG13. Adult legs were dissected and mounted in Hoyer's/lactic media. Clones of jing22F3 that also express UAS-Jing FL, UAS-Jing_ZnfEnR or UASJing_ZnfE1A were obtained using MARCM (LEE and LUO 2001). Flies of the genotype y,w,hs-Flp; FRTG13, jing22F3 /CyO; tubulin-Gal4/TM2 were crossed to w; FRTG13, piMyc, tubulin-Gal80/CyO; UAS-Jing FL (or its variants) and the resulting larvae heatshocked as above. To check for a genetic interaction between jing and PcG genes, we used the stocks PcXT109,FRT2A/TM6C and FRTG13,Df(2)vgD, Asxxf23, PclXM3/CyO (obtained from A. Busturia's lab). Oregon R and jing22F3/Cyo females were crossed with males from these two stocks and allowed to developed in non-crowded conditions at 25 C. Only female progeny was scored. Immunohistochemistry To generate an anti-Jing antibody, we expressed a 290 amino acids peptide corresponding to the N-terminal part of Jing protein (AAK49526.1) as a GST fusion. The protein was expressed in E. coli and purified from the soluble fraction by affinity chromatography. Rabbits were immunized at Cocalico Biologicals. The resulting antibodies did not allow us to detect endogenous Jing protein. However, they can detect overexpressed Jing. Other antibodies used in this work are: anti-Hth (CASARES 8 Culi et al. and MANN 1998), anti-Tsh (from SK Chan), anti-Dll (COHEN et al. 1993), anti-Dac (Developmental Studies Hybridoma Bank, DSHB), anti-Wg (4D4; DSHB), anti-Ubx (FP3.38), and anti-myc (Mab 9E10, Babco). To detect beta-galactosidase activity, imaginal discs were dissected in PBS, fixed in 1% glutaraldehyde for 2 minutes on ice, washed twice with PBS and stained with 0.2% X-gal overnight at 37 C. In situ hybridization to detect jing transcripts in imaginal discs was carried out with an antisense RNA probe derived from the Zn finger region using standard procedures (details can be provided upon request). cDNA identification and construction of transgenes. To identify the transcripts encoded in the proximity of P1094 insertion site, we screened an imaginal disc cDNA library constructed in λgt10 using a fragment of genomic DNA adjacent to the P element insertion site as probe. We identified four independent partial clones that were grouped into two non-overlapping families. Our cDNAs, together with additional cDNAs identified by BDGP, suggest the existence of at least four different jing transcripts that are generated through the use of alternative promoters and alternative splicing (Supplementary Fig. 1). Three of these transcripts only differ in their 5'UTRs and thus, code for the same protein. In contrast, a fourth larger transcript of 7 Kb includes sequences from a previously identified gene named 1.28, which is located about 110 Kb upstream of jing (MAHAFFEY et al. 1993). At least two of the four cDNAs isolated from the imaginal disc library correspond to this larger 7 Kb transcript. Thus, our data indicate that 1.28 and jing are a single gene, which we suggest should retain the name jing. To generate UAS-Jing FL, we cloned the coding region from the 7 kb transcript into pUAST (BRAND and PERRIMON 1993). UAS-Jing_Znf, UAS- 9 Culi et al. Jing_ZnfEnR and UAS-Jing_ZnfE1A were generated by cloning the Jing Zinc finger region (from A119 to I1288; Jing protein AAK49526.1) in frame into the vectors pCS2-MT-NLS, pCS2-MT-NLS-EnR and pCS2-MT-NLS-E1A respectively (GLAVIC et al. 2002). Note that a nuclear localization signal, as well as six myc tags were included in these transgenes. Afterwards, the constructs were transferred to pUAST for P element transformation and expression in Drosophila. RESULTS Genetic interaction between jing and homothorax To identify additional genes that may function during the same processes as hth during development, we carried out a genetic modifier screen in Drosophila. This screen was based on the observation that some alleles of hth display a dominant phenotype in the adult flies, in particular, a broadening and increase in pigmentation of male fourth abdominal segment (A4) (RIECKHOF et al. 1997). This change in pigmentation reflects a partial transformation of the lightly pigmented A4 towards a darker A5. Different hth alleles have variable degrees of transformation, suggesting that this pigmentation phenotype is sensitive to gene dosage and possibly to the action of modifiers. In the screen we crossed hthk1, an allele exhibiting a strong and highly penetrant increase in pigmentation (Fig. 1B), to a series of overlapping deficiencies of the second chromosome. This set of deficiencies uncovers approximately 85% of the genes on that chromosome. We identified one deficiency, Df(2R)cn88b that, as a heterozygote, resulted in a strong widening of the A4 pigmentation when crossed to hthk1 (data not shown). On its own, this deficiency does not show an increase in abdominal pigmentation. Based on these observations, we mapped the gene 10 Culi et al. responsible for the enhancement of the hthk1 phenotype by testing lethal P element insertions that map to this deficiency. These crosses lead to the identification of two insertions, P{PZ}jing01094 and P{lacW}jingK03404, that also enhanced the hthK1 tergite phenotype (Fig. 1 and data not shown). Both of these P elements are inserted into the gene jing, which codes for a Zn-finger transcription factor. We confirmed that the interaction between hthk1 and these P elements was caused by a decrease in jing activity by analyzing an EMS induced allele, jing22F3, which has a premature stop codon N-terminal to the Zn-finger DNA binding domain (Methods). As with the P insertions, jing22F3 increased A4 pigmentation in hthK1 males (Fig. 1C), confirming that jing and hth genetically interact in this assay. A role for jing in the establishment of the PD axis in the leg To analyze further the relationship between hth and jing, we characterized jing’s role in adult development. Because strong alleles of jing are lethal when homozygous, we performed a clonal analysis using the alleles jing22F3 and jing47H6, two strong and possibly null alleles that gave similar phenotypes. Clones of cells homozygous for these jing alleles in the abdomen show a strong decrease in pigmentation, a phenotype that was not further analyzed (data not shown). In the leg, jing clones have profound effects on development. Clones located in the most proximal segments of the leg, the coxa and the trochanter, do not have any observable phenotype. However, clones located in the femur, tibia or first tarsal segment display the loss of bracts located at the base of the bristles. This suggests a transformation towards a more proximal fate, because bracted bristles are characteristic of the distal regions of the leg (Fig. 2). We also observed fusions between the femur and the tibia and between the tibia and the first tarsal segment. In these cases we always detected 11 Culi et al. jing– tissue in the fusion area. These fusions are usually associated with a thickening of the affected segments (Fig. 2D,E). In some legs, we also observed small outgrowths and internalized vesicles that seem to have detached from the leg cuticle (Fig. 2B and not shown). We never observed a phenotype in the last four tarsal segments. However, it is difficult to unambiguously identify clones in this region of the legs because of limitations with the marker used to identify the jing- cells (forked; f). Taken together, the leg phenotypes suggest that jing participates in the establishment of the PD axis of the legs and, more specifically, that jing is required for the establishment of intermediate fates along the PD axis, a similar domain to that governed by dac (Fig. 2). In contrast, hth and tsh are required for the establishment of proximal (coxa and trochanter) fates. Thus, jing and exd/hth are required in mostly complementary domains of the leg. jing is required for wing vein and alula development In the wing, jing– clones display two different phenotypes. In the wing blade, the clones are generally associated with a decrease or absence of wing vein differentiation (Fig. 3). However, this phenotype occurs with a low penetrance and not all veins are equally sensitive to the loss of jing. The distal portions of longitudinal veins 4 and 5 are very sensitive, as well as both cross-veins. The phenotype is stronger when the jing– clones affect both the dorsal and ventral aspects of the same vein. Interestingly, some clones adjacent but not including a longitudinal vein induce the non-autonomous elimination of that vein. Occasionally, we also observed the ectopic differentiation of a vein abutting a jing– clone (Fig. 3C). Clones located at the wing margin moderately disorganize the regular arrangement of bristles 12 Culi et al. and can induce formation of a few extra-bristles, in particular at the distal longitudinal vein 3 (not shown). A second phenotype we observed in the wing is an absolute requirement of jing for the differentiation of the alula, also known as the alar lobe (FlyBase). Clones located in the alula cause a reduction in its size and its fusion with the main portion of the wing blade (Fig. 4B). Other phenotypes associated with jing– tissue in the alula region are pigmented vesicles inside the wing (Fig. 4B, inset) and the formation of small outgrowths (Fig. 4C). We also observed detached vesicles and outgrowths in the hinge region of the wing (Fig. D and E). However, in this region it is not possible to identify the f marker. jing is required to repress hth expression in the medial leg To better analyze the role of jing during PD axis formation in the leg, we generated clones of jing– cells during the first instar larval stage and analyzed the expression of several PD markers in 3rd instar leg imaginal discs. In particular, we focused on Tsh, Hth, Dac, and Dll, four transcription factors expressed in partially overlapping and concentric domains that are essential for the establishment of the leg PD axis. In agreement with the adult jing– phenotypes, clones located in the proximal hth/tsh domain, which gives rise to the coxa and trochanter, do not show any change in the expression of these PD markers (Fig. 5C, D, E and not shown). In contrast, clones located in the intermediate dac region express hth, which is usually restricted to the proximal region of wild type discs (Fig. 5C and D). However, in some jing– clones Hth expression is weak and is not always seen in all cells of the clone. Further, we note that ectopic expression of full-length Jing does not repress hth in the leg, suggesting that it must work with other factors to repress hth (not shown). To confirm 13 Culi et al. that the Hth derepression was caused by the elimination of jing and not by some other associated mutation in the chromosome, we used the MARCM system (LEE and LUO 2001) to re-supply full length Jing in the jing– clones. In these clones, no ectopic Hth expression was detected (Fig. 5F). In addition to Hth derepression, jing– clones located a few cell diameters distal to the Tsh expression domain show Tsh derepression (Fig. 5E). Clones located distal to the Dac domain did not show any change in the expression of these four markers. jing– clones were recovered in all regions of the leg discs, including the most distal segments corresponding to the presumptive tarsi. Because we never observed adult phenotypes in the most distal four tarsi (see above), it is likely that jing is not required in this domain. However, we observed ectopic Hth is in distally positioned jing– clones in the antenna, suggesting that jing is also required in this disc to maintain hth repression (not shown). Although ectopic expression of Hth can repress Dll and dac (ABU-SHAAR and MANN 1998), we have not detected any consistent change in the expression of Dll or dac in jing– clones, perhaps because Hth derepression in these clones is not high enough (Fig. 5 and data not shown). Using in situ hybridization we found that jing is broadly expressed throughout the leg and wing discs (Fig. 5A, B). The expression of a lacZ insertion into jing (P{PZ}jing01094) also suggests that it is widely expressed throughout imaginal disc development (data not shown). jing is required for the repression of tsh, but not hth, in the alula region of the wing imaginal disc The analysis of jing loss of function clones in the adult indicates that this gene is essential for the differentiation of the alula. The development of the alula is a poorly defined process. The transcription factors of the Iroquois complex (Iro-C) are 14 Culi et al. required for the formation of the alula (GOMEZ-SKARMETA et al. 1996; KEHL et al. 1998). However, no change in the expression of Araucan, one the Iro-C genes, was observed in jing- clones in the alula region of 3rd instar imaginal discs (data not shown). Prompted in part by our finding that some jing– clones in the leg show derepression of hth and tsh, we examined the expression of these factors in jing– clones in wing imaginal discs. Surprisingly, we observed strong derepression of Tsh in jing– clones. This derepression is restricted to a very specific area of the wing disc that corresponds to the region that gives rise to the alula (Fig. 6A). Since there is no well-defined genetic marker for the alula, we resorted to the use of an antibody against Wg, which is expressed along the D/V boundary and also in two concentric rings in the wing pouch. The inner (more distal) Wg ring bisects the alula, as shown by the expression of wg-lacZ in adult wings (NEUMANN and COHEN 1996). Thus, using Wg as a marker, we mapped the region were Tsh is derepressed to a small area close to the intersection of the DV boundary with the inner Wg ring and only in the posterior compartment (Fig. 6A). Tsh derepression always occurs distal to the inner Wg ring and in both ventral and dorsal compartments. Using the P{PZ}jing01094 enhancer trap, we detected a weak expression of jing-lacZ in most cells of the wing disc (not shown). Interestingly, cells at the DV boundary, where Wg is highly expressed, are refractory to Tsh derepression. This result is in agreement with previous reports indicating that Wg represses tsh expression (WEIHE et al. 2004; WU and COHEN 2002; ZIRIN and MANN 2004). In contrast to these effects on tsh, hth is expressed in the alula region of wild type discs and appears to be expressed normally in jing– clones. In the wing pouch, a region devoid of hth expression, jing– clones show very weak derepression of hth (Fig. 6D). However, this level of derepression appears to have no developmental 15 Culi et al. consequences, because, as described above, adult wings with jing– clones do not have PD abnormalities (Fig. 3). Intrigued by the strong de-repression of tsh in jing– clones located in the alula region, we determined if tsh is also derepressed in two classical mutations that show a reduction or absence of the alula, Alula (Alu) and elbow (el) (DAVIS et al. 1997; LINDSLEY and GRELL 1968). Alu is a dominant mutation showing reduced and fused alula. el, on the other hand, is a recessive mutation but some hypomorphic alleles survive to adulthood as homozygotes and display an absence or reduction of the alula. Remarkably, when we analyzed the wing imaginal discs from larvae of these two genetic backgrounds, we also observed derepression of tsh in the same region of the disc as in jing– clones (Fig. 6B, C). Thus, the jing– clones and the Alu and el discs all strongly derepress tsh in the presumptive alula region. All three genotypes have small or reduced alulae, suggesting that repression of tsh in this region of the wing disc is essential to allow the development of the alula. One additional phenotype we observed in jing– clones was a weak derepression of Wg close to the DV boundary (Fig. 6E). This ectopic expression, albeit weak, might be sufficient to induce the formation of the extra wing margin bristles observed in some jing– clones comprising the wing margin. jing interacts with Polycomb Group genes and represses transcription. The observed jing phenotypes described above are likely mediated, at least in part, by the derepression of hth or tsh. In addition, we observed weak derepression of hth in the wing pouch (Fig. 6D) and in the distal antennal domain (not shown) and of wg close to the DV boundary (Fig. 6E). These data suggest Jing could be acting as a transcriptional repressor. Additionally, a mammalian gene with high homology to 16 Culi et al. Jing in the Zn-finger region, called AEBP2, encodes a transcriptional repressor (HE et al. 1999). More recently, AEBP2 was co-purified in a complex with the Polycomb (Pc) ESC-E(Z) proteins (CAO et al. 2002; CAO and ZHANG 2004). Although this is an intriguing finding, there is no in vivo evidence to suggest that AEBP2 or Jing has Pclike activity. Thus, we decided to test if jing genetically interacts with members of the Polycomb group (PcG). PcXT109 is a null allele that displays several haplo-insufficient phenotypes, including the partial transformation of second and third legs into first legs, a transformation of abdominal segment A4 towards a more posterior fate, and a partial transformation of the wings towards halteres. In PcXT109/+ heterozygotes, Ubx is also weakly derepressed in the posterior compartment of wing imaginal disc (Fig. 6F). This Ubx ectopic expression is expanded in the double heterozygote jing22F3/+; PcXT109/+ (Fig. 6G). Accordingly, adult wings from these double heterozygotes display a much stronger transformation towards haltere than single Pc/+ mutants. Curiously, this transformation is mainly restricted to the posterior compartment, which forces the wing to bend. In the most extreme cases, the wing is deeply folded and the posterior wing margin has broad nicks. We used this phenotype to quantify the degree of interaction between Pc and jing. 14% (n=142) of females PcXT109/+ have curved wings, whereas 64% (n=108) of the double heterozygous females show this phenotype, indicating a strong genetic interaction between jing and Pc. We also tested if jing interacts with other members of the PcG. Flies with one copy of a recombinant chromosome containing mutations in the three PcG genes Posterior sex combs (Psc), Additional sex combs (Asx) and Polycomblike (Pcl) have almost normal wings (Fig. 6H). However, reducing the jing dosage disturbs the normal arrangement of bristles in the posterior wing margin and increases the appearance of cuticle patches that have a high density of small trichomes, indicative of a partial wing to haltere transformation 17 Culi et al. (Fig. 6I). Together, these data demonstrate that jing genetically interacts with several members of the PcG. Surprisingly, the transformation of abdominal A4 to A5 and the extra sex combs phenotypes observed in PcG mutations were not significantly enhanced by a reduction in jing dosage, suggesting that jing may be involved in only a subset of Pc-dependent functions (see Discussion). The mammalian jing homolog AEBP2, was also shown to be a transcriptional repressor in cell culture assays, consistent with its suggested role as a member of the PcG (HE et al. 1999). The transcriptional repressor domain was mapped to the central Zn finger. To further explore the possibility that Jing contains a repressor domain, we analyzed the activities of four different Jing constructs in vivo: 1) full length wild type Jing (Jing-FL), 2) Jing’s Zn finger domain fused to the repressor domain of Engrailed (ZnfEnR), 3) Jing’s Zn finger domain fused to the activation domain E1A (ZnfE1A) and, 4) the Zn finger domain alone (Znf). Overexpression of Jing-Znf in the wing posterior compartment, using the En-Gal4 driver, has a moderate ability to induce extra-veins (Fig. 7). Overexpression of Jing-ZnfEnR or Jing-FL has a much stronger effect at generating an extra-vein phenotype. In contrast, overexpression of Jing-ZnfE1A with the same driver did not result in any noticeable phenotype. We confirmed that the three transgenes were expressed at equivalent levels by immunostaining using a myc tag present in these transgenes (Fig. 7). Thus, the addition of the repressor domain of Engrailed to the Zn-finger domain of Jing stimulates its ability to generate extra veins and, on the contrary, addition of the activation domain of E1A suppresses this potential. These results indicate that Jing acts as transcriptional repressor, at least during vein differentiation. Because one of the most striking phenotypes induced by the absence of jing is the loss of the alula, we determined whether Jing also works as a repressor during this 18 Culi et al. process. For this experiment we used the MARCM system to eliminate endogenous Jing in clones and at the same time re-supply the mutant cells with the activation- and repression- domain Jing fusions. As described above, jing– clones in the alula region trigger the derepression of tsh. This phenotype can be suppressed by the simultaneous expression of Jing-Znf or Jing-ZnfEn, but not Jing-ZnfE1A (Fig. 8). This experiment suggests that, as with vein differentiation, Jing is a transcriptional repressor during alula specification. DISCUSSION In this work, we describe the use of a modifier screen in Drosophila to identify jing as a gene that genetically interacts with hth. Our subsequent analysis of jing demonstrates that it plays a key role in forming the PD axis of the leg, a process that also depends on hth function. Moreover, we find that jing behaves as a repressor of hth expression during leg development. Thus, although the genetic modifier assay (pigmentation of the adult male A4 tergite) is distinct from PD axis specification, the screen nevertheless successfully identified a new player in leg development. In addition to PD axis specification, we also found that Jing plays several other roles during adult development. It is required for the formation of the alula and for the correct specification of the tergites and for the differentiation of wing veins. Our analysis strongly suggests that jing is used, in at least some contexts, as a transcriptional repressor. In the leg, the major target of repression is the gene hth, which is normally restricted to the proximal-most domain of the leg disc. tsh is also derepressed in jing– clones in the leg, but only within a small region of the disc, just distal to the endogenous tsh domain. In the wing, we also observed the derepression of tsh in jing– clones and more weakly, of hth and wg. That Jing behaves as a 19 Culi et al. transcriptional repressor is also supported by our finding that Jing-ZnfEnR, but not Jing-ZnfE1A, can rescue the tsh derepression observed in jing– clones in the wing. Our data also indicates that Jing requires the contribution of other factors to repress transcription, since jing is ubiquitously expressed in imaginal discs and its overexpression does not repress hth transcription in the proximal domain (not shown). One such factor could be Dac, a transcription factor expressed in an intermediate domain along the leg PD axis and that is also required to repress hth. Our findings are also consistent with the characterization of a mammalian homolog of Jing, called AEBP2 (CAO et al. 2002; CAO and ZHANG 2004; HE et al. 1999). The initial characterization of this gene demonstrated its potential to act as a transcriptional repressor and mapped the repression domain to one of the Zn fingers (HE et al. 1999). More recently, AEBP2 has been found to be part of a Pc complex that contains a histone methyltransferase activity (CAO et al. 2002; CAO and ZHANG 2004). These findings suggested that Jing may also be part of a Pc complex in Drosophila, a possibility that is supported by our results. In particular, we show here that jing genetically interacts with several members of the PcG. Accordingly, we speculate that the derepression of hth and tsh that we observe in the absence of jing function may in part be due to the requirement of jing to maintain the repression of these genes in a Pc-dependent manner. Consistent with this view are recent findings demonstrating that, in the wing pouch, tsh repression is maintained by Pc-mediated silencing (ZIRIN and MANN 2004). In contrast to the situation in the wing, it appears that in the leg, hth (but not tsh) is repressed by Pc-mediated silencing (J. Zirin and R.S.M., unpublished observations), a finding that is also consistent with the data presented here. 20 Culi et al. If Jing is a component of a Pc complex, one of the surprising findings described here is the degree of spatial specificity for the requirement for jing function. In the leg, we only observed phenotypes in jing– clones located in the medial domain along the PD axis. In these clones, hth was derepressed. However, in more distal clones, no affect on hth expression was observed. Similarly, in the wing, the predominant affect of jing– clones is on the development of the alula and the corresponding derepression of tsh in the presumptive alula region of the wing imaginal disc. No tsh derepression was observed in the remainder of the wing pouch. This observation is in contrast to the affect of Pc– clones, which show derepression of tsh throughout the wing pouch (ZIRIN and MANN 2004). The underlying reason for the localized requirement is not clear, especially if jing is broadly expressed throughout leg and wing discs. One intriguing possibility is that distinct Pc complexes may be required to maintain gene silencing in different cell types. Accordingly, jing may encode a more specialized component of some Pc complexes that help them achieve these cell-type specific repressor functions. The fact that Jing has a putative DNA binding domain, and thus could help target a subset of Pc-containing complexes, is also consistent with this proposal. We envision a possible molecular mechanism in which Jing binds the regulatory region of its target genes and, together with other transcription factors that provide regional specificity (e.g. Dac), represses their transcription. Subsequently, Jing may help recruit members of the Pc group complex and thus stabilize a repressed chromatin state. We also find it intriguing that jing has been found in several different modifier screens (SEDAGHAT et al. 2002); J.M., unpublished). Based on these observations, it appears as though jing-mediated repression may play a role in many cellular processes during development. In the future, it will be most interesting to biochemically confirm the interaction between 21 Culi et al. Jing and Pc complexes in Drosophila and to better understand the basis of the functional specificity described here. ACKNOWLEDGMENTS We thank D Montell, A Busturia, A García-Bellido, the Bloomington Stock Center and the Developmental Studies Hybridoma Bank for flies and materials. Mathew Wosnitzer for carrying out the modifier screen with hthk1. D Ferrés-Marcó and MJ García-García for their initial work on jing. E Caminero for her excellent technical help; and to members of the lab for suggestions throughout this project. This work was funded by (-) grants to R.S.M. from NIH, a grant from the Human Frontiers Science Program (RG0042/98B) to J.M. and an institutional grant from Fundación Ramón Areces to Centro de Biología Molecular SO. JC is supported by the "Ramón y Cajal" Spanish program. LITERATURE CITED ABU-SHAAR, M., and R. S. MANN, 1998 Generation of multiple antagonistic domains along the proximodistal axis during Drosophila leg development. Development 125: 3821-3830. ALDAZ, S., G. MORATA and N. AZPIAZU, 2005 Patterning function of homothorax/extradenticle in the thorax of Drosophila. Development 132: 439446. AZPIAZU, N., and G. MORATA, 2000 Function and regulation of homothorax in the wing imaginal disc of Drosophila. Development 127: 2685-2693. BESSA, J., and F. CASARES, 2005 Restricted teashirt expression confers eye-specific responsiveness to Dpp and Wg signals during eye specification in Drosophila. Development 132: 5011-5020. BISHOP, S. A., T. KLEIN, A. M. ARIAS and J. P. COUSO, 1999 Composite signalling from Serrate and Delta establishes leg segments in Drosophila through Notch. Development 126: 2993-3003. BOLINGER, R. A., and G. BOEKHOFF-FALK, 2005 Distal-less functions in subdividing the Drosophila thoracic limb primordium. Dev Dyn 232: 801-816. BRAND, A. H., and N. PERRIMON, 1993 Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401-415. 22 Culi et al. CAMPBELL, G., 2002 Distalization of the Drosophila leg by graded EGF-receptor activity. Nature 418: 781-785. CAMPBELL, G., and A. TOMLINSON, 1998 The roles of the homeobox genes aristaless and Distal-less in patterning the legs and wings of Drosophila. Development 125: 4483-4493. CAO, R., L. WANG, H. WANG, L. XIA, H. ERDJUMENT-BROMAGE et al., 2002 Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science 298: 1039-1043. CAO, R., and Y. ZHANG, 2004 SUZ12 is required for both the histone methyltransferase activity and the silencing function of the EED-EZH2 complex. Mol Cell 15: 57-67. CASARES, F., and R. S. MANN, 1998 Control of antennal versus leg development in Drosophila. Nature 392: 723-726. CASARES, F., and R. S. MANN, 2000 A dual role for homothorax in inhibiting wing blade development and specifying proximal wing identities in Drosophila. Development 127: 1499-1508. COHEN, B., A. A. SIMCOX and S. M. COHEN, 1993 Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development 117: 597-608. COHEN, B., E. A. WIMMER and S. M. COHEN, 1991 Early development of leg and wing primordia in the Drosophila embryo. Mech Dev 33: 229-240. COHEN, S. M., 1990 Specification of limb development in the Drosophila embryo by positional cues from segmentation genes. Nature 343: 173-177. COHEN, S. M., G. BRONNER, F. KUTTNER, G. JURGENS and H. JACKLE, 1989 Distalless encodes a homoeodomain protein required for limb development in Drosophila. Nature 338: 432-434. COUSO, J. P., and S. A. BISHOP, 1998 Proximo-distal development in the legs of Drosophila. Int J Dev Biol 42: 345-352. DAVIS, T., M. ASHBURNER, G. JOHNSON, D. GUBB and J. ROOTE, 1997 Genetic and phenotypic analysis of the genes of the elbow-no-ocelli region of chromosome 2L of Drosophila melanogaster. Hereditas 126: 67-75. DONG, P. D., J. CHU and G. PANGANIBAN, 2000 Coexpression of the homeobox genes Distal-less and homothorax determines Drosophila antennal identity. Development 127: 209-216. DUNCAN, D. M., E. A. BURGESS and I. DUNCAN, 1998 Control of distal antennal identity and tarsal development in Drosophila by spineless-aristapedia, a homolog of the mammalian dioxin receptor. Genes Dev 12: 1290-1303. GALINDO, M. I., S. A. BISHOP and J. P. COUSO, 2005 Dynamic EGFR-Ras signalling in Drosophila leg development. Dev Dyn 233: 1496-1508. GALINDO, M. I., S. A. BISHOP, S. GREIG and J. P. COUSO, 2002 Leg patterning driven by proximal-distal interactions and EGFR signaling. Science 297: 256-259. GEBELEIN, B., J. CULI, H. D. RYOO, W. ZHANG and R. S. MANN, 2002 Specificity of Distalless repression and limb primordia development by abdominal Hox proteins. Dev Cell 3: 487-498. GLAVIC, A., J. L. GOMEZ-SKARMETA and R. MAYOR, 2002 The homeoprotein Xiro1 is required for midbrain-hindbrain boundary formation. Development 129: 1609-1621. GOMEZ-SKARMETA, J. L., R. DIEZ DEL CORRAL, E. DE LA CALLE-MUSTIENES, D. FERRE-M ARCO and J. MODOLELL, 1996 Araucan and caupolican, two members of the novel iroquois complex, encode homeoproteins that control proneural and vein-forming genes. Cell 85: 95-105. 23 Culi et al. GOTO, S., and S. HAYASHI, 1997 Specification of the embryonic limb primordium by graded activity of Decapentaplegic. Development 124: 125-132. HE, G. P., S. KIM and H. S. RO, 1999 Cloning and characterization of a novel zinc finger transcriptional repressor. A direct role of the zinc finger motif in repression. J Biol Chem 274: 14678-14684. JIANG, J., and G. STRUHL, 1996 Complementary and mutually exclusive activities of decapentaplegic and wingless organize axial patterning during Drosophila leg development. Cell 86: 401-409. KEHL, B. T., K. O. CHO and K. W. CHOI, 1998 mirror, a Drosophila homeobox gene in the Iroquois complex, is required for sensory organ and alula formation. Development 125: 1217-1227. LECUIT, T., and S. M. COHEN, 1997 Proximal-distal axis formation in the Drosophila leg. Nature 388: 139-145. LEE, T., and L. LUO, 2001 Mosaic analysis with a repressible cell marker (MARCM) for Drosophila neural development. Trends Neurosci 24: 251-254. LINDSLEY, D. L., and E. H. GRELL, 1968 Genetic variations of Drosophila melanogaster. J Publs Carnegie Instn. LIU, Y., and D. J. MONTELL, 2001 Jing: a downstream target of slbo required for developmental control of border cell migration. Development 128: 321-330. MAHAFFEY, J. W., D. F. JONES, J. A. HICKEL and C. M. GRISWOLD, 1993 Identification and characterization of a gene activated by the deformed homeoprotein. Development 118: 203-214. MANN, R. S., and M. AFFOLTER, 1998 Hox proteins meet more partners. Curr. Op. Genet. Dev 8: 423-429. MARDON, G., N. M. SOLOMON and G. M. RUBIN, 1994 dachshund encodes a nuclear protein required for normal eye and leg development in Drosophila. Development 120: 3473-3486. MARTINI, S. R., G. ROMAN, S. MEUSER, G. MARDON and R. L. DAVIS, 2000 The retinal determination gene, dachshund, is required for mushroom body cell differentiation. Development 127: 2663-2672. MORATA, G., 2001 How Drosophila appendages develop. Nat Rev Mol Cell Biol 2: 89-97. NEUMANN, C. J., and S. M. COHEN, 1996 Sternopleural is a regulatory mutation of wingless with both dominant and recessive effects on larval development of Drosophila melanogaster. Genetics 142: 1147-1155. PANGANIBAN, G., 2000 Distal-less function during Drosophila appendage and sense organ development. Dev Dyn 218: 554-562. PENTON, A., and F. M. HOFFMANN, 1996 Decapentaplegic restricts the domain of wingless during Drosophila limb patterning. Nature 382: 162-164. PICHAUD, F., and F. CASARES, 2000 homothorax and iroquois-C genes are required for the establishment of territories within the developing eye disc. Mech Dev 96: 15-25. RAUSKOLB, C., 2001 The establishment of segmentation in the Drosophila leg. Development 128: 4511-4521. RAUSKOLB, C., and K. D. IRVINE, 1999 Notch-mediated segmentation and growth control of the Drosophila leg. Dev Biol 210: 339-350. RIECKHOF, G., F. CASARES, H. D. RYOO, M. ABU-SHAAR and R. S. MANN, 1997 Nuclear translocation of Extradenticle requires homothorax, which encodes an Extradenticle-related homeodomain protein. Cell 91: 171-183. 24 Culi et al. SEDAGHAT, Y., W. F. MIRANDA and M. J. SONNENFELD, 2002 The jing Zn-finger transcription factor is a mediator of cellular differentiation in the Drosophila CNS midline and trachea. Development 129: 2591-2606. SEDAGHAT, Y., and M. SONNENFELD, 2002 The jing gene is required for embryonic brain development in Drosophila [corrected]. Dev Genes Evol 212: 277-287. SHEN, W., and G. MARDON, 1997 Ectopic eye development in Drosophila induced by directed dachshund expression. Development 124: 45-52. SONNENFELD, M. J., N. BARAZESH, Y. SEDAGHAT and C. FAN, 2004 The jing and ras1 pathways are functionally related during CNS midline and tracheal development. Mech Dev 121: 1531-1547. THEISEN, H., T. E. HAERRY, M. B. O'CONNOR and J. L. MARSH, 1996 Developmental territories created by mutual antagonism between Wingless and Decapentaplegic. Development 122: 3939-3948. VACHON, G., B. COHEN, C. PFEIFLE, M. E. MCGUFFIN, J. BOTAS et al., 1992 Homeotic genes of the Bithorax complex repress limb development in the abdomen of the Drosophila embryo through the target gene Distal-less. Cell 71: 437-450. WEIHE, U., R. DORFMAN, M. F. WERNET, S. M. COHEN and M. MILAN, 2004 Proximodistal subdivision of Drosophila legs and wings: the elbow-no ocelli gene complex. Development 131: 767-774. WU, J., and S. M. COHEN, 2000 Proximal distal axis formation in the Drosophila leg: distinct functions of teashirt and homothorax in the proximal leg. Mech Dev 94: 47-56. WU, J., and S. M. COHEN, 2002 Repression of Teashirt marks the initiation of wing development. Development 129: 2411-2418. XU, T., and G. M. RUBIN, 1993 Analysis of genetic mosaics in developing and adult Drosophila tissues. Development 117: 1223-1237. ZIRIN, J. D., and R. S. MANN, 2004 Differing strategies for the establishment and maintenance of teashirt and homothorax repression in the Drosophila wing. Development 131: 5683-5693. FIGURE LEGENDS Figure 1. hth genetically interacts with jing. (A, E) Dorsal view of the male abdomen in the indicated genotypes. The abdominal segment A4 (asterisk in A) has a thin stripe of darkly pigmented cuticle in wild type and jing22F3/+ flies (A and D). Heterozygous hthK1 flies have a darker A4 (B). This pigmentation phenotype is exacerbated by reducing jing genetic dosage (C and E, arrows). The abdomens shown in D and E 25 Culi et al. were dissected and mounted in Hoyer's/lactic. The abdomens shown in A, B and C were photographed without previous dissection. Figure 2. Phenotypes of jing– clones in the legs. (A) A wild type leg is composed of nine segments. From proximal to distal: coxa (Co), trochanter (Tr), femur (Fe), tibia (Ti) and tarsal segments from 1st to 5th . The bristles located in the coxa, trochanter and proximal femur do not have bracts, whereas the bristles located in distal segments do have bracts, as shown in the close-up views. The region of the leg that requires Jing activity is shadowed in green. (B) Two jing22F3 clones located in the femur segment are outlined in blue. The mutant territory can be recognized by the f bristles. Most of the bristles in the distal clone lost the bracts, suggesting a transformation to a more proximal fate. The inset shows an internal vesicle containing a f bristle and located in the femur. (C) The bristles of a clone located in the tibia, outlined in blue, also lost the bracts, as shown in the inset. (D and E) A clone originated in the tibia induced the fusion of this segment with the femur (D). Similarly, a clone in the 1st tarsal segment induced a fusion between this segment and the tibia (E). The f bristles present in these two clones do not have bracts. (F) A 1st tarsal segment displays an overgrowth, which is probably caused by the presence of a jing clone. In this region of the leg, it is difficult to identify the clones because of limitations of the f marker. Figure 3. jing phenotypes in wing vein differentiation. (A) A ventral jing22F3 clone marked by the f trichomes is outlined in blue. Inside the clone, the longitudinal vein 2 (L-2), a ventral vein, has lost most of its characteristics, like corrugation and pigmentation. However, it preserves the vein-specific trichome orientation. Part of L2 that is not included inside the clone but abuts it, is non-autonomously affected. (B) 26 Culi et al. A dorsal jing22F3clone is outlined in blue. Part of L-4 abutting the jing clone is nonautonomously eliminated. The proximal segment of L-4 is ventral and thus, not affected by the dorsal clone. (C) Wing showing ventral (red) and dorsal (blue) jing22F3 clones. An elongated ventral clone strongly decreases the pigmentation and corrugation of L-2 (asterisk). Another ventral clone decreases the pigmentation of L-4 but does not completely eliminate the vein. In the same clone, an ectopic vein appears along the clone border (arrow). The distal L-5 is completely eliminated by two clones which affect the ventral and dorsal wing surfaces. Figure 4. jing phenotypes in the alula and hinge. (A) Close-up view of the hinge and alula region of a wild type fly. (B) A jing22F3 clone induced the fusion of the alula with the main blade. The clone segregated from the cuticle and formed an internal vesicle (arrow and inset). (C) A small clone induced a finger-like outgrowth of the alula cuticle. The mutant cells are identified by the f trichomes and are outlined in blue. We also observed formation of darkly pigmented vesicles (arrow in D) and finger-like outgrowths (arrow in E) in the hinge and proximal wing region. Figure 5. hth and tsh are de-repressed in jing - clones. (A and B) In situ hybridization to detect jing transcripts shows general expression in both leg (A) and wing (B) imaginal discs. (C and D) jing 22F3 clones in leg imaginal discs are marked by the absence of GFP (green). In some clones Hth (red) is ectopically expressed. hth derepression is only observed in the Dac (blue in D) expression domain. Note that not all the cells of a clone show the same level of hth derepression (asterisks). (E) In jing 22F3 clones, marked by the absence of GFP (green), Tsh (red) is ectopically expressed 27 Culi et al. but only in a region a few cell diameters distal from the Tsh endogenous expression domain (arrow). (F) Leg disc containing jing 22F3 clones marked by the absence of arm-lacZ (green) and that also express UAS-Jing-FL, using MARCM. In these clones, hth is not derepressed. Figure 6. (A) Wing imaginal disc where jing22F3 clones were induced and marked by the absence of GFP (green). Tsh (red) is ectopically expressed (arrows) wherever the clones are located in the presumptive alula region. The intersection between the DV stripe and the inner ring of Wg (blue) expression marks the presumptive alula region. (B and C) Wing imaginal discs of the Alu56c (B) and elA1 (C) genotypes. Tsh (red) is ectopically expressed (arrows) in the alula region. Wg (blue) is shown in C as a reference. (D and E) In the wing imaginal discs, hth (purple in D) and Wg (purple in E) are weakly derepressed (arrows) in some jing22F3 clones, marked by the absence of GFP (green). (F and G) Ubx (blue) is weakly derepressed in the posterior wing pouch of a larvae with the genotype PcXT109/+ (arrow in F). This derepression is enhanced in the genotype jing22F3/+; PcXT109/+ (arrow in G) and is not observed in jing/+ larvae. The asterisk marks a third leg imaginal disc, which has a high level of Ubx protein. The degree of Ubx derepression varies from disc to disc in both genotypes; shown are representative examples. Notably, derepression of Ubx in the wing disc is in the same region that shows derepression of tsh in jing– clones. (H and I) Close up views of the posterior wing margin near the distal vein L5 from a Df(2)vgD, AsxXf23, PclXM3/+ female fly (H) and a Df(2)vgD, AsxXf23, PclXM3/jing22F3 female fly (I). The bristles from the posterior wing margin are more disorganized when jing dosage is reduced (I). 28 Culi et al. Figure 7. Jing activity can be phenocopied by a Jing-En-repressor fusion. UAS-Jing full length (FL) and three different transgenes containing the Zn finger regions of Jing alone or fused to the Engrailed repressor domain (EnR) or the E1A activation domain, were expressed in the posterior compartment with the driver Engrailed-Gal4, as indicated. Overexpression of UAS-Jing-FL and UAS-Jing ZnfEnR induce the differentiation of ectopic vein material (arrows). UAS-Jing Znf has a much reduced capacity to induced extra-veins. On the contrary, overexpression of UAS-Jing ZnfE1A does not have any phenotype. The panels at the right show that the three transgenes containing the Zn finger domain were expressed at similar levels, as revealed by antimyc immunostaining. Figure 8. Jing represses transcription. Wing imaginal discs containing clones of cells mutant for jing22F3 and expressing UAS-Jing-FL, UAS-Jing ZnfE1A or UAS-Jing ZnfEnR as indicated, and detected with the anti-Jing or anti-Myc antibodies (green). UAS- Jing-FL can completely rescue jing loss of function. Thus, Tsh (red) is not ectopically expressed in jing clones located in the alula region (arrowhead). Expression of UAS-Jing ZnfEnR almost completely reverts tsh derepression (arrow; most discs from this experiment do not show detectable tsh derepression). On the contrary, UAS-jing ZnfE1A is unable to rescue jing loss of function, allowing a strong ectopic expression of Tsh (arrow). Supplementary Figure 1. jing genomic region. The four jing transcripts and the insertion sites for the transposable P elements P1094, P07112 and Prh623 are shown 29 Culi et al. in the map. The coding region is shown in black in the different transcripts. The first codon in exon 4 is an stop codon. 30