Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Signal transduction wikipedia , lookup
Tissue engineering wikipedia , lookup
Endomembrane system wikipedia , lookup
Cell growth wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cell culture wikipedia , lookup
Cytoplasmic streaming wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cellular differentiation wikipedia , lookup
List of types of proteins wikipedia , lookup
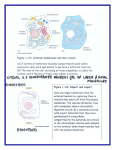
Acta Bot. Neerl. March 39(1), 1990, 1-18 p. Review plant cytoskeleton: its significance in plant The development J. Derksen F.H.A. Wilms and E.S. Pierson ED Department of Experimental Botany, University ofNijmegen, Toernooiveld,NL-6525 Nijmegen, The Netherlands CONTENTS Introduction 1 Structure and General composition organization of the of the The cytoskeleton in cell The cytoskeleton in 1 cytoskeleton 4 cytoskeleton and differentiation morphogenesis 7 12 plant morphogenesis remarks Concluding 12 cell differentiation. Key-words: cytoskeleton, morphogenesis, INTRODUCTION The of cytoplasm It cytoskeleton. eukaryotic connects cells contains the various with each other and with the in cesses the cell, cell and other cytoplasmic elements of the cell membrane and is involved in many plasma including three-dimensionalnetwork of filaments: the a organelles division, morphogenesis, dynamic redistribution of pro- surface components, endo- and exocytosis and the positioning of cytoplasmic elements (reviews: Dustin 1984; Lackie 1986; Bershadsky & Vasiliev 1988; plants: Traas 1989). The three major constituents of this system mediate filaments. They can be are microtubules, microfilaments based discriminated, on their diameter and and inter- by of means immunocytochemistry. The informationconcerning the animal cells. However, to do justice to all studies been discussed (1982). see For Fosket As plant as concerning extensively by a recent review on plant cells plant cytoskeleton. & Hardham and biochemistry seems increasing, Literature (1982), genetics scanty when compared it will be not prior Hepler (1985) of plant possible to to 1980 has and Lloyd cytoskeletal proteins (1989). cells are and cell expansion immobile, a particular plant shape by polar growth, within a tissue must and structure following this i.e. the Gunning the cell divisionand cell expansion, i.e. first in cytoskeleton interest has been rapidly composition we will discuss embryonic, plants with STRUCTURE AND occur of the recent special in a or on reference only be obtained of by directional cell death. Thus, cell division co-ordinated way. Here cytoskeleton results can specific eukaryotic we shall describe cells in general and the organization of the cytoskeleton in higher, to COMPOSITION morphogenesis OF THE and cell differentiation. CYTOSKELETON Microtubules Microtubules are average external tubular structures with diameter of about 25 internal diameter of about an nm. They consist of 1 two 15nm and evolutionally an related 2 J. a-tubulin and proteins, each p-tubulin, 50 000 kD. The microtubularwall is numbers that Protofilamentsconsist of occur. and asymmetrical are microtubule all respect Microtubules assumes — and 1978) steady a side. Dynamic microtubules proposed with with together mitotic dynamics have been described Borisy 1988). Tubulins of et al. (Cleveland 1980; continuously 1978), but explain some may the relative herbicides associate Olmsted with in vivo, as a as see well relate & also may play an with 1 In nm dimers in a in 10° a of of to the slowly, of rapidly eventually al. 1984). However, be much both at in ends, behave to the the growing according dynamic instability (e.g. to cells of interphase stable. In such more at disassembling their cases Sammak & disappearance occurrence 1986a). different evolutionary flagella (Cavalier- of a cell wall. This difference A of variety may also largely has been found proteins microtubule-associated proteins (MAPs) determine microtubule elements and organelles. enzymes in which Ca 2+ et stability /calmodulin al. 1985, to to (review: and Their activity appears & Vallee 1982; Larsson (Theurkauf a of cilia and microtubules to colchicine and their sensitivity MAPs below). -(-side and disassembly population & The first model 1984). appear to be less conservative than in animal cells plant the treadmilling (Margolis which has been attributed to Mole-Bajer role important —side. assemble and disassemble. Two microtubules appear tempered cascade of phosphorylating a polar proto- the shifted about polarity the at to association with other cytoskeletal regulated by side are & Kirschner mode of 1989), insensitivity (Bajer in resulting processes: appeared higher plants Fosket et (Salmon microtubules: 1986, these assembly pressure. This differencemay result from the Smith but different tubulin dimers long, a-P P-tubulin distinct a the existence of a population a spindles of the microtubules animals, part show explain to continuous a model dynamic instability nm orientation and same dynamic instability (Mitchison state the approximately protofilaments, direction, one -|-side, of weight ET A L. 1984). instability predicts microtubules. In the oriented in called the static elements but are no mechanisms have been molecular a made up of 13 chain of about 8 a Microtubules thus left-handed helix (Dustin Wilson is protofilaments have the each other. to all are filaments. The a-tubulin side with generally DERKSEN to its be and cAMP Schulman et al. 1985). Microtubules have known role in been related movement of various types of to metaphase involved in the transport of vesicles and (Dustin the and dynamics of the of are reticulum endoplasmic their well cilia/flagella, they neurons 1984; Bershadsky & Vasiliev 1988). Microtubules organization Hotani like in organelles, besides movements: chromosomes and can be and other animal cells also involved in probably (Chen & Lee 1988; Vale & 1988). Microfilaments The backbone protein of about 42 kD. of microfilaments is formed In the filaments actin molecules double-stranded twisted rope. The Individual microfilaments have Microfilaments considered are also to occur. dynamic Bershadsky cells & Vasiliev (review; Staiger stress a diameter of 5-7 and both fibres, 1988). They & Schliwa are actin which has piled major repeat distance is Microfilaments often ation like in muscles and by 1986). microscopic preparations represent treadmilling occur where can nm in they 38 in such a a show a way stable The microfilaments that actin filaments, which have a can be a polarity. have been regular organiz- as recently weight form 13 molecules. distinct (reviews: also form loose bundles, such to as dynamic instability bundles with very molecular involving nm, and and large are up Lackie in many seen 1986, plant in electron been confirmed CYTOSKELETON for SIGNIFICANCE IN cells with plant filaments the of use also form more can immunological probes less dense or The different types of association various associated 2+ Ca /Ca 2+ Actin is of actin (Lancelle filaments and play cells (Condeelis 1987). Plasma-streaming depends and vesicle-mediated secretion often appear in Jackson 1982; reticulum has been endoplasmic & Schliwa 1987; Staiger stability depend been identified thought to be actin proteins 1986; sensitive are to role. be actin to the on (Lackie et actin filaments. Also 1987). Actin 1989). 1974; Parthasarathy on movements Kristen their now important regulatory an Hepler & associated with membranes. As in microtubules many associated which thus regular component of plant a & Schliwa Staiger 1988). -calmodulin, 3 networks, often from which numbers have proteins & Vasiliev Bershadsky DEVELOPMENT Also the al. 1985; organelle dependent (reviews of the organization dependent (Quader et al. 1987). Intermediatefilaments Unlike microtubules and microfilaments, intermediate filaments geneous, yet evolutionally related, proteins (reviews in Nagle In animals they the eukaryotic tissue are cell in specific, indicating al. 1982). intermediate proteins, filaments. al. et been described isolated a spectrin-like by Goodbody et al. but do not by and The protoplasts (Powell carrot Goodbody et with al. to nm. Intermediate These bundles contain microtubules plant (Dawson from plant part of the proteins detected al. et & Yan 1985; (1988) membranes. at the a with animal intermediate filaments (1989). Recently, Wang ankyrin-like protein an nm. immunological homology distributions of extraction-resistant against filamental organization a P-proteins also be diameter of 50-100 co-localize intermediate filamental proteins, must Roop 1988). this system in have have the Maybe membrane plasma residues cell of fern spermatozoids also co-localize with microtubules in immunofluorescent preparations, 1988) show to hetero- (1989). Antibodies raised al. a Distinct is identical spectrin-like protein et first described in were of which show some They 1989). recently (Marc cells plant bundles with large number of different family of less conservative character The diameter of individual intermediate filaments is 7-10 filaments form Hargreaves 1988; a Steiner & general. Intermediate filaments of et a are in phloem (Marc perhaps or cells form tubular regarded, by definition, as a & Gunning 1988). They might detect MAPs. structures (Cronshaw et al. intermediate filament plant-specific 1973), that system. Cytoskeleton-associated proteins The various associated their to organization be present in animal systems plants (Lackie few have been identified yet. possible 1n of microtubules and microfilaments proteins and function. Numerous proteins are known 1986; Bershadsky & Vasiliev 1988), but in higher They are summarized in Table I with their together functions. higher plants, the existence of an but the basis for such Cytoplasmic a acto-myosin concept has been set as 1987) and kinesin but expect them to be (Vale et present al. in 1985) driven system has still myosin microtubule-based motors, such Vallee we cytoskeleton-associated determine largely have as not higher plants as is the yet not been proven, clearly present. dynein-like MAP-1C been established & (Paschal firmly in plants, well. The latter may mediate the interactions between microtubules and the tubular endo- plasmic reticulum (e.g. Vale & Hotani both actins and microtubules as 1988). Directional movement may also depend on shown for the alga Bryopsis (Menzel & Schliwa 1986). 4 J. DERKSEN Table 1. ETAL. Microtubule (MT) and actin filament (AF) associated proteins that have been identifiedin the cytoplasm of cells of higher plants Protein References Group MAPs MT Cyr Kinesin MT Moscatelli et & Palevitz Calmodulin MT Wick et Possible role Microtubule bundling (1989) al. (1988) Force-generating, organelle transport. al. (1985) Ca 2+ -binding protein, regulatory functions Troponin Lim AF/MT et al. 2+ Ca (1986) -binding protein binds to tropomyosin, regulatory functions Van Ankyrin AF Wang& Myosin AF Vahey et al. (1982) Parke Yan et Grolig AF Spectrin The presence Yan (1988) Connecting spectrin-like proteins movements actin to membrane proteins indicates that the associations of those in animal cells. to cytoskeletal proteins of animal cells will be present somehow in cells. GENERAL ORGANIZATION The techniques used The organization of OF Electron microscopy. microtubules, microfilaments ditions used, during the compared Classical electron the and Northcote problems 1973; Howard & & Staehelin 1988). immunological probes Large be can surfaces at The embedded material (Wilms cell parts can Light microscopy. against (Lloyd et animal (IF) probes al. 1979; Wick et al. of use 1985). serial to the con- depolymerized contrasted poorly cryo-techniques, can be as to study of the al. use of by using cleaving polyethylene glycol (PEG) 1985) this may be useful. procedure be evolutionally spatial & 1986; Hepler 1989). obtained (Hawes sectioning, though appear et be conditional for the Also sections of mounts (Hereward 1986; Lancelle Lancelle & proteins, especially tubulin, allow enable visualiz- soluble components of the by may (e.g. analyses Cytoskeletal elements Immunofluorescent the techniques and whole using only techniques. be studied. by of these al. et 1990) also be studied antibodies raised obscured 1979; Emons & Derksen quantitative Traas become may the ultrastructural level allowing techniques (Traas 1984; areas can use number of a actin filaments, may become are or overcome Aist using intermediate filaments, but due elements medium, small cytoplasm. Moreover, only Fixation fibrous embedding CYTOSKELETON microscopical (EM) techniques and cytoskeletal elements, especially procedure to THE elements has been studied cytoskeletal ation of Craig intracellular al. (1989) Wang & most membrane proteins al. (1988) et et of ankyrin-like and We expect that to (1986) actin with the membranes may be similar plant actin Connecting Organelle movement, force generation, al. (1986) et al. Tang (1988) Larger is laborious. conservative could be used in organization plant and cells. in entire cells 1981) and in sections of PEG-embedded material (Hawes & CYTOSKELETON SIGNIFICANCE 1. Microtubules in root cortex cells of Fig. (1984). Horne the longitudinal axis though investigation techniques ning are the two cells on the as well almost are 1000. x three-dimensional (McCurdy useful for Traas el al. the microtubules right, of et phalloidin (Wulf of actin organization al. al. et filaments, 1988)). large scale-studies, but for detailed analysis a required (e.g. Segaar 1990). microscopy light microscopic techniques (CSLM; of the dynamics Quader e.g. image photography (e.g. understanding helical, in of fiuorochrome conjugates of the extremely of advanced use laser enhanced are are Magnification: antibodies have been used The IF methods The of the cell. The introduction 1985). allowed 1979) EM Lepidium. Immunofluorescence preparation according to In the left two cells the microtubules parallel to 5 IN DEVELOPMENT & in Schnepf Lichtscheidl & Weiss of the cytoskeleton cells such plant 1986) 1988) as and confocal scan- video/computer will contribute to a better and its interactions with other cellular structures. General organization Microtubules microtubules in Gunning present throughout the cytoplasm, but large arrays of parallel cortical always present, Hardham & microtubules throughout are are are the and interconnected and cell (Lloyd 1984; Traas thought 1984). Recently it to Derksen, unpublished data). If these interconnected microtubules could considerable forces on not only 1989; form could be Nicotiana about 50% of the cortical microtubules is & J. in vacuolated cells especially conspicuous 1982; Lloyd see an also almost shown were withstand strong determine the also present the at the (reviews cortical in of (H. Kengen dynein-type such but could even exert of forces, helix protoplasts a their environment. At the surface of the nucleus microtubule not The uninterrupted that organizing centres (MTOCs) initiatemicrotubule assembly after cell division(e.g. Wick & Duniec do 1). interconnected regularly interconnections Fig. organization plasma of the cortical membrane (Gunning et microtubular skeleton may self-assemble into cytoskeleton, al. 1978) and are present that 1983). They probably as nucleating in anucleate sites are cytoplasts highly organized patterns (Bajer & J. 6 Fig. 2. Actin preparation x 8000. filaments with in anucleate an rhodamine-phalloidin of subprotoplast as a pollen of tubes to Rutten probe, according DERKSEN ET AL Nicotiana. Immunofluorescence Derksen (1990). Magnification & (Photograph: T, Rutten.) Mole-Bajer 1986b). microtubules takes protoplasts In place at the of the alga Mougeotia reorganization of the cortical from the nucleus plasmalemma, independent (Galway & Hardham 1986). Like microtubules, actin filaments are present in cortical arrays nucleus is anchored in with the cells (review: basket of actin filaments and a periphery (Derksen Staiger & Schliwa 1986). et al. microfilaments may show self assembly Intermediate filaments in membrane and fluorescent be can seen preparations, seem to connect as et al. Interactions between & Derksen see in cytoplasts also Fig. 2). associated with the spindles. Arrays periphery interphase The plasma streaming co-localize with microtubules in those of mitotic 1987). the nucleus but in anucleate 1989; probably present that al. plasma- immuno- of filament bundles cells (Goodbody et al. 1989). cytoskeletal cytoskeletal elements elements form Davis 1981; Griffith & Pollard 1986) are patches also in filaments may be cross-linked have cells (Rutten et connect These bundles reflect is uncertain, origin the nucleus with the cells 1989; Hargreaves The various plant 1986b). Their (Traas bundles large been observed in both by MAPs 1982). In an or integrated system. by plant actin-associated cells they dry-cleaved preparations often et proteins (e.g. co-localize, (Traas and sections of freeze-substituted material (Tiwari Microtubules and actin et al. Bennett & and cross-links 1985; Pierson al. 1984; Lancelle et al. et al. 1987). CYTOSKELETON SIGNIFICANCE Fig. 3. Micrograph of microfilaments present or are a dry-cleaved preparation at the surface to accompany microtubules Single microfilaments dry cleaved data); see (J.J.). of a protoplast from Often these filaments (T). Magnification; were co-aligned of Nicotiana preparations also IN DEVELOPMENT x 59 000. a 7 Nicotiana cell culture. seem to be connected Numerous with coated putative pits (\J7), (Photograph: H. Kengen.) with microtubules Fig. 3). However, doubt exists about distances over protoplasts (H. Kengen to up & J. Derksen, the actual 1 -46 pm in unpublished of these filaments nature (see below). A clear co-localization cent of preparations over large parts tubes pollen of the cell has been observed in immunofluores- (Raudaskoski et al. distribution of the microtubules has been reported distribution(Derksen & Traas Intermediate filaments of (Goodbody filaments that nect J. al. et are 1989). visible in an intact actin microtubule THE skeleton, cells dry-cleaved but may be et al. least 1989) partly, and the on actin (Emons&Traas depends IN et least at partly patchy a identical way to cells and that accompany microtubules and 1986; Quadereta/. 1986; The distribution of the degradation (Goodbody CYTOSKELETON at co-localize with microtubules in present proteins coated pits with microtubules Derksen, unpublished data). depend, 1984). plant The Pierson 1987; to larger on microtubules. al. 1989). CELL bundles is MORPHOGENESIS Kengen& independent become They H. the con- from dispersed after AND DIFFERENTIATION Determination of the division plane The of position microtubular Prior to the drugs cell versa may (Burgess by bands bands in nuclear 1988). may be largely disposition nucleus will microtubules determined of the nucleus move radiating 1970; Pickett-Heaps 1974). during phragmosome pre-prophase cause division, the may be determined vice nucleus positioning The fundamental is & not regulatory Libbenga microtubules, & Lloyd as anti- 1984). towards the future division plane, which from the nucleus to the cells These microtubules may formationand coincide in time and (Venverloo by (Clayton place 1987). However, unquestioned (Clayton & periphery already with the formationof the a role of the preprophase Lloyd 1984; Mineyuki mechanisms are unknown. and be present Also, et al. microfilaments are 8 J. in the present metaphase preprophase bands (McCurdy preprophase band microtubules the and may filaments remain, determined path (Traas show distinct et al. of cell al. et al. 1987; Lloyd 1990) but process, polarity (Traas al. band but not changes Hormonal control elements. causes Pisum sativum and a al. Vigna few hours. In cortex ethylene high & Shibaoka & Shibaoka growth against treatment explants, (Wilms 1984). However, Hormones also (Fragata may act epidermal the number of 1974; effects the direction of cell epidermal cells of maize ethylene production (Imaseki organization of the development Medicago mesophyll by changing both & the cells (Meijer of internodes pea microtubules and by gibberellic depend of the pumps microfilaments (see acid the colchicine on 1984). Auxin cytoskeleton. et al. 1988). Since auxin induces cortical microtubular skeleton & Simmonds it diverse: it may protect stimulation is indirect. However, the presence of auxin a cause Gibberellic acid and the orientation of the cortical micro- 1988). Hormones may and channels 1981; Saunders 1986; review: microtubules and are can it may be concluded that the action of auxin of al. Doonan 1989). also Mita & Shibaoka see et and low temperature cremart organization expansion distribution of cation Hepler the coleoptiles (Bergfield 1985), cytoskeleton conditional for the (Saunders for discussion on 1982; cells transverse conversely, growth cells of Roberts re-orientation of micro- Physcomitrella. the effects of gibberellic acid indirectly et al. cytoskeletal epidermis & Wolters-Arts cells of microtubules in cochicine inhibitionbut concentration used tubules in unknown. benzylaminopurine (100 gM) tip-growing It increases 1987). 1981; Lang cells of tobacco may be inhibited by colchicine. The differences however, may treatment the underlying basically between hormones and the of microtubules by colchicine, prevents depolymerization (Mita with the pre- not role in cell division regulatory cell division are still concentrations of rearrangement of cortical causes (Akashi a re-orientation of cortical microtubules in of microtubules in depolymerization in the establish- The actual mechanisms plane. to and phragmoplast, radiata (Steen & Chadwick showed that (1985) and the precise relationship a tubules is accelerated after et involved are of the cytoskeletal organization Ethylene within pre- Lammeren et al. 1989; Traas (see which indicates cytoskeletal organization prior Little is known about the 1985) spindle 1985), in determination of the division in the 1989). & Wick (Gunning band actin along out involved in the co-ordination of the microtubules probably only et 1988). During preprophase apparatus ET AL Both microtubules and actin filaments probably are but the cytokinetic 1988). & Traas 1988; Lloyd disappear the meiotic divisions and Calmodulin is associated with the prophase the guide to configurations during 1989; Bednara meiotic division ment et help al. et DERKSEN Schnepf above), such on the 1986). changes in on seems to protoplasts act on cell the be of polarity plasma-membrane As Ca may 2+ also may affect affect the organization of cytoskeletal elements. Controlof cell wall It is generally deposition believed that the orientationof nascent cortical microtubules. Several models have been trol of microfibril orientation (reviews; Robinson & All models imply that microtubules connected cellulose microfibrils is controlled by proposed to to describe microtubular Quader 1982; Heath & Seagull the plasma membrane, con- 1982). i.e. the cortical microtubules, would prevent free diffusionof cellulose microfibril synthesizing complexes in the membrane, leading Herth 1985). Also the to parallelism insertion of of microtubulesand Golgi-vesicles nascent microfibrils (see also: with non-cellulosic wall material has CYTOSKELETON been thought SIGNIFICANCE IN DEVELOPMENT be under microtubular control to 9 Roo (Goosen-de 1973, also see Herth 1985). As discussed in occur a clear relationship been In thought to Traas al. mainly cases, al. et 1984; Emons 1989; effected may be marine algae and thus A & Hoffmann simultaneous deposited will and Often such a similar In 1989). a other or lead as drugs Valonia ventricosa affect the in factors. The also from transverse to parallel the to be co-incidental. ation in so (Wilms & far, cell to cells elongating of cellulose Patterning instances some et al. 1986), electric (Roland number and (Emons 1985) 1987). density evidence for a et al. 1990; Wilms be, e.g. Ca 1988). 2+ et (Quader al. et It has also been Particular wall textures of cellulose synthesizing cytoskeleton, orientation. vector-sum nor If out may expanding a absent In been microtubular cortex cells of cell axis (Wilms colchicine did 1989). Thus, common & not such regulatory of both microtubules & Hardham be more 1982), might complicated 1990; Wilms & Kengen or al. may as or by such different a relationship be mechanisms that microfibrillarorganiz- in the exist, thought plasma based arise from (Emons 1986). nascent they to membrane(Emons be taken not to rejected cell the internal geometry of the cell between the orientation of orienting on interfere otherwise. Such have also been complexes 1990). depend 1986), turgor pressure (Derksen proposed that co-orientation itself it relationship can determining Cytoskeleton the cell’s pointed causal be within the or on expansion deposition spontaneously by self-assembly al. cells mesophyll changes Wolters-Arts in combination with cell dimension and matrix substances It should be In to fields (Preston et In the the orien- and relations with microtubular patterns may need reconsider- thought ation would occur wall 1988). cremart (Gunning differentiation. Various factors may be conditional, factors have been not may microtubule-microfibril interactions may be diverse and Obviously, The do 1988) and longitudinal elimination of microtubules by orientation with respect transverse generally thought 1990). effect, microfibrils has supply of microtubules and microfibrils cellulose microfibrils in nascent al. after deposition deposition. al. et and simultaneous changes might be solely co-incidental and depend and et same deposition after cessation of auxin in microfibril orientation change exist cytoplasmic organization (Hayano orientation of microtubules organization 1988). However, the has 1985). change time interval same the different. Also in clearly hairs, root (Emons in cell wall changes to cannot in microfibril to the entire control of microfibrilorientationhas been inferred (Bergfield Derksen Raphanus change microtubules and microfibril coleoptile segments explants, relationship a relationship in ordered patterns microfibrils is nascent between relationship tobacco seems to Also in seed hairs 1982; Emons & Wolters-Arts 1983; by colchicine, deposition, indirectly Boergesenia forbesii observed in maize 1987). hairs: Emons direct microtubular control a tation of microtubules and (Hahne al. 1986; Traas & Derksen microfibril on indicate necessarily et deposition during xylogenesis. growing tip cells, root but microfibrils were still occurred, effect of colchicine a in 1985; of the microtubules depolymerization such (Quader cells and obligatory (e.g. Lloyd 1984). tubes: Derksen et exist seems to be number of a (pollen microtubular control of microfibril by Hepler (1985) elongating cylindrical cells, guard on may as pre-requisite or microfibrils and the the act absence as of vectors, co- the the orientation of the microfibrils. and cell expansion cells a periphery. central vacuole develops, leaving only a small layer of cytoplasm at J. DERKSEN 10 In microtubules generally appear stretching cells, tion of cell expansion (Gunning be oriented to & Hardham 1981; Gunning transverse 1984; Traas Traas ential cell tip 1989). of al. Traas 1985; (1984) expansion, more or of presence meristematicand see that also transverse less as in the to in expanding pitches (Lloyd the 1983, root of cells cortex of both axial and circumfer- vector-sum stretching randomly organized cells. The orientation of microtubules in microtubules at et (Traas al. 1985; Emons surfaces, of non-expanding cells in these roots, may support this cortex the direc- Fig. 1). microtubules cells would be determined in a similar way tip-growing The 1989; supposed would be oriented Raphanus the et al. et AL In many cells, 1982). microtubular skeleton shows helix-like configurations with different to ET assumption (Derksen al. et 1986a). It remains open whether cell indirectly, by This might whether these expansion either as al. a Such observed in and in Avena a the exact changes out by pressure. active root cells force from (Traas transverse to et al. & Oshima 1984; Hogetsu & longitudinal or Roberts of helical patterns occurrence epicotyls (Iwata oblique more or expansion (Lloyd 1984; cell during and Pisum mesocotyls, would behave configurations relate the microtubule orientation demands to more a a i.e. cytoplasm, cell to 1986) Hogetsu 1988). occur expansion role at the the one, In after mainly but fail point to behaviour of microtubules, whereas the yet needs considerablereorganization cell axis immediately of during stretching. orientation of microtubules in long appreciable in cell (Wilms change the to without change dynamic for the loss in diameterof the helix , Wilms & Derksen took into that the rather of Nicotiana gradually play a static, passive longitudinal to differentiationand also a compensate explants occurs supposed or or occur mechanisms. transverse might and assumption requires one In tissue change that the helical and Pisum Raphanus the microtubules from directly by resisting turgor or microtubular skeleton behaviour cannot be reconciled with the assumptions The first second of the expansion, latter could has ceased. elongation out deposition, rigidity from cell result microtubules. The microtubule-based motors. coleoptiles these studies the these by which becomes stretched spring, 1985). Both the and coworkers assumed Lloyd et by organizations helical determined the control of cellulose occur generation involving less like is after the cells changes explantation. cell extension and has been related This to de- polarity. However, ethylene production by wounding & Derksen account the 1988). To explain dynamic properties in orientation resulted from plasma membrane, during a the mechanism involved, of the microtubules. polarity determining dynamic phase They factors in the of the microtubulesafter explantation. The behaviour of the cortical actin filaments described, (Derksen but the et al. during organization of the endoplasmic cell has elongation not yet been bundles remain essentially the same 1985). Differentiation Cell differentiation is different functions. deposition. Here we In the event plants, will discuss leading to cells differentiation is a few with quantitatively often examples, namely accompanied or by qualitatively a local wall vascular elements, stomatal cells and statocytes. The densities of microtubules increase thickening along the lateral walls of young gradually just before sieve elements in root the initiation of protophloem wall of wheat CYTOSKELETON SIGNIFICANCE (Eleftheriou skeletal 11 of the best studied one cell differentiation.In during in close relation with locally is 1987). Xylogenesis organization IN DEVELOPMENT elements cytoskeletal for Zinnia & Seagull (Falconer studied in and Seagull particular system (Fukuda & et al. These cells have 1988). Fukuda & The specific bands of microtubules of actin filaments the surface. over forming a transverse the sites of local wall at disappear orientations form bundles. The actin dots are co-ordinated way. bundles and wall of the deposition cells take least shape on of the 1984). the local guard tubules and cellulose cells 1989; cells and from radial, a prior in a role in the a guard mother depends at kidney-like the orientation of clear correlation between micro- be to to the firmly established. deposition of the pairs preceded by preprophase of guard Hardham changes and transverse the bands in the cells and the & 1989; Cleary of the microtubules over like in act Like in secondary 1986). & Wick the organization regions of microtubules reflects increases regular of microtubule results in the finally Palevitz & Mullinax 1989, and references in these the subsidiary the divisions of the radial division of the mothercell is development, pattern, cells. Their function subsidiary walls. In these cells density (Cho the destroys patterns. asymmetric of cortical microtubules and from mirror images until 1985). Cytochalasin disrupts which may indicate 1986), of wall material which cell wall. The radial arrays in both the Palevitz oblique, time the microtubules the microtubule microfibril orientation appears asymmetric, to changes gradually same a less axial distributed regularly Colchicine also Herth al. et grasses, Palevitz 1982; Kristen (reviews; radial organization new seen reached via are more or Microtubules and microfilamentsappear to organization periclinal vascular elements, microtubule The initial, deposition. cells and the deposition cells. The guard cellulose microfibrils in cell wall in development the reviewed this their orientation At the microtubule and actin filament place, forming partly wall deposition (see deposition (Dauwalder stomalal During initially are change In Zinnia, calmodulin is found between specific extensively present exclusively between the microtubule bands and are tubes (Derksen & Traas changes dots of actin actin filaments and also affects regular pattern of pollen large predominant. are finally located under the sites of cell microtubulepattern and cell wall the and Meanwhile the axial microtubules oblique elements xylem been Kobayashi, who recently network with the actin dots in the darns. This pattern and 1985). has also been Kobayashi 1989). characteristic sequence of actin and microtubule patterns. The patterns deposited Herth 1973, starts differentiate into which culture, 1988; Kobayashi Falconer & by cells elegans Roo deposition the cyto- to wall material is (Goosen-de The formation of bands of microtubules before local wall reported with respect cases xylem cells, pairs 1989; plane of of to to axial in the subsidiary Mullinax & papers). During from radial oblique, asymmetric, further transverse guard in cells. In Avena, in the last stage of differentiation, both guard cells and subsidiary cells show axial microtubule Mullinax 1989). Thus, patterns (Palevitz & microtubule patterns of Statocytes Lepidium situated in the top of the endoplasmic reticulum Sievers 1979). Hensel (1985, stabilizing co-ordinated show cell, are a a distinct way here cytoplasmic are 1987). involved seems Microtubules appear to be less endoplasmic to of the nucleus depend seems to reticulum on (Hensel actin filaments depend on a clear sequence in organization. whereas in the distal part of the present that too, in the different cells. in The cell, amyloplasts nucleus and as is rough graviperception (Volkmann typical organization is entirely actin dependent, the distal organization position This in occurs has been shown & by involved but may yet contribute in 1984). Thus, endoplasmic and not on reticulum microtubules. Also the actin filaments here, whereas in other systems it J. DERK.SEN 12 depends mainly role of the reticulum might is also thought depend Thus, cell polarity part on a depend to elements. cytoskeletal the distal to In tobacco the cortical must be from separated perhaps interchangeable continuous transport of statocytes (Hensel of the cell polarity each graviperception, endoplasmic 1985), graviperception i.e. microtubules cytoskeleton, explants, cells PLANT IN their change itself, however, (Hensel involving a 1985; 1987). differentpart the cell. In cells that do remains unchanged, shows meristematic not observed in Vinca shoot cortical a al. in explantation. Initially, the longitudinal different, are or even et al. randomly organized planes Selker 1988; Lang These orientations become by before the first Marc & Hacket meristem may become in aligned randomly centres (Lang microtubule as Selker patterns acid gibberellic 1989; Marc & Hacket between different cells Hedera (Marc & Hacket CONCLUDING 1989) to guard be involved The in the division Hedera tunica intercellularly. we in such a particular changes cytoskeletal Hormones factors cell to have reviewed specific in they to polarity actions vivo needed remains sequences that may explain as be are with other partly, hormone dependent, cytoskeleton appears as an indicator for the actual mechanisms involved. may to graviperception. used occur plant morphogenesis but open, in a highly early cell Changes co-ordinated way. at contacts. occur complex and still are also be involved. Whether The various types of cytoskeletal organization prove the versatile character of the different cyto- composition structures may least cell differentiation of tracheids appears of the various elements, their and their relation with other cellular probably skeleton least in the co-ordination within tissues and organs, but other affect skeletal elements. The actual in that will show the involvement of the present clearly sensory function cannot independent from direct cell specific at in different cells organization generally thought and the the The micro- clear co-ordination between and cell differentiation.The organization probably partake contacts are be at occurs cells, 1989). differentiation.However, in be, division mother cells promote the process of microtubule pattern formation in plant morphogenesis even may The been and Hedera REMARKS The localization studies cytoskeleton to seems 1989). with microtubules have also parallel longitudinal (1988) One of planes. almost tubular pattern in the tunica cells may indicate the outline of the leaf primordia develop axis of the orientation of the division planes. However, in Allium shown the occur, future division orientation of the division random are As 1989). microtubule pattern 1989). preferential microtubules et to oriented divisions. Meristematic apices (Sakaguchi & Hacket with regions (Mineyuki show (Marc parallel with microtubules that properties preferential do in to apparently randomly that also planes parallel the different microtubuleorientations tunica cells to but in three regions where cell divisions distributed and with having several times after transverse divide anymore, the orientation of the cortical microtubules not cortical microtubules will become regions MORPHOGENESIS polarity the orientation of microtubules shifts from the a distortionof this transport. The on CYTOSKELETON THE cells in occur to seems As cytoskeleton. of the these the versatile, and microtubules, indicating on various ET AL largely probably structures and dynamics and their mutual inter- unknown. The interactions of the cyto- transient and therefore will be difficult to CYTOSKELETON SIGNIFICANCE IN DEVELOPMENT determine from techniques also appears However, static images the basis for is set, cytoskeleton particular As several groups studies promises and must The latter may microtubular control of interaction with an the synthesizing complexes. of cytdskeletal also are for those botanists interested in the exciting detection and localiz- proteins. on via occur expression function of the cytoskeleton and function of the organization identification, discussion pending differentialgene studying are the to be to of the the light microscopic developed cells. cytoskeleton-associated control that a newly and that limits lateral diffusion of the cellulose plasmamembrane genetic deposition, of plant understanding interest also for the still cellulose microfibril in has been made in ation of intermediate filaments and the be of use promising further a a start as the alone, be very to 13 of cues and proteins the route, en near future plant morphogenesis cell differentiation. plant ACKNOWLEDGEMENT The authors preparation indebted are of the Professor M.M.A. Sassen for his critical support to the during manuscript. REFERENCES Akashi, T. & Shibaoka, H. (1987): the on microtubules Plant Cell Bajer, in & colchicine cells J. effects that not animal (ed.); Dynamic Effects of giberellin of cortical stability of internodes. pea 28: 329-348. Mole-Bajer, like but cold epidermis Physiol. A.S. plant and arrangement specifically microtubules. aspects with disassemble In: Soiter, ofmicrotubule biology. D. Ann. New York. Acad. Sci. 466: 767-784. — & — (1986b): Reorganization cells endosperm plant and cell of J., V. A. A.M. of the fragments Willemse, M.T.M. (1990):Organization during megasporogenesis (Mill.) H. phalloidin. 102: 263-281. & Lammeren, Gasteria with verrucosa rhodamine- analogues brain. ankyrin: associated are cultured Proc. cells and with Natl. Acad. Sci. with micro- USA 78: 7550-7554. Bergfield, Reorientation Speth, V. & Schopfer, of microfibrils and P. (1988): microtubules the outer epidermal wall ofmaize auxin-mediated Bot. Acta 101: 57-67. skeleton. and J. A.D. growth. & Vasiliev, In: Siekievitz, P.H. Organelles. Burgess, of & Plenum the nuclear Proloplasma 71: (1988): (ed.): Series Press, New (1970): Interactions York, between (1988): reticulum in 10:93-114. behaviour Dynamic cells. living Cell 54: 37-47. Cho, S-O tation & Wick, S.M. (1989); Microtubule stomatal differentiation during L. Clayton, Cyto- on Cellular microtubules in a ferm. & Lloyd, C.W. between the division Allium cells study. A.L. & (1984): and plane treated with anti-tubulin Cleary, orien- in grasses. J. Eur. The relationship spindlegeometry CIPC and griseofulvin: plexes in Lolium D.W., Cowan, N.J., (1980): A.R. (1989): Microtubule in the S. W.J. & Kirschner, evolutionary using J.S. cloned specific The (1974); tube Expt. and Cell of F-actin of Amarryllis Res. 88:435-439. Staehelin, acterization of novel and L.A. (1988): High features associated of the membrane pressure and char- endoplasmic systems. Eur. 46: 80-93. J. Cell Biol. Cronshaw, J., structural cDNA identification protoplast of intact plant tissues. Evaluation reticulum M.W. conservation of 20: 95-105. pollen & com- 149:67-81. Lopata, M.A., MacDonald, R.J., Rutter, p-tubulin Cell belladonna. Craig, rigidum: Protoplasma Number and and a- probes. in an J. Cell Biot. 34: 248-253, Hardham, organization during development of stomatal freezing London. envelope during mitosis 77-89, at coleoptiles during J.M. microtubules, mitoytic spindles C. Lee, endoplasmic Condeelis, R., Bershadsky, eukaryote flagella. Biosystems Cleveland, (1981): Erythrocyte mitotic structures in in higher of the actin skeleton visualized V. immunoreactive tubules in Acta Bol. Neerl. 39: 43-48. Davis, & Bennett, Duval in and L.B, of evolutionary origin phylogeny Cell Sci. 92: 581-594. microtubules Haemanthus in vivo. J. Cell Biol. Bednara, The (1978): and Chen, (1986a): Drugs T. Cavalier-Smith, Gilder, studies Nicoliana. J. J. in & Stone, D. Curcurbita, Ultraslrucl. Res. 45: (1973); Fine Cucumis and 192-205. 14 J. R.J. Cyr, & Palevitz, M Dauwalder, B.A. Microtubule bind- (1989): S.J S., Roux, Distribution of & Planta apices. root in Monoclonal J.S. and plumules with & higher plant (1985): filament anti- cells. J. Cell Biol. J„ Microtubules — in & and vegetative Ear. J. Cell J.A. Traas, Biol. control of cellulose B„ Wall Cell & Walls J.A. Traas, vitro: effects skeleton. (eds): Proc. Seed Plants, drugs Int. Groupe Growth Ferns of the with and & Sex. 4th Cell France. pollen the Went, in cyto- J.L. v. in Reproduction 64-67. Mosses. Pudoc, Jeucken, A.A.M. J.A. Traas, The (1986a): differentiatingroot tips & van microtubular of Raphanus Traas, bution J.A. & of actin filaments in J.H., Cove, D.J. Immunofluorescence in intact cell Sci. 75: Distri- differentiating cells of Plant Sci. 43: 77-81. & C.W. Lloyd, of the of (1985): microtubules Physcomitrella moss and CIPC-treated J. Cell cells. tip 131-148. Dustin. P. (1984): Eleftheriou. E.P. Microtubules. of Springer, Microtubules Berlin. and cell sieve differentiatingprotophloem Trilicum aestivum L. J. Cell Sci. 87: 595-607, Emons, microfibril (1985): A Planta mathematical deposition Reiss, D. and Proceedings 19B6. — Goldberg, of the Helicoidal growing stitution & study. Derksen, J. a in root hairs of R. (eds.): Wall Cell In: cell Vian, B., Walls 1986. Meeting, Paris France. microfibril Fosket, D.E. in Seagull, J. Acta Bot. Neerl. 39: (1988): Xylogenesis R.W. Continuing deposition during III: development. Protoplasma (1989): Cytoskeletal proteins Academic 393-461. M. Fragata, In; A. higher plants. Vol. Biochemistry of Plants, 144: of their and Marcus (ed.): 15: Molecular The Biology. London. Press, The mitotic (1970): site ofaction apparatus: a possible giberellic acid. Naturwissenschaften 57: 139-147. H. & H. Kobayashi, of the (1989): Dynamic cytoskeleton during tracheary- differentiation. Growth Develop. 31: Diff. 9-16. M.E. & Hardham, A.R. cell reorganization, of ment the 135: wall of axis of protoplasts the alga (1989): the On establish- regeneration Mougeotia. Protoplasma cells. J. filaments and cell The wall L.M. & with associated in antigens plant relationship between in primary Morphologi- 22: 279-300. Pollard, T.D. (1982): The filaments C.W. microtubule- thickenings Acta Bot. Neerl. aspects. tubule Lloyd, elements of the cucumber. I. tracheary of actin & of Cel! Sci. 93: 427-438, (1973): Roo, L. organelles AJ. distribution associated intermediate cal Microtubule and in elongation Goodbody, K.C., Hargreaves, cell (1986): synthesis 130-143. microtubules proteins. J. Biol. interaction and micro- Chem. 257: Grolig, F„ Williamson, R.E., Parke, J., Miller, C.H. streaming in a tip- alignment during tip dry-cleaving and freeze sub- (1986): Microfibrils, microtubules the trichoblast Neerl. 35: 311-320. of Equisetum B.H. in the (1988): Myosin alga peptides reacting and their B.E.S. morphogenesis primordium (ed.): Hardham, for initiation the a monoclonal cell 2+ in the & -sensitive poly- anti-myosin streaming endoplasm. 47: 22-31. (1981): in a Microtubules developing of Azolla pinata. Cytomorphogenesis Verlag, New —, and Ca Chara. Detection of two with localization Eur. J. Cell Biol Gunning, deposition Can. J. Bot. 67: 2401-2408. Acta Bol. Derksen, microtubules on 1016. Anderton, for helicoidal tip growth. 4th Cell and microfilaments in hyemale. wall. 163: 350-359. cell and microtubule morphogenesis: — rosettes model in cells with Groupe Paris, (1989): helicoidal colchicine 9143-9151. Plasma-membrane (1986): wall in do not control 113:85-87. Equisetum hyemale. — Microtubules orientation Protoplasma — (1982): of in root hairs. element tracheary Griffith, A.M.C. effect M.M. & Falconer, Goosen-de wall 117: 19-27. suspension (1987): development in elements The element (1986b): microscopy lineages Normal patens. T. micro- the cell wall of J.A. & Wolters-Arts, A.M.C., Traas, (1990): Galway, Oostendorp, Equisetum hyemale root tips. Doonan. —, skeleton Bol. Neerl. 35: 223-231. —, Cortical (1983): deposition in Equisetum hyemale. Protoplasma organization in coated cells. Eur. plant 68-81. Lammeren, sativus L. Ada A.M.C. tubules and microfibril Fukuda, G., and pits membrane of ETAL. 41:57-64. Wolters-Arts, genes Wageningen. —, Biol. Coated (1986): plasma in tissue culture microfibril of tobacco M.T.M. Symp. of Goldberg, R. Paris, interfering Willemse, 8th D. and Proceedings 1986. (1984): of In: 1986. Paris Meeting, cells 142-148. (1986): Cytoskeletal Reiss, (1985): generative 38: deposition. In: Vian, (eds.): — E.S. Pierson, pollen tubes. the and microfibrils 100: 1793-1798. Derksen, & on root hairs of C.W. Lloyd, to intermediate antibody cross-reacts gen in — J.A. Traas, J. Cell (1986): seedlings: pea & vesicles 168:461-470. Hulme, P.J., L. Hardison, calmodulin Immunocytochemical localization Dawson, — from carrot. Planta 177:245-260, ing proteins DERKSEN organ: In: and The cytoroot Kiermayer, O. in Plants. J.E. (1978): Evidences Springer York. A.R. & Hughes, of microtubules cortex in Azolla in descrete root-tip regions cells, and of an CYTOSKELETON SIGNIFICANCE hypothesis on microtubules. — & & development of cortical Planla Physiol. and moplasts Sci. G. & Microtubules. Ann. Rev. Imaseki, Inc. Boca of J. cytokinesis. Cell I wata, cal Cortical (1985): micro- for cell necessary A., Larkins, A (1989): Planla Dawson, fibrillar cross-reaction 166: 309-311. P.J., Butcher, K.C. Goodbody, monoclonal cytoplasmic its division? A.J., & bundles from carrot animal (ed.): C.W. cells, intermediate C.R. of the microscopy matrix in of culture. suspension Ear. cells Cell J. cells. the 38: — & tron J.C. Horne, of microscopy. wall glycol transmission R.M. Jr Brown, Orien- marine algae. Plant In: cytoskeleton: Lloyd, Growth and C.W. a Oriented cellulose critical The (ed.): Academic Development. in Press, of polarity — of from B —, 233-262. Hereward, revealed Oxford by W. Robards, 285-310. Hogetsu, root Cell T. of Howard, Res. plant Botanical plasmalemma Plant A.W. Oxford ed.: of cell (,Lepidium Y. reticulum cytoskeleton. of microtubule higher plants 13:621-635. In; 1985. Oxford. & Aist, of the J.R. & arrangement Alaska. Bot. 49: The cyto- 1-12. Movement and Cell London. in J. & Willemse, the actin skeleton Gasteria with verrucosa rhodamine- 178:531-539. Hepler, P.K. (1986): D.A. & freeze fixation of cells. plant in alata. freeze-substituted of freeze-substituted tubules ethylene and cells with plant of cells. 150: 72-74. Protoplasma W.R. Lang, J.M., Eisinger, of tubes pollen 140: 141-150. Protoplasma (1989): Immuno-gold labelling sections on P.K. (1987): Ultrastructure of Hepler, P.K. Hepler, the on cellulose & P.B. Green, orientation microfibrils polylamellatecell in Plant J.M. Lang Selker, (1989): apical epidermis cells R., Delorenzo, (1982): of of pea micro- epicotyl Proloplasma walls. 110; Microtubule preparations J., dependent kinase from in rat cold J. of & endo- and calmodu- stable brain. M. Vallano, Identification (1985): binding proteins in patterns of Vinca minor. Submitted. Goldenring, R. genous calmodulin microtubule Neurochem. 44: 1566-1574. Lichtscheidl, 27:939-945. ultrastructure — lin Immunofluores- var. & of Nicotiana Larsson, Fracture Microscopy (1986): In: 1985. formation. cytology: 5-14. Oxford. (1973): some wall L. cells: cytology: visualized rapid cytoskeleton Effects Microscopy University Press, of Pisum sativum RJ. structural 172: 377-384. D.H. Botanical Oshima, the roots endoplasmic University Press, microscopy cells of Cell Northcote, (1985); Physiol. Lepidium polar plant freeze-etch. J. Cell Sci. & the 129: 178-187. The (ed.): F.V. & of the planes cence (1985): A.W. Robards, Herth, and turnover statocytes. Exp. P.K. Hepler, in Zinnia Bot. 48:1-22. Prog. A.A.M., Bednara, Planla Cresti, M. actin cress (1987): Cytodifferentiation in root of roots affects from statocytes L.), Protoplasma formation microtubules 162:404-414. Cytochalasin of polarity sativum role statocytes sativum L. Planla (1985): A (1984): in cultured Prog. Cell Duval for of the differ- Protoplasma 31:153-165. the W. during (1989): Organization of A method (1988): disposition and molecular and molecular Lancelle, S.A., Callaham, comparison of Cytoskeleton General microtubules. (1986): H. (Mill.) H. Shibaoka, elements Allen & Unwin. van, C.W. 143: 29-37. Microfilaments. J.M. Lloyd, In: spatial during pollen development Cell London. — General phalloidin. Seagull, R.W. (1982): fibrils and the Hensel, (1987): M.T.M. of cell during regeneration giant 29: London. Press, H. & the tracheary (1986): Behaviour. 29: 785-793. models. U. Lammeren, (1988): meso- Physiol in Plant Growth and Devel- between cytoskeleton: Lackie, elec- 137:35-45. J. Micro. T. & selected I.B. & Plant Polyethylene tissues for of microtubules in Physiol. Heath, (1985): plant Hayano, S., Itoh, tation of skeleton: embedding Cell (1982): Actomyosin. Cytoskeleton Protoplasma Kristen, 201-210. — W.T. The opment. 3-29. Academic grown Biot. and coleoptiles Plant actin filaments and microtubules cytoplasmic L.) epicotyls. fila- and CRC Press (1988); Arrangement of corti- Avena Interrelation high voltage carota 249-264, and and cytoskeleton (.Daucus carrot and Pisum entiation Conventional (1985): T, in Kobayashi, H., Fukuda, ments. J. Cell Sci. 92: 371-378. Hawes, K. & Hogetsu, Jackson, against Takahashi, N. (ed.): In: Hormones, 807-805. G.W., Lloyd, raised antibody with Ullrastrucl. J. Florida. Raton, microtubules cotyls tubular lattices: absent from mature mesophyll and Hargreaves, (1985): Ethylene. H. Chemistry of Plant 157-179. F. freeze-substitution. Res. 66: 224-234. (1985): Preprophase bands, phrag- Hoffmann, 15 preservation by 698. spatial control Suppl.2: Hahne, 33:651 S.M. Wick, of arrays 143: 161-179. Hardham, A.R. (1982): Plant — the IN DEVELOPMENT of I.K. & Weiss, submicroscopic Allium cepa enhanced inner contrast D.G. structures (1988): Visualization in epidermal the cytoplasm cells light microscopy. Eur. of video- by J. Cell Biol. 46: 376-382. (1979): Hyphal tip fungus Fusarium: cell improved Lim, S.S., Hering, spread G.E. & occurrence Borisy, G.G. of anti-troponin T (1986): cross Wide- reactive 16 J. in components non-muscle cells. J. Cell Sci. 85: Lloyd, C.W. The (ed.) (1982): Development. Academic Press, Helical microtubular (1983): in Cytoskeleton in arrays Plant London. onion — Towards (1984); influence Int. — Rev. Cytol. 86: Actin in (1988); helical wall on model R. plants. patterns Sci. 90: for the P. plants. Higher visualized microtubules with 30 185-188. G. & antitubulin. of higher & J.A. Traas, studies. & J.L. plane of F-actin of carrot Development suspension S.M. Wick, surface and (1988): of Pleridium spermatozoid Monoclonal of antibodies of components & — Monoclonal (1988): tic and & 142: immunofiuorescent surface cell meristem assembly A localization to the and Protoplasma Wilson, L. disassembly method for of microtubules layers: application & shoot in apical (1978): Opposite of microtubules at end steady McCurdy, D.W., Sammut, of transverse cortical M. D. & siphonous movement tubules M. Shliwa, green & alga actin of arrays in wheat 147: 204-206. (1986): II. filaments. Eur. in the Chloroplast of both micro- J. Cel! Biol. 40: Meijer, E.G.M. &Simmonds, D.H. (1988): Interphase microtubule arrays Mineyuki, Y., Wick, Preprophase —, Marc, the J. & from Mita, T. Gunning, B.E.S. (1988); microtubules the nuclear cycle B.A. the cell in of onion band from guard mother random of cytoplasmic cells of Allium cepa L. 178: 291-296. & Shibaoka, 100-109. guard Siena, Italy, B.A. (1989): Microtubule band preprophase mother cells of Avena sativa L. 149: 89-94. Intermediate biology. Am. filaments: review a Pathol. Surg. J. Microtubule-associated (1986): B.A. of The (1982): stomatal 12 pro- C.W. Lloyd Growth and The (ed ): complex in cytoskeletal participation B. Mullinax, as a cell differen- Cytoskeleton Academic Development. in Press, (1989): Developmental changes of arrangement stomatal cells of Oat cortical (Avena microtubules sativa J., Miller, C.H. & Anderton, in Mold. monoclonal Eur. antibody. (1986): B.H. identified Higher plant myosin heavy-chain a Cell L.) 13: 170-180. Cytoskel. J. Cell using Biol. Parthasarathy, M.V., Perdue, T.D., Witztum, (1984): Giberellin stabilizes Protoplasma J. Alvernaz, Actin (1985); of the component B.M. & 41: J.D. Pickett-Heaps, Robards, Plant A. & normal a vascular many (1987): microtubule Nature 330: MAP1C. in as 1318-1323. R.B. Vallee, the transport by network cytoskeleton cells. Am. J. Bot. 72: Paschal, Retrograde associated protein 181-183. (1974): A.W. (ed.): Ultrastructure. Plant microtubules. Dynamic 219-255. Aspects of McGraw-Hill, Maidenhead. Pierson, E.S., Derksen, Organization in pollen —, Kengen, tubes of Eur. J. H.M.P. and J. & Traas, of microfilaments tubes grown angiosperms. actin in vitro & Nicotiana or in of higher associated The plant tabacum various Micro- (1989): L. Slabas, in pollen and Lilium 150: 75-77. A.R. & Lloyd, detergent-resistant cytoskeleton contains protoplasts fibrillar bundles in tubules. J. Cell Sci. vivo in co-localize longiflorum Thumb. Protoplasma (1982): (1986): 14-18. J. Derksen, filaments J.A. and microtubules Cell Biol. 41: Powell, A.J., Peace, G.W., C.W. H. of University accompanying (1988): J.B, tubules (1989); Development microtubules in onion leaf sheath cells. 119: and uncoupling 174:518-526. Palevitz, preprophase mesophyll proto- Plant. 72:511-517. experimental cells. Planta microtubules in Plama of and formation root-tip cultured Physiol. S.M. & bands kinetics cycle: their in of higher plants. in Int. I: 4-16. the In: 286-295. plasts & in plant Motility Bryopsis. requires organized arrays and B.E.S. Gunning, visualization actin microfilaments root-tip cells. Protoplasma Tenth 9-13. Immunofluorescent (1988); of London. Parke, 148: 70-79. vitro. Cell 13:1-8. state in Plant higher plant cells. new in tiation. In: fern — (1989): of Hedera. R.L. Margolis, Menzel, in a G., Reproduction of the Proc. Cai, Presence reproduction in higher plants. 1988. Palevitz, & basic Palevitz, of the mito- 15-24. W.P. Hackett, to components cytokinetic apparatus Protoplasma — antibodies novel R., teins. Ann. Rev. Cell Biol. 2:421-457. model spermatozoid detect June J.B. Olmsted, 142: 5-14. — the Suppl. the aquilinum. Protoplasma 312: Cresti, M., Gori, Sexual (eds): the sexual on R.B. Nagle, 102: 211-221. cytoskeletal Dynamic Nature (1988): tube. In: pollen E. May-4 Vignani, M. 205-209. Protoplasma in detercells. A., Cresti, Pacini, reorganization Gunning, B.E.S., Hardham, A.R., Perkin, Marc, J., to The role (1988): division & Plants. formation mining the Drug and Mullinax, 239-241. — (1984): growth. Springer Verlag, Berlin. Bradley, Nature 279: Tiezzi, A. in tobacco Symp. J. Cell. (1979): Cytoplasmic cells in A., Bartalesi, 1-35. —, Slabas,A., Powell, A., MacDonald, plant Moscatelli, kinesin dynamic a of microtubules M. Kirschner, microtubule ETAL. 237-242. root hairs. Nature 305:311313. & of instability Growth and — T. Mitchison, 1-19. DERKSEN nucleus- addition to 56: 319-335. micro- CYTOSKELETON Preston, R. D. (1988): Cellulose microfibril-orientating mechanisms in H. Quader, & of E, Planta 174:67-74. (1986): observation onion bulb in fluorescence adaxial scales. epidermis 131: Protoplasma 250-252. skeleton in ing G. & Cobaea seed hair cell-wall during E. Schnepf, Schnepf, U. & Herth, W„ Ryser, Planta E, skeletal elements in cotton seed hair their possible 168: A. Hoffmann, & of the movement bulb E. cell wall in involvement possible and onion of actin. 17-26. chemical in and Role of the microtubule tubes: pollen ultrastructural immunohisto- an approach. Biol. Cell helical by mediation D.G. & tubule-microfibril The H. syndrome. 164:439 109-126. Academic The (1982): In: Lloyd, in Plant Growth and Cytoskeleton (1985): reorientation: arrays. Planta Quader, K. Roberts, microtubule Ethylene-induced Robinson, & 447. C.W. (ed.): London. Press, B. & Mosiniak, cell walls and in Derksen, i S., of Vinca Salmon, E.D., M.L. & (1990): The actin skeleton tubes pollen of sea Nicotiana T. & Hara, N. (1988): labelled (1984): Spindle microtubule embryo’s; analysis using tubulin and measurement after laser vation P.J. & of Borisy, G.G. microtubule a of photo bleach- Direct obser- M.J. redistribution in Funaria. (1986): in dynamics cellular of living activation plasma-membrane A Tang, ion cells. Planta and channels vibrating-microelectrode cytoskeleton-inhibitor study. calcium Plant green of the at alga spatio- organizing 39: 29-42. M. Actin localization (1987): 141; W.E. bound J. Biochem. Molecular (1988); intermediate filaments. and Ann. Rev. P.K. and Scordilis, S.P. (1989): chain polypeptide in tubes. J. Cell Sci. 92: 569-574. & Vallee, R.B. Molecular (1982): of the c-AMP dependent protein to microtubule-associated Wick, B.E.S. of cellular J.A. & immunocytochemical identi- protein 2. R.E. & Chem. 257:3284-3290. S.C., Gunning, bound D.R. myosin heavy a pollen characterization Traas, Ethylene 67:460-466. Physiol. Hepler, of gration (1981): evidence of microtubule 57:593-625. X., Tiwari, A.V. tissue, Roop, biology of Theurkauf, S.M., Williamson, and (1984): Cytoskeleton function in cells of higher inte- plants. 99:635-695. (1984); Visualization of the cytoskeleton and coated of cleaving. means dry pits membrane of plant cells 119: Protoplasma 212-218. The In: Plant Braat, P. cortical root Membrane: Moller, Derksen, arrays cells J.W, during Berlin. (1984): Changes the of Rahanus (eds.): I.M. Structure, Function Biology. Springer, & microtubule C. and Larsson, Plasma and Molecular —, —, and 167:402-409. following cytokinin treatment Emons, A.M.C. & differentiation in of sativus. Eur. J. Cell Derksen, Microtubules in root hairs. J.W.M. (1985): J. Cell Sci. 76:303-320. Doonan, J.H., Rawlins, D.J., Shaw, P.J., Watts, & Lloyd, in the Biol. C.W. (1987): An actin network cytoplasm throughout the cells and —, associated the microtubule Chadwick, Immunochemical —, Cytokinin in Biol. 34:229-238. (1988): Nature 332:724-726. Saunders, and tem- systems higher plants. Proloplasma stem P.M. & skeleton. R.J., Saxton, W.M., Karow, J.R. in the shoot Planta 175:403-411. urchin & pea mediation. by J. Cell Biol. 99:2165-2174. Sammak, D.A. effects in J. Cell Biol. fluorescence redistribution ing. expression. of cortical microtubules in major L. Leslie, in fluoresceine internal (in press). Mclntosh, dynamics J. of Hogetsu, Arrangement apex of helicoidal versatility Planta Sakaguchi, level: supramolecular subprotoplasts tabacum. (1987): Morphogenesis of plant 140: 75-91. Protoplasma Rutten, T. & M. the at in sub- 1-12. kinase Roland, J.C., Reis, D., Vian, 8., Satiat-Jeunemaitre, geometry function microtubule example an of Schliwa, broad potential 24; 5320-5327. transition Acta 801. Need. Nicotiana Development. functional flagellar apparatus kupfferei: C.J. & fication micro- The flexibility Biochem. C.W. Roberts, 1.N., Lloyd, Plant Nore, P.S. & kinase: protein Biochemistry (1990): temporal and Rev. /Calmodulin-dependent centriole-dissociated Gloeomonas Staiger, 2+ and multi interphase-mitosis Steiner, 177-188. 61: tissues. P.J. Segaar, Steen, Louhelainen, J. (1987); cytoskeleton Ann. polarity. Ca (1985): specificity centers. (1987): Shape reticulum endoplasmic Eur. J. Cell Biol. 44: in 137: 56-62. Schnepf, cells: epidermis developmentin Raudaskoski, M., Astrom, K., Pertilla, K., Virtanen, I. & K.H. Spitzer, the (1987): Cyto- role regulatory organization. Proloplasma —, Cellular Schulman, H., Kuret, J., Jefferson, A.8., diverse development: pattern- differentiation. 152: Planta chlorotetracycline. 37: 23-47. Physiol. porary vitro: using Schnepf (1986): strate The cyto- (1986); 1-10, —, Funaria microtubule-associated Deichgraber, —, in 272-281. Endoplasmic cytoplasmic streaming: microscopical cells cell walls. plant Schnepf, reticulum and 17 SIGNIFICANCE IN DEVELOPMENT associates with the cell J. is present cycle of carrot dividingnucleus. J. Cell 105: 387-395. Burgain, S, organization & Dumas of the de Vaulx, R. (1989): cytoskeleton during The meiosis in 18 J. microtubules eggplant (Solanum melongena (L.)): and F-actin meiotic both are for necessary coordinated division. J. Cell Sci. 92: 541-550. plant I. cells. concomitant DERKSEN bands Preprophase of appearance stration in 48: in dry cleave demon- Eur. J. Cell Biol. preparations. 159-164. (1982): Tomato teins from a involved Trautwein, actin and land higher R.D., Rese, cation of T.S. & novel & Scordilis, contractile S. pro- Cell Motil. 2:131-147. Sheetz, force R. myosin: plant. M.P. (1985): Identifi- generatingprotein, kinesin, microtubule-based in B, motility. Cell 40: 559-569. organized —, Muto, kmesm-dnven microtubule ment. J. Cell Biol. Venverloo, C.J. of the plane & Libbenga, K.R. (1987): Regulation positioning Physiol. D. Plant & in multicellular Vol. organs. Physiology Springer, Wang, Y.D. teins Sievers, A. In: phragtnosome of Plant W. and Movements. Feinleib, N.S. 573-600, Berlin & cell S.M. & microscopy Gunning, of structurally J. Cell Biol. 89: & Duniec, labelling dividing plant Wilms, F.H.A. of cortical cells. & J. Double (1985): of calmodulin 126:198-206. Protoplasma Derksen, microtubules J. (1988): Reorganization during cell J. Orientation & Wolters-Arts, of cellulose 127-132. A.M.C. microfibrils in (1990): cells cortex Effects of microtubule-depoly- explants. merizing drugs. differentiation 146: explants. Protoplasma Derksen, immuno- and tubulin in Submitted. ation of cytoskeletal elements and cellulose Neerl. explants. micro- Acta 801. 39,49-52. Wulf, E., Deboben, A., Bautz, F.A., Faulstich, Wieland, T. (1979); Fluorescent for the visualization of cellular phallotoxin, actin. H. & a tool Proc. Nall. Acad. Sci. USA 76:4498-4501. Yan, of leaf cells. fibrils in cortex cells of tobacco Physiology in arrays plant & microscopy The 131:267-284. Haupt, L.P. (1988): membrane Tongbao (Science Bulletin) Wick, and I. (1979); Graviperception (eds): Encylopedia of 7, S. of tobacco of cell division in vacuolated cells formation. J. Volkmann, move- 107:2233-2241. function of nuclear M.E. meristematic fluorescence —, by microtubule K. 685-690. in tobacco works in vitro Immunofluorescence (1981); stabilized M.. Vahey, M., Titus, Vale, simultaneous cells: plant envelope- 97:235-243. —, Seagull, R., Osborn, M., Weber, microfibrils and develop nuclear tubulin. J. Cell Biol. associated ET AL. Duniec, J. The membrane pro- of Vicia faba. Kexue 33: 231-235. and microtubule G.Q. (1986): role (1983): Immunofluorescence of tubulin Yan, L.F., Wang, X.Z., Teng, X.Y., Ma, arrays in in the (Science Actin and growth Bulletin ) of myosin pollen in Y.Z. & pollen tubes. Kexue 31: 267-272. Liu, and their Tongbao