Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Genomic library wikipedia , lookup
Medical genetics wikipedia , lookup
DNA vaccination wikipedia , lookup
Genealogical DNA test wikipedia , lookup
Cancer epigenetics wikipedia , lookup
Minimal genome wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
Population genetics wikipedia , lookup
Biology and consumer behaviour wikipedia , lookup
Neocentromere wikipedia , lookup
Non-coding DNA wikipedia , lookup
Gene expression programming wikipedia , lookup
Genomic imprinting wikipedia , lookup
Point mutation wikipedia , lookup
X-inactivation wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Genetic engineering wikipedia , lookup
Extrachromosomal DNA wikipedia , lookup
Genome editing wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Gene expression profiling wikipedia , lookup
Dominance (genetics) wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Genome evolution wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
Genome (book) wikipedia , lookup
Helitron (biology) wikipedia , lookup
Quantitative trait locus wikipedia , lookup
Hybrid (biology) wikipedia , lookup
Designer baby wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
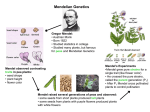
Mendel and After The science of Genetics began with Mendel’s classic experiments with garden peas in the 1850s. He presented his results as two lectures in 1865 and published them as a single paper a year later. The two fundamental laws of inheritance derived from his experimental results are known as Mendel’s Laws and the science on which they are based is known as Mendelian Genetics. Mendel is widely praised as a brilliant scientist, even a towering genius, but several authors have criticised his methods or the statistical validity of his data, a few have claimed he was a fraud. Gregor Johann Mendel (1822-1884) The most recent review, published in 2001, concluded that “his report is to be taken entirely literally . . . His experiments were carried out in just the way they are recounted . . . There is no credible evidence to indicate that he was inaccurate or dishonest in his description of his experiments or his presentation of his data.” Who was Mendel? Mendel’s life story is related in several biographies and innumerable biological textbooks. In outline: • Born in obscurity of humble, peasant stock he became a monk at the Augustinian Abbey at Brno, Austria. • Self-taught but with a keen interest in agriculture and horticulture he carried out numerous experiments on the hybridization of peas in the monastery garden. • From his results he deduced the two fundamental laws of heredity and the set of ratios that now bear his name. • He reported his results in 1865 at two meeting of the Brno Natural Science Society, where they were received in silence. • His now famous paper was published a year later in the Proceedings of the Brno Natural Science Society and ignored until his work was rediscovered in the beginning of the 20th Century. • Disillusioned by the reception of his paper Mendel gave up his research and in 1868 was elevated to the position of Abbot. In 1958 American science historian Loren Eiseley, in his book Darwin’s Century: Evolution and the Men who Discovered It, described the reception of Mendel’s paper by the Brno Natural Science Society: ‘At the end of the blue-eyed priest’s presentation of his researches, the still existing minutes of the Society indicate that there was no discussion . . . No one had ventured a question, not a single heartbeat had quickened. In the little schoolroom one of the greatest discoveries of the nineteenth century had just been enunciated by a professional teacher with an elaborate array of evidence. Not a solitary sole had understood him.’ In April 1965, the centenary of Mendel's paper, eminent evolutionist and Director of the Natural History Museum Sir Gavin de Beer declared on the radio: 'There is not known another example of a science which sprang fully formed from the brain of one man.' To an audience at the Royal Society he later delivered an address with the title, 'Genetics: The Centre of Science', in which he explained himself more fully: ‘It is not often possible to pinpoint the origin of a whole new branch of science accurately in time and place . . . But genetics is an exception, for it owes its origin to one man, Gregor Mendel, who expounded its basic principles at Brno on 8 February and 8 March 1865 in a (two-part) paper that would revolutionize the study of heredity. In a small monastic herb garden an obscure Moravian monk had personally discovered the basic principles of heredity that had eluded mankind for thousands of years . . . Mendel was a scientific revolutionary of the highest order.’ Mendel’s paper was not entirely ignored: it was cited at least 15 times between 1866 and 1899. But no one recognized it’s relevance to the science of inheritance until 1900, when three botanists independently carried out breeding experiments similar to those of Mendel and obtained similar results. They were Hugo de Vries in Holland, Carl Correns in Germany and Erich von Tschermak in Austria. De Vries cited Mendel’s paper in a footnote and is credited with its ‘rediscovery’. A fourth contender for the ‘rediscovery’ is the American William Jasper Spillman, whose paper on transmittance of parental characters in wheat, published in 1901, is seldom credited in this context. Mendel’s Experiments Mendel began his experiments in 1844 with 34 varieties of peas, which he subjected to two year’s trial so that he could identify varieties that were true-breeding for specific, easily recognizable characters. From these he selected 22 varieties that were true breeding, and 7 pairs of contrasting characters. For the next 8 years he carried out crosses between varieties with contrasting pairs of characters, studying in total about 10,000 plants. Mendel’s Laws Mendel’s laws are the Law of Segregation and the Law of Independent Assortment. The Mendelian ratios are 3:1, 1:2:1 and 9:3:3:1. Understanding these requires knowledge of a few technical terms – gametes, chromosomes, genes, and alleles – and some basic facts of reproductive biology. Gametes are eggs and sperms, the cells central to sexual reproduction. Chromosomes are strands of DNA and protein on which genes are located. A gene is a segment of DNA constituting a unit of heredity: it carries the code for a specific physical characteristic (unit character) such as seed colour in peas, eye colour in humans. Each gene can exist in several forms called alleles and a chromosome carries only one allele of each gene. Alleles of a gene occupy the same position, called a locus, on homologous chromosomes. If the alleles of a gene at a specific locus on homologous chromosomes are identical, the organism is said to be homozygous for that gene. If the alleles are different, the organism is said to be heterozygous for the gene. The term genotype refers to the identity of the alleles of a gene; phenotype refers to the physical characteristic determined by the combination of alleles. Alleles may be dominant, co-dominant or recessive. Meiosis The nuclei of all somatic (body) cells are diploid - they contain two sets of chromosomes, one set derived from each parent. Our somatic cells contain 46 chromosomes comprising two homologous sets of 23. Gametes are haploid – they contain one set of chromosomes. They are produced by a mode of cell division called meiosis during which homologous chromosomes exchange a random selection of genes in a process called crossing over, and then the cell divides twice to form four gametes, each with a single unique set of chromosomes. Mendel’s first law, the Law of Segregation, states that during gamete formation the alleles for each gene segregate so that each gamete carries only one allele for each gene. The allele for green pods is dominant over the recessive allele for yellow pods and so in the F1 generation all pods are green. A cross between a plant that is homozygous for green pods and one homozygous for yellow pods In the F2 generation the ratio of phenotypes is 3:1 and the ratio of genotypes is 1:2:1 The result of self fertilization of F1 plants that are heterozygous for green pods Mendel’s second law, the Law of Independent Assortment, states that genes for different traits segregate independently during the formation of gametes. In a 2-gene cross between plants homozygous for dominant round yellow seeds and plants homozygous for recessive wrinkled green seeds all the F1 plants are heterozygous for round yellow seeds. When the F1 plants are self fertilized the ratio of phenotypes in the F2 generation is: Round yellow Round green Wrinkled yellow Wrinkled green 9 3 3 1 Who wrote Mendel’s Laws? Despite what every textbook tells us Mendel’s Laws were not written by Mendel; they are implicit in his data but they are not explicitly stated in his paper. What became the first law was treated by Mendel as an assumption or hypothesis and it was Hugo de Vries in 1900 who first cited it as a law discovered by Mendel. In 1936 Sir Ronald A. Fisher, statistician and Darwinist, wrote: “Each generation finds in Mendel’s paper only what it expects to find; each generation has ignored what did not confirm its own expectations”. This remains true today. In 2001 science historian John Waller added: “It would be hard to imagine a finer demonstration of the cumulative effects of succeeding generations projecting the present back on to the past than is to be found in our current image of Gregor Mendel.” In the first generation of genetics textbooks following the rediscovery of Mendel their authors, including William Bateson (1902), S. Herbert (1910) and J. Wilson (1916), wrote vaguely about ‘Mendel’s Law’ as a general recognition of the segregation of unit characters into gametes, producing offspring in predictable proportions. In 1909 Bateson avoided the term law in favour of scheme, principle and phenomena. And in 1911 W. E. Castle condensed ‘Mendel’s Law’ into three principles: unit characters, dominance and segregation. In 1915 Thomas Hunt Morgan of Columbia University retained the singular ‘Law’ but a year later in his 1916 book, A Critique of the Theory of Evolution, he defined for the first time “Mendel’s first law – the law of segregation” and “the second law of Mendel, which may be called the law of independent assortment of different character pairs”. So Mendel’s laws as now universally accepted were first formulated by Thomas Hunt Morgan 50 years after the publication of Mendel’s paper. Morgan’s formulation of Mendel’s Laws was largely ignored for the next two decades, despite his receiving the Nobel prize in 1933. Then in 1938 A. F. Shull published his textbook, Heredity, in which he presented the two laws as formulated by Morgan, without ambiguity or attribution , as Mendel’s Laws, after which they became dogma in virtually all succeeding textbooks, and Morgan’s role in their formulation was forgotten. As for the Mendelian ratios, Mendel recognized the 3:1 and 1:2:1 ratios associated with segregation but the 9:3:3:1 ratio associated with independent assortment does not appear explicitly in his paper and it was Correns who first drew attention to it in 1900. What was Mendel really doing in the 1850s Mendel was not the self-taught peasant turned monk of many biographies. His family were farmers and he had a successful secondary education, cut short through lack of funds when his father suffered an accident and could no longer pay the fees. Mendel entered the monastery as a way of continuing his education and ultimately fulfilling his ambition to be a teacher. After he had completed his novitiate the monastery supported Mendel through two years at the University of Vienna, where he studied physics and maths under Christian Doppler and botany under Franz Unger. On returning to the monastery he worked as a supply teacher at the Brno Realschule teaching physics and natural sciences, became an active member of the Brno Natural Science Society and began his experiments with peas (and later with hawkweeds, beans and bees). For his experiments, rather than a ‘small monastic herb garden’, he had the use of a 2 hectare garden and a 27.5 x 4.5m greenhouse. His work was highly organized, his experiments were well planned and he obviously knew precisely what he was doing – but what was it? The clues are in the title and introduction of his 1866 paper. The title of Mendel’s paper is ‘Experiments in plant hybridization’ and in the introduction he states explicitly that he is presenting the results of a detailed experiment, the aim of which was to establish a ‘generally applicable law governing the formation and development of hybrids’. Mendel’s contemporaries would have understood clearly what he was about, but unfortunately for him the period of his experiments coincided precisely with a paradigm shift in the biological sciences brought about by the publication in 1859 of Darwin’s Origin of Species. The context in which Mendel was working was an intellectual dead-end and was soon forgotten. What Mendel was doing was systematically exploring an issue that had preoccupied the botanical community for the previous century, known as ‘species multiplication by hybridization’. Mendel was the last of a line of botanists convinced that they could produce what had become known as ‘constant hybrids’, essentially hybrids that had become new species. The concepts of dominance and segregation now attributed to Mendel were well known to his predecessors in the field. The possibility that new species might arise by hybridization was proposed in the 1750s by the great Linnaeus, originator of the system of classifying organisms in terms of species, genus and family. His classification scheme was founded on the plants and animals of Europe but became overwhelmed by the quantity and wonderful variety, particularly of plants, that were being brought back from around the world by explorers. Linnaeus began to think the hitherto unthinkable: had God really made all these species in one creative episode or had he arranged things so that new species might arise by cross-breeding among an originally much smaller suite of primordial forms? This idea was not deemed heretical because cross-breeding between plant species is a common and well documented phenomenon, but Linnaeus took it a stage further – he speculated that if interbred through several generations such hybrids might eventually become new species. Over the next century this idea consumed a great deal of scientific effort – major prizes were offered for success but never claimed. Repeatedly investigators found themselves unable to stabilize hybrid forms, whose offspring either reverted to parental type or died out through low fertility. Despite the consistent failures many botanists remained convinced that it would be possible to produce new species by hybridization. One such botanist was Franz Unger, Mendel’s professor at Vienna, who had assured Mendel that hybridization might well be a source of new species. So what Mendel was doing was trying to produce ‘stable hybrids’ and, as he stated in his paper, to find a ‘generally applicable law governing the formation and development of hybrids’. Not only did he fail, he proved conclusively that no hybrid lineage is capable of forming only hybrid offspring. Mendel had spent a decade working on his experiments, had received support from the abbot and practical help from fellow monks, and had discussed his work with other botanists as well as fellow members of the Brno Natural Science Society. Accordingly we can assume that his work and what he hoped to achieve would already have been well known to many in the audience when he presented his two lecture to the Society in 1865. If the myths that have accumulated round Mendel are removed we can see that he was not expounding his discovery of the laws of heredity but was actually reporting unequivocal failure. He had produced interesting statistical patterns that he couldn’t fully explain but had failed in his aim to stabilize plant hybrids. Given that many of the audience already knew what he was about, we can reread their silence as reflecting sympathetic understanding rather than incomprehension. Mendel had devoted most of his scientific life to what proved to be an intellectual dead end. He thought his data applied only to hybrids and he had no understanding of their significance to the science of heredity. By the time his work was rediscovered the scientific context in which he was working had been forgotten. After Mendel – significant discoveries and key concepts • 1878 - Walther Flemming discovered chromosomes and the process of mitosis, though it was 10 years later that chromosomes were named by Wilhelm van Waldeyer. • 1883 – Edouard van Beneden discovered the process of meiosis. • 1886 - August Weissman proposed his germplasm theory , that inheritance only takes place by means of the germ cells—the gametes. Other cells of the body—somatic cells—do not function as agents of heredity. In 1890 he recognized the significance of meiosis for reproduction and inheritance, and the role of chromosomes in passing on hereditary information. • 1886 - Hugo de Vries discovered new forms of evening primrose flowers, showed that the traits were transmitted to the next generation via the seeds, and introduced the term mutation. • 1889 - Hugo de Vries proposed that different characters have individual hereditary carriers and that inheritance of specific traits comes in particles, which he called pangenes. Pangene was later abbreviated to gene by Wilhelm Johannsen in 1909. • 1894 - Eduard Strasburger recognized the haploid/diploid distinction and its role in reproduction. • 1903 - Walter Sutton proposed that chromosomes are implicated in Mendelian inheritance and carry the units of heredity. • 1905 - William Bateson coined the word genetics and is regarded as one of the founders of the discipline. Born in Whitby, he was a Fellow of St John’s, Cambridge and from 1910 Director of the John Innes Institute. • 1908 - Sir Archibald Garrod of St Bartholomew’s Hospital, London, reported that the disease alkaptoneuria, which runs in families, follows a Mendelian pattern of inheritance and is caused by “an inborn error of metabolism”. He went on to show that the cause is a defective enzyme and deduced that the failure in the enzyme results from a change in a gene. He coined the adage: “one gene, one enzyme”, which was verified in the 1940s by Beadle and Tatum of Stanford University and then modified by Linus Pauling in the 1950s to “one gene, one protein”. • 1909 - William Bateson introduced the term allelomorph, later abbreviated to allele. • 1911 - Wilhelm Johannsen introduced the terms genotype/phenotype and homozygous/heterozygous. • 1911 - Thomas Hunt Morgan discovered crossing-over in chromosomes during meiosis in Drosophila. 1915 – Thomas Hunt Morgan and his students published The Mechanism of Mendelian Heredity, in which they presented a synthesis of their work, unifying the genetic research of cytologists and breeders. They had established that the Mendelian unit characters of the breeders are the genes, which exist as discrete points on the chromosomes of the cytologists. Genes can travel into the next generation separately, as Mendel had described, if they are on different chromosomes, or together if they are on the same chromosome, though they might still be transmitted separately as a result of crossing-over. This work introduced the famous metaphor of a chromosomes as a linear series of beads, with the beads corresponding to genes. By the 1920s it was established beyond reasonable doubt that chromosomes are involved in heredity and that they carry the hypothetical abstractions known as ‘genes’. But what are genes? Chemical analysis of chromosomes had shown that they are composed of approximately equal proportions of protein and what is now known as DNA, deoxyribonucleic acid. Both are macromolecules composed of chains of subunits, but which of them carries the hereditary information? Early opinion strongly favoured proteins: they are astonishingly versatile in function and correspondingly variable in structure and so the underlying genetic code must be equally versatile. As polymers composed of up to 20 different amino acids, theoretically at least, proteins have such versatility. In the early 1900s the composition of DNA had been determined by Phoebus Levene of the Rockefeller Institute for Medical Research. He identified three components and showed that they were linked in the order phosphate-sugar-base to form units that he called nucleotides. He identified the sugar as desoxyribose. Levene identified four bases – adenine, cytosine, guanine and thymine (A, C, G, T) which he thought were present in equal proportions. In 1910 he proposed his tetranucleotide hypothesis, that the four bases are linked to form a tetranucleotide and that DNA comprises a chain of identical repeating tetranucleotides. Such a molecule is too simple to carry genetic information and so the protein component of the chromosomes must be the basis of heredity. Thereafter most research on the nature of genes focussed on protein until the 1940s. In 1944 Erwin Shrödinger published his influential book, What is Life?, in which he looked at the phenomenon of life from the viewpoint of physics. In addressing the problem of heredity he proposed that the hereditary material is a molecule with a structure that does not repeat itself. He called it an aperiodic crystal, whose aperiodic nature allows it to encode an almost infinite number of possibilities from a small number of atoms. The ‘blueprint’ of life would be found in a molecule whose structure had something of the regularity of a crystal but must also embody a long irregular sequence, a chemical structure capable of storing information in the form of a genetic code. He believed that a protein was the obvious candidate, with its varying amino acid sequence supplying the code. Coincidentally, also in 1944 the first experimental evidence pointing unequivocally to DNA as the stuff of genes was published, in what is now a classic paper, by Oswald T Avery and his associates of the Rockefeller Institute, from their work on Pneumococcus bacteria. They had shown that hereditary traits could be transferred from one bacterial strain to another by purified DNA, but their paper was received with incredulity and raised a storm of controversy. Avery and his associates were medical microbiologists and doubt was cast on their biochemical competence, the efficacy of the enzymes used to digest protein from their DNA extract and the purity of the DNA. The verdict from one Nobel Prize winning geneticist was that ‘their so-called nucleic acid is probably nucleoprotein . . . with the protein too tightly bound to be detected by ordinary method’. Support for Avery came from biochemist Erwin Chargaff, who said he didn’t give a damn for what the geneticist thought of him, he was deeply impressed by Avery’s work. If Avery was right and DNA is the molecule of heredity then there should be demonstrable differences in proportions of the four nucleotides between species, such as a horse and a cat or a mouse, and in 1950 he proved this to be the case. He also showed that the four nucleotides are not present in equal proportions (thus disposing of Levene’s hypothesis) but that the proportion of adenine always equals that of thymine, that of cytidine equals that of guanine. Evidence that finally convinced most of the doubting geneticists that DNA carries the genetic code came in 1951 from an elegant experiment carried out on bacteriophage viruses by microbiologists Alfred Hershey and Martha Chase. A phage was known to consist of a core of DNA surrounded by a protein coat and that it inject its genetic material into a bacterium where it takes over the machinery of the cell to produce new phages. By using radioactive tracers Hershey and Chase were able to show that it is only the viral DNA that enters the bacterium, the protein being left behind in the empty viral coat. With DNA established as the carrier of the genetic code and its composition determined through the work of Levene and Chargaff the final clue to its structure came from the X-ray diffraction studies of Malcolm Wilkins, Rosalind Franklin and Raymond Gosling at King’s College, London, which showed conclusively that the DNA molecule has a helical structure. In February 1953 chemist Linus Pauling published a paper proposing that DNA has a triple helix with a phosphate-sugar spine at the centre. This interpretation was incorrect and what is now universally accepted as the correct one was published shortly after on 25th April 1953 by James Watson and Francis Crick, who proposed that the DNA molecule consists of two helical of chains of phosphate-sugar groups that run in opposite directions and are linked by paired bases – Morgan’s metaphorical string of beads was resolved as the famous double helix and genes are short sequences on a DNA molecule. Other key discoveries in genetics: Chromosomes as determinants of sex – in many animal groups sexual differentiation is controlled by specific sex chromosomes. In mammals these are designated X and Y, with females having XX and males XY. In birds the situation is reversed. Sex linkage relates to genes that are located on the sex chromosomes. These genes are considered sex-linked because their expression and inheritance patterns differ between males and females, as in red-green colour blindness, which results from a defect in a recessive gene on the X chromosome and is commoner in males. Polygenic characters – controlled by several genes – height or hair and skin characteristics. Pleiotropic genes – responsible for or affect more than one phenotypic character: • Cystic fibrosis – lungs, pancreas, liver and kidneys. • Albinism – caused by a mutation in the gene for tyrosinase, which affects the production of melanin – affects skin, hair, eyes – light sensitivity. Polyploidy – more than 2 sets of chromosomes – rare in animals, very common in plants – 99% of ferns, 50 – 75% of flowering plants, including almost all crop plants. • • • • • Triploid – apples, bananas Tetraploid – apples, durum wheat potatoes, cotton Hexaploid – bread wheat, oats, Chrysanthemums Octoploid – strawberry, sugar cane, Dahlias 48-ploid (1440 chromosomes) in an Adder’s Tongue fernOphioglossum reticulatum Ironically for Mendel’s aspirations, new plant species commonly do arise through hybridization followed by polyploidy. References Fairbanks, Daniel J., Bryce Rytting (2001): Mendelian controversies: a botanical and historical review. American Journal of Botany, 88, 737-752. Franklin, Allan, A. W. F. Edwards, Daniel J. Fairbanks, Daniel L. Hartl (2008): Ending the Mendel-Fisher Controversy. University of Pittsburgh Press. Marks, Jonathan (2008) The construction of Mendel’s laws. Evolutionary Anthropology, 17, 250-253. Monaghan, Floyd, Alain Corcos (1984): On the origins of the Mendelian laws (1984): Journal of Heredity, 75, 67-69. Olby, Robert C. (1985): The Origins of Mendelism. University of Chicago Press. Olby, Robert C. (1997): Mendel, Mendelism and Genetics. http://www.mendelweb.org/MWolby.html Ryan, Frank. (2016). The Mysterious World of the Human Genome. William Collins Waller, John (2002): Fabulous Science, Oxford University Press. Chapter 7, The priest who held the key: Gregor Mendel and the ratios of fact and fiction