Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cell encapsulation wikipedia , lookup
Cell culture wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cytokinesis wikipedia , lookup
Phosphorylation wikipedia , lookup
Signal transduction wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cell growth wikipedia , lookup
Endomembrane system wikipedia , lookup
Protein phosphorylation wikipedia , lookup
Biochemical switches in the cell cycle wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
List of types of proteins wikipedia , lookup
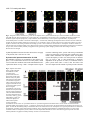
1929 Journal of Cell Science 113, 1929-1938 (2000) Printed in Great Britain © The Company of Biologists Limited 2000 JCS1193 Chromatin-bound Cdc6 persists in S and G2 phases in human cells, while soluble Cdc6 is destroyed in a cyclin A-cdk2 dependent process Dawn Coverley*, Cristina Pelizon, Sarah Trewick‡ and Ronald A. Laskey Wellcome/CRC Institute, University of Cambridge, Tennis Court Road, Cambridge, CB2 1QR and Department of Zoology, University of Cambridge, Cambridge, UK *Author for correspondence (e-mail: [email protected]) ‡Present address: ICRF, Clare Hall Laboratories, South Mimms, Herts, EN6 3LD, UK Accepted 8 March; published on WWW 10 May 2000 SUMMARY Cdc6 is essential for the initiation of DNA replication in all organisms in which it has been studied. In addition, recombinant Cdc6 can stimulate initiation in G1 nuclei in vitro. We have analysed the behaviour of recombinant Cdc6 in mammalian cell extracts under in vitro replication conditions. We find that Cdc6 is imported into the nucleus in G1 phase, where it binds to chromatin and remains relatively stable. In S phase, exogenous Cdc6 is destroyed in a process that requires import into the nucleus and phosphorylation by a chromatin-bound protein kinase. Recombinant cyclin A-cdk2 can completely substitute for the nucleus in promoting destruction of soluble Xenopus and human Cdc6. Despite this regulated destruction, endogenous Cdc6 persists in the nucleus after initiation, although the amount falls. Cdc6 levels remain constant in G2 then fall again before mitosis. We propose that cyclin A-cdk2 phosphorylation results in destruction of any Cdc6 not assembled into replication complexes, but that assembled proteins remain, in the phosphorylated state, in the nucleus. This process could contribute to the prevention of reinitiation in human cells by making free Cdc6 unavailable for re-assembly into replication complexes after G1 phase. INTRODUCTION Romanowski and Madine, 1996, 1997; Rowles and Blow, 1997; Stillman, 1996; Wuarin and Nurse, 1996). Assembly takes place sequentially. ORC, which occupies predetermined sites of initiation in yeast, recruits Cdc6, which in turn directs loading of the Mcm complex (Cocker et al., 1996; Coleman et al., 1996; Donovan et al., 1997; Romanowski et al., 1996; Rowles et al., 1996; Tanaka et al., 1997). Together they form a pre-replication complex (pre-RC), which is activated at the start of S phase. Analysis of the proteins associated with yeast replication origins in cyclin-dependent kinase (cdk) mutants, and of the behaviour of the Mcm proteins in the presence of protein kinase inhibitors in mammalian cells (Coverley et al., 1998; Dahmann et al., 1995; Piatti et al., 1996), suggest that phosphorylation events at the start of S phase are required for activation of the pre-RC and that continued kinase activity throughout S and G2 prevents reassembly of new complexes. The proteins of the pre-RC generate a specific DNase I protection profile in G1, which becomes less extensive in S and G2 phases (reviewed in Diffley, 1996). This is consistent with destruction or displacement of some of the proteins involved in initiation. In Xenopus, yeast and mammalian cells, Mcm proteins are displaced from chromatin as cells proceed through S phase (Chong et al., 1995; Krude et al., 1996; Kubota et al., 1995; Madine et al., 1995b; Todorov et al., 1995) and in yeast, Cdc6 protein levels decrease (Cocker et al., 1996; Jallepalli et Duplication of nuclear DNA is strictly coordinated with the rest of the cell cycle so that it occurs only once before the cycle is reset at mitosis. Regulated initiation of mammalian DNA replication can now be achieved entirely in vitro using soluble protein extracts made from human cells synchronised in S phase. A single round of DNA synthesis can be induced in nuclei from HeLa cells chemically synchronised in late G1 (Krude et al., 1997) or nuclei from mouse cells released from quiescence into G1 (Stoeber et al., 1998). In the latter case the proportion of nuclei which begin DNA synthesis is greatly increased by supplementing S-phase extracts with recombinant Cdc6 protein. Cdc6 was first identified in yeast (Zhou et al., 1989), but homologues have been found in higher eukaryotes including Xenopus (Carpenter et al., 1996) and humans (Hateboer et al., 1998; Saha et al., 1998; Williams et al., 1997). All evidence suggests that it is a key regulatory protein, which is essential for initiation of DNA replication (Carpenter et al., 1996; Cocker et al., 1996; Coleman et al., 1996; Hateboer et al., 1998; Kelly et al., 1993; Yan et al., 1998). Cdc6, the origin recognition complex (ORC) and the Mcm family of proteins assemble at sites of future initiations in G1 in a regulated process known as replication licensing (reviewed in Botchan, 1996; Diffley, 1996; Muzi-Falconi et al., 1996; Newlon, 1997; Key words: Cdc6, Proteolysis, S phase, Cyclin-dependent kinase 1930 D. Coverley and others al., 1997; Nishitani and Nurse, 1995; Piatti et al., 1996, 1995). In higher eukaryotes the fate of Cdc6 after initiation is still controversial. There is confusion in the literature about the behaviour of endogenous human Cdc6 in S phase. A number of reports looked at the total level of endogenous Cdc6 in the cell cycle by western blot analysis in Wi38 cells (Williams et al., 1997), CV1 cells (Petersen et al., 1999), 3T3 cells (Saha et al., 1998) and U2OS cells (Jiang et al., 1999). They found that apart from a drop in level during quiescence and during mitosis the total amount of Cdc6 protein remains relatively stable in late G1, S and G2 phases. These observations are not in question. However, analysis of the subcellular distribution of Cdc6 upon transition from G1 to S phase has yielded conflicting data. Immunofluorescence data showing the subcellular distribution of overexpressed Cdc6 in U2OS cells (Jiang et al., 1999; Petersen et al., 1999; Saha et al., 1998) or endogenous Cdc6 in CV1 cells (Petersen et al., 1999) find that nuclear staining is present in G1 but absent in S phase. Total levels of endogenous protein remain fairly constant, so loss of S-phase staining has been linked to increased cytoplasmic staining. There is some evidence in favour of increased cytoplasmic Cdc6 in S phase (Jiang et al., 1999; Petersen et al., 1999; Saha et al., 1998). Contradictory to these observations, several studies report persistent Cdc6 in the S-phase nucleus. In nuclei from 3T3 cells, Cdc6 remains present beyond G1 (Stoeber et al., 1998) and in HeLa cells the level of nuclear bound Cdc6 was unchanged between G1, S phase and G2 (Fujita et al., 1999) although cytoplasmic Cdc6 became apparent in S phase. In addition, nuclear Cdc6 has been detected by immunofluorescence in S phase in newborn human fibroblasts and in 90% of nuclei in premalignant human cervix tissue (Williams et al., 1998). Our analysis shows conclusively that Cdc6 persists in human nuclei beyond S phase and also demonstrates the existence of two isoforms, one of which is unstable in isolated nuclei. The relevance of these isoforms to the apparent conflict in the literature is discussed. In addition, by monitoring the fate of exogenous free Cdc6 in vitro, we extend the results obtained in overexpression experiments (Jiang et al., 1999; Petersen et al., 1999; Saha et al., 1998). Free Cdc6, not assembled into replication complexes, is destroyed by proteolysis in a reaction triggered specifically by cyclin A-cdk2 in the S-phase nucleus. Thus reassembly of replication complexes in S phase is prevented, allowing only one round of replication within a single cell cycle. MATERIALS AND METHODS Cell culture and synchronisation HeLa cells, NIH 3T3 cells and Wi38 cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 10% foetal calf serum, 10 i.u./ml penicillin and 0.1 mg/ml streptomycin at 37°C. Sf9 insect cells were grown in Grace’s medium containing 10% foetal calf serum and 5 µg/ml gentamycin (all reagents from Gibco-BRL). HeLa cells were synchronised in S phase by double thymidine block (0.4 mg/ml) of 17 hours and 15 hours separated by a 9-hour release. To generate a G1 population, pre-synchronised cells were arrested in mitosis by 40 ng/ml nocodazole (Sigma) and released into G1 by mitotic shake off. NIH 3T3 and Wi38 cells were synchronised in G0 by contact inhibition as previously described (Stoeber et al., 1998). G1 phase populations were achieved by releasing confluent cells from quiescence by subculturing and replating 1 in 6 into standard medium for 16-18 hours. Synchronies were checked by flow cytometry and by in vitro elongation assays, which report on the proportion of nuclei in S phase. Preparation of nuclei and cytosolic extracts Nuclei were prepared as previously described (Krude et al., 1997). Briefly cells were washed and swollen in hypotonic buffer then scraped from the plate and disrupted by 10 light strokes in a dounce homogeniser, not 25 as previously described. In some experiments homogenisation was done in the presence of 1 µM okadaic acid (Calbiochem). Nuclei were pelleted and the supernatants used to produce cytosolic extract. Nuclei were washed and stored as previously described (Stoeber et al., 1998). Supernatants or homogenised mitotic cells were spun at 14,000 rpm for 20 minutes to remove residual insoluble material and frozen in aliquots in liquid nitrogen. All materials were frozen and thawed only once. Production and purification of Cdc6 Sf9 cells were infected with a 6×His-tagged Xenopus Cdc6 recombinant baculovirus (Coleman et al., 1996) or a derivative in which the serine residue in all five Cdk consensus phosphorylation sites were changed for alanines. This mutant is designated M9 (C. Pelizon et al., manuscript in preparation). A baculovirus expression construct coding for 6×His-tagged human Cdc6 was produced by inserting the coding sequence for human Cdc6 (from GenBank) into the BamHI site of vector pVL-6His. Recombinant protein was purified using a nickel affinity column according to published procedures (Kunagai and Dunphy, 1995). Column fractions containing Cdc6 were pooled and dialysed against 20 mM Tris, pH 7.4, 150 mM NaCl, 5% glycerol, 1 mM DTT then stored at −80°C. Antibodies Anti-Xenopus Cdc6 polyclonal antibody was raised in rabbits using full-length recombinant Xenopus Cdc6 as antigen and purified by standard procedures (Harlow and Lane, 1988). Purified anti-Xenopus Cdc6 antibody was used at 1:1200 dilution for immunofluorescence studies, and under these conditions did not cross-react with human or mouse Cdc6. The anti-human Cdc6 polyclonal antibody has been published previously by this laboratory and was produced and purified as described (Stoeber et al., 1998). Actin in isolated nuclei was detected on western blots using affinity-purified rabbit anti-peptide actin antibody (Sigma). Nuclear import, protein degradation and phosphorylation reactions Cdc6 (100 ng) was incubated with or without intact nuclei (1-5×104) in 10 µl of cytosol, for analysis by western blot. Reactions were scaled up 2-3 times for immunofluorescence studies. Cytosols were prepared and supplemented as for replication reactions (Krude et al., 1997) with 1.3 mM MgCl2, 20 mg/ml creatine phosphokinase and a 10× frozen reaction pre-mix (to give 40 mM Hepes, pH 7.8, 7 mM MgCl2, 3 mM ATP, 0.1 mM GTP, 0.1 mM CTP, 0.1 mM UTP, 0.1 mM dATP, 0.1 mM dGTP, 0.1 mM dCTP, 0.5 mM DTT, 40 mM creatine phosphate) just before use. Reactions were incubated for 30 minutes, or as specified in figure legends, at 37°C. Reactions were variously supplemented with 0.5 mM olomoucine (Sigma), importin β dominant negative mutant (Kutay et al., 1997), 100 nM leptomycin B (Nishi et al., 1994), 0.05 mM MG132 (Sigma), p21 (Furuno et al., 1999) or sf9 cell lysates containing human cyclin-cdk complexes. Protein kinase activity levels in unpurified sf9 lysates made from cells infected with human cyclin A and cdk2 or cyclin E and cdk2, or cyclin D and cdk6 baculovirus constructs, were normalised by their ability to phosphorylate the retinoblastoma protein in standard kinase assays. Cdc6 in S phase in human cells 1931 5×105 Rb phosphorylating units were added per 20 µl reaction. Reaction products were analysed in one of two ways. For immunofluorescence studies, reactions were diluted 50-fold in phosphate-buffered saline (PBS) or PBS/0.5% Triton X-100 and then fixed with an equal volume of 8% paraformaldehyde. Nuclei were spun through 30% sucrose in PBS onto polylysine-coated coverslips as described (Mills et al., 1989). Exogenous Cdc6 was detected using Xenopus specific purified polyclonal antibody in ‘antibody buffer’ (0.1% Triton X-100, 0.02% SDS, 10 mg/ml BSA in PBS) and visualised using anti-rabbit-FITC (Vector labs). DNA was counterstained with propidium iodide for visualisation by fluorescence confocal microscopy and digital images were merged using Adobe photoshop. For immunoblotting, whole reactions were stopped by the addition of SDS-PAGE loading buffer and heated to 95°C for 10 minutes. Reaction products were separated by electrophoresis through 7 or 8% polyacrylamide gels and transferred to nitrocellulose. Blots were developed with polyclonal anti-Xcdc6 or polyclonal anti-human Cdc6 and anti-rabbit conjugated to horse radish peroxidase (Sigma), using standard protocols. Blocking and antibody incubations were in 0.4% Tween 20, 10% low-fat milk powder in PBS. Bands were visualised using Enhanced ChemiLuminesence (ECL, Amersham) and preflashed Kodak film. Quantification of band intensities was done using NIH image software and short ECL exposures. Replication initiation and elongation assays Nuclei (approximately 1×104 per 10 µl reaction) were incubated at 37°C for 2 hours in 75 mM NaCl, 250 mM sucrose, 0.5 mM spermine, 0.15 mM spermidine, 3% BSA for elongation assays or S-phase cytosolic extract for initiation assays supplemented with ATP and ATP-regenerating system and 10× nucleotide mix as above. In addition reactions were supplemented with biotinylated dUTP, which was detected after fixation in paraformaldehyde, using streptavidinFITC (Mills et al., 1989). Replication is expressed as the percentage of nuclei in a population that show fluorescence. RESULTS Exogenous Cdc6 accumulates in intact G1 nuclei We first asked whether Cdc6 enters the nucleus under in vitro replication conditions. We chose to use purified recombinant Xenopus Cdc6 protein in these studies for three reasons. Firstly, because we know it stimulates replication in mammalian nuclei in vitro (Stoeber et al., 1998), secondly because we have a mutant form in which all five consensus cdk phosphorylation sites are mutated and thirdly, the ease with which Xenopus Cdc6 is detected in immunofluorescence assays or by western blot using species-specific polyclonal antibodies that do not recognise either the endogenous human or mouse proteins. Using G1-phase cytosol prepared from 3T3 cells 16.5 hours after release from quiescence we monitored the accumulation of exogenous Cdc6 in nuclei from the same cells. In immunofluorescence assays (Fig. 1A) we found that Cdc6 efficiently accumulated in G1 in vitro. In order to prove that accumulation is not the result of diffusion through a damaged nuclear membrane, we included a dominant-negative mutant of the import factor importin β (Kutay et al., 1997). The mutant protein completely abolished accumulation of Cdc6 in the nucleus (+ import inhibitor). This provides an important control for nuclear membrane integrity. In addition, this data is strong evidence that Cdc6 is actively transported into G1 nuclei and is consistent with reports of a functional nuclear localisation signal (NLS) (Petersen et al., 1999; Takei et al., 1999). We then asked whether imported Cdc6 binds to chromatin. Accumulated Cdc6 resisted extraction with detergent (+TX100 wash) showing that it binds to structures within the nucleus, probably chromatin. Furthermore, detergent extraction did not significantly decrease Cdc6 staining intensity, suggesting that all of the Cdc6 in the nucleus is chromatin bound. The functional significance of this binding has not yet been demonstrated. We looked for evidence that nuclear protein export might regulate Cdc6 levels using leptomycin B, a potent inhibitor of Crm1-dependent export (Nishi et al., 1994). In several immunofluorescence experiments with this drug, we detected no increase in protein in the nucleus (Fig. 1A, +leptomycin B) implying that in late G1, nuclear protein export does not control nuclear Cdc6 levels. However, as a number of soluble export factors can be lost from isolated nuclei (Kehlenbach et al., 1998) it is likely that nuclear protein export does not function optimally in our system. This has allowed identification of another clear level of regulation (see later). Exogenous Cdc6 does not accumulate in the nucleus in S phase We examined the behaviour of exogenous Cdc6 using nuclei and cytosols from different phases of the cell cycle. We used isolated nuclei and the equivalent soluble extracts from quiescent 3T3 cells (G0), 3T3 cells released from quiescence for 16.5 hours (late G1) or 21 hours (S) and from HeLa cells released for 4 hours from a mitotic nocodazole block (early G1), or released for 2 hours (S) or 8 hours (G2) from thymidine arrest. Exogenous Cdc6 protein accumulated in the nucleus in G1 and G0 3T3 nuclei and to a lesser extent in early G1 HeLa nuclei, but failed to accumulate in the nucleus in S phase and G2 (Fig. 1B). This is consistent with reports which look at the localisation of overexpressed Cdc6 in intact cells (Jiang et al., 1999; Petersen et al., 1999; Saha et al., 1998). Absence of Sphase nuclear staining in our experiments suggests that exogenous recombinant Cdc6 is subject to controls similar to overexpressed Cdc6 in intact cells. We next asked why exogenous protein fails to accumulate in S phase. Neither S-phase nuclei nor S-phase cytosol allow nuclear accumulation of Cdc6 We dissected the G1-specific nuclear accumulation of Cdc6 by mixing nuclei and cytosol from different phases of the cell cycle and assaying for the presence of Cdc6 in the nucleus by immunofluorescence (Fig. 2A). G1, S, G2, M and G0 cytosols were assayed for the ability to support accumulation of exogenous Cdc6 in early G1 HeLa cell nuclei. Only G1 and G0 cytosols supported accumulation of Cdc6. These observations show that factors in the soluble fraction from S and to a lesser extent G2 cells can prevent Cdc6 accumulation in G1 nuclei. Fig. 2A also compares the effect of S phase and G1 cytosolic environments on import into G1, S and G2 nuclei. Neither S phase nor G2 nuclei accumulated Cdc6, even in a G1 cytosolic environment. Therefore, contributions of either S/G2 nuclei or S/G2 cytosol prevent accumulation of Cdc6 in the nucleus. Fig. 2B shows a control reaction in which the behaviour of Xenopus Cdc6 is compared to that of Xenopus nucleoplasmin. Incubation with either S phase or G1 cytosol produced the usual S-phase-dependent exclusion of Cdc6 from G1 nuclei while nucleoplasmin efficiently accumulated in both. This shows that 1932 D. Coverley and others Fig. 1. Exogenous Cdc6 accumulates in G1 nuclei but not S-phase nuclei. (A) Recombinant Xenopus Cdc6 accumulates inside intact nuclei from NIH 3T3cells, when incubated in G1 cytosol (Cdc6 only). Accumulation in the nucleus in G1 is completely blocked by inclusion of a dominant negative importin β fragment (+ import inhibitor) but is unaffected by a nuclear protein export inhibitor (+ leptomycin B). Accumulated protein is resistant to extraction with detergent (+ TX100 wash), indicating that imported Cdc6 is bound to structures within the nucleus. (B) Cdc6 accumulates inside intact nuclei from G1 or G0 NIH 3T3 cells or G1 HeLa cells, but not in nuclei from S-phase 3T3 cells or S-phase or G2 HeLa cells, during incubation in matching cytosols. A clear transition is seen between G1 and S phase. Nuclei are stained red with the DNA stain propidium iodide. Cdc6 is detected using a species-specific affinity purified polyclonal antibody raised against recombinant Xenopus cdc6, and a secondary antibody conjugated to FITC, generating a yellow image in positive nuclei when images of the two fluorochromes are merged. S-phase-dependent exclusion from the nucleus does not apply to all proteins, but is selective for Cdc6. S-phase nuclei promote destruction of Cdc6 Recombinant Cdc6 does not accumulate in the nucleus in S phase, so we looked at the total level of Cdc6 protein, which survives under the conditions used in the immunofluorescence experiments shown in Figs 1 and 2. Complete nuclear import reactions containing nuclei, cytosol and 100 ng recombinant Cdc6 were analysed by western blot after separation by SDSPAGE. When incubated with S-phase nuclei and S-phase cytosol the majority of Cdc6 was destroyed after 30 minutes (Fig. 3A, lanes 3 and 5). Cdc6 destruction is absolutely dependent on the presence of nuclei, as incubation with Sphase cytosol alone (lanes 1 and 4) does not affect Cdc6 survival compared to the amount of protein present at the start Fig. 2. S-phase nuclei and Sphase cytosol both block accumulation of Cdc6. (A) Nuclei from synchronized cells were mixed with cytosols from different cell-cycle phases and assayed for their ability to accumulate exogenous Cdc6. S-phase nuclei do not support accumulation in any cytosol and S-phase cytosol does not support accumulation in any nuclei, therefore both S cytosol and S nuclei contain regulators of Cdc6 subcellular distribution. G2 nuclei behaved the same as S nuclei, while G2 cytosol and M-phase cytosol were similar, but slightly less efficient than S-phase cytosol, at blocking accumulation into G1 nuclei. G0 cytosol behaved like G1 cytosol and supported accumulation into G1 nuclei. Nuclei are stained red with propidium iodide; Cdc6 staining as in Fig. 1, generates a yellow image in positive nuclei. (B) Xnucleoplasmin accumulates in the nucleus in S phase. To show that the differing ability to accumulate Cdc6 in the nucleus between G1 and S phase is not a general phenomenon we monitored the behaviour of another protein under identical conditions. FITC-conjugated Xnucleoplasmin accumulated efficiently in G1 nuclei when incubated in both G1 and S-phase cytosols, while in the same experiment Cdc6 did not. Cdc6 is detected using species-specific anti-Cdc6 antibody and FITC-conjugated secondary antibody. Nuclei are not counterstained. Cdc6 in S phase in human cells 1933 Fig. 3. Phosphorylation inside the nucleus in S phase promotes destruction of free Cdc6. (A) Destruction of Cdc6 in S phase is dependent upon the presence of nuclei. Incubation of 100 ng of recombinant Xenopus Cdc6 in S-phase cytosol in the absence of nuclei (lanes 1 and 4) does not result in any loss of protein compared to the amount of Cdc6 present at the beginning of a reaction (lane 2). Inclusion of S-phase nuclei (lanes 3 and 5) or G1 nuclei (lane 6) resulted in substantial loss of exogenous Cdc6 within 30 minutes at 37°C. (B) S-phase nuclei also promote destruction of Cdc6 in G1 cytosol (lane 3), but G1 nuclei do not (lane 2). The slight increase in survival of Cdc6 seen with G1 nuclei compared to the control reactions that contain no nuclei (lanes 1 and 4) is abolished when nuclear import is inhibited (lane 5). This suggests that the G1 nuclear environment can protect Cdc6 from slow degradation by cytosolic extract. (C) The recombinant Cdc6 that survives in S phase fractionates entirely with the nucleus after permeabilization with detergent. (D) Rapid Sphase destruction of Cdc6 (lane 2) is dependent upon nuclear import. Inclusion of importin β dominant-negative mutant protein to an S-phase nuclei/S-phase cytosol incubation (lane 3) resulted in Cdc6 survival, compared to survival in the control reaction. (E) Cdc6 is protected from destruction during incubation with S-phase nuclei/S-phase cytosol by either olomoucine (OM) or MG132. This is consistent with a model in which cdk phosphorylation activates proteosome-mediated destruction of Cdc6. (F) Mutant Xenopus Cdc6 protein, in which the serine residues in all five consensus Cdk phosphorylation sites were mutated to alanines, survives incubation with S-phase nuclei/S-phase cytosol, arguing for direct phosphorylation of Cdc6 as the trigger for destruction. Destruction assays ran for 30 minutes and were stopped by addition of SDS-PAGE loading buffer. Total products of each reaction were analysed by western blot for survival of the exogenous protein using Xenopus Cdc6-specific polyclonal antibody. In C, reactions were first mixed with an equal volume of 0.5% Triton X-100 then separated into soluble and insoluble fractions by centrifugation to assess whether surviving Cdc6 is present in the nucleus or cytosol. of the reaction (lane 2). When incubated in S-phase cytosol, G1 nuclei also promote destruction of Cdc6 (lane 6). Since Sphase cytosol alone does not catalyse destruction of Cdc6 it is the combination of S-cytosol and G1 nuclei that is responsible for loss of exogenous protein in this experiment. This suggests that G1 nuclei can be induced to perform S-phase functions by soluble S-phase factors, and parallels the induction of DNA replication in G1 nuclei by S-phase cytosol (Krude et al., 1997; Stoeber et al., 1998). Furthermore, transient exposure to Sphase cytosol will activate G1 nuclei to perform both Cdc6 destruction and initiation of DNA replication even in G1 cytosol (not shown), suggesting that the switch to S-phase function is irreversible. G1 nuclei do not promote destruction when assayed in G1 cytosol (Fig. 3B). In fact, by monitoring survival of the whole population of exogenous Cdc6 (rather than just the nuclear protein by immunofluorescence), G1 nuclei were found to promote overall survival of Cdc6, as slightly more Cdc6 is observed at the end of an incubation if G1 nuclei are included (compare lane 2 to lanes 1 and 4). During similar reactions containing only cytosol and Cdc6 some loss of protein (91% survival) was observed after 30 minutes while significant loss (56% survival) was not seen until after 60 minutes (not shown). G1 nuclei could protect exogenous Cdc6 from this ‘slow mode’ cytosolic degradation by sequestering protein within the nucleus. To ask whether improved survival is dependent on nuclear import we included importin β dominant-negative mutant protein. When import is blocked, protection by G1 nuclei is abolished (compare lanes 2 and 5). This is consistent with a model in which chromatin binding protects against Cdc6 instability. Similar to the protection by G1 nuclei, a small amount of Cdc6 always survives proteolysis in S-phase nuclei in vitro, even when reactions are incubated for several hours rather than the usual 30 minutes. Furthermore, when increasing numbers of nuclei are added the amount of protein that survives increases (not shown), suggesting that nuclei contain a limited number of binding sites capable of protecting Cdc6. Consistent with this, surviving Cdc6 protein fractionates with the nucleus after permeabilization with detergent (Fig. 3C). Slow degradation in G1 cytosol differs from the rapid destruction catalysed by S-phase nuclei in several ways. Most significantly, when nuclear import is blocked by the mutant importin β protein, S-phase destruction of Cdc6 is reduced (Fig. 3D). The rapid destruction of Cdc6 by S-phase nuclei is in agreement with the absence of immunofluorescence signal in S-phase nuclei (Figs 1 and 2), and demonstrates a role for regulated proteolysis of Cdc6 in the mammalian cell cycle. 1934 D. Coverley and others Other workers have proposed that the lack of immunofluorescence signal in S-phase nuclei is caused by export of Cdc6 from the nucleus, as overexpressed Cdc6 was found to remain in the nucleus (and not accumulate in the cytoplasm) when transfected human tumour cells were treated with leptomycin B and cycloheximide (Jiang et al., 1999). In those experiments cytoplasmic absence could reflect cessation of synthesis of new Cdc6, because of cycloheximide rather than blocked nuclear export. In addition they do not report quantification of the Cdc6 levels remaining in the nucleus and therefore do not rule out a contribution of nuclear proteolysis. Our experiments, which follow the fate of a defined amount of protein, are consistent with these previous reports but reveal an additional level of regulation. S-phase destruction of Cdc6 is regulated by cyclindependent kinases It has been reported that loss of Cdc6 immunofluorescence signal from the nucleus in S-phase mammalian cells is regulated via phosphorylation of conserved N-terminal CDK consensus sites (Jiang et al., 1999; Petersen et al., 1999; Takei et al., 1999). Phosphorylation drives Cdc6 from the nucleus. We asked whether S-phase-specific degradation of Cdc6 is also regulated by cdk-dependent phosphorylation. Cdc6 was incubated with S-phase cytosol and nuclei in the presence of the cdk inhibitor olomoucine or MG132, a peptide inhibitor of the proteosome (Lee and Goldberg, 1998). Destruction of exogenous Cdc6 was efficiently blocked by either drug (Fig. 3E), consistent with a model in which cdk phosphorylation targets Cdc6 for destruction. More conclusively, a nonphosphorylatable mutant version of Xenopus Cdc6 remains unaffected during incubation with S-phase nuclei/S-phase cytosol (Fig. 3F), demonstrating directly that phosphorylation of Cdc6 is an essential regulatory event in the pathway to destruction. We also looked for any indication that S-phase-specific destruction of Cdc6 might be linked to ongoing DNA synthesis or transcription, but we saw no effect on Cdc6 survival when aphidicolin or actinomycin D were included (not shown). Activation of Cdc6 destruction by recombinant cyclin A-cdk2 Cdc6 has been shown conclusively to interact specifically with active cyclin A-cdk2 complexes (Petersen et al., 1999) and to disappear from the nucleus when cyclin A-cdk2 is activated at the start of S phase. We asked whether destruction of Cdc6 could be promoted in G1 cytosol by recombinant cyclin A-cdk2 (Fig. 4A). Compared to the control reactions with no additions, cyclin A-cdk2, but not equivalent amounts of cyclin E-cdk2 or cyclin D-cdk6 activity, caused efficient destruction of recombinant Xenopus Cdc6, in the absence of any nuclei. Importantly, when human Cdc6 was incubated in G1 cytosol, supplemented with cyclin A-cdk2 it behaved in exactly the same way as Xenopus Cdc6 (Fig. 4B), eliminating the possibility that the regulated destruction seen here is specific to the Xenopus protein. Although the in vivo phosphorylation event that marks Cdc6 for destruction is mediated by a nuclear-associated protein kinase, this experiment shows that when cyclin A-cdk2 is added the proteolysis event can occur in the cytosolic fraction. Additional analysis in which Cdc6 was incubated with S-phase Fig. 4. Cyclin A-cdk2 activated destruction of Cdc6. (A) Equivalent amounts of recombinant human cyclin A-cdk2, cyclin E-cdk2 or cyclin D1-cdk6 activity were added to G1 cytosol in the presence of exogenous Cdc6. Cdk2 or cdk6 uncomplexed to cyclin were included as controls. Cyclin A-cdk2 promoted destruction of Cdc6 in the absence of any nuclei. (B) Like Xenopus Cdc6, destruction of human Cdc6 is regulated by cyclin A-cdk2. Cytosols from G1 HeLa cells or G1 Wi38 cells were supplemented with cyclin A-cdk2 as in A. (C) Destruction of Cdc6 by S-phase nuclei/S-phase cytosol is efficiently inhibited by the specific cdk inhibitor p21. Reactions were processed and analysed as for Fig. 3, except that recombinant human Cdc6 was detected with anti-human Cdc6 polyclonal antibody. nuclei in buffer, in the absence of cytosol (not shown), indicated that proteolysis is not dependent on the cytosolic fraction. These observations are consistent with presence of the proteosome in both the nuclear and cytoplasmic compartments (reviewed in Lee and Goldberg, 1998). When S-phase nuclei are incubated in, but then removed from, S-phase cytosol the activating kinase does not leak out (not shown), consistent with the kinase being chromatin bound. The experiments shown here argue that this kinase is chromatin-bound cyclin A-cdk2. Finally, to confirm that destruction of Cdc6 is activated by a cdk2-dependent kinase we have shown that the G1 cyclindependent kinase inhibitor p21 (Furuno et al., 1999) blocks Sphase destruction of Cdc6 (Fig. 4C). Consistent with this, other workers have reported that all cells in an asynchronous Cdc6 in S phase in human cells 1935 Fig. 5. Human Cdc6 protein persists in the nucleus in S phase. (A) HeLa cells arrested in mitosis (M), early G1, S phase (S) or G2 were separated into soluble and nuclear associated fractions by dounce homogenization and centrifugation. 1×105 cell equivalents were loaded for each sample and separated by SDS-PAGE. Human Cdc6 was detected using affinity-purified polyclonal antibody raised against recombinant human Cdc6. Two Cdc6 isoforms are present in S and G2 nuclei. (B) The slower migrating isoform of nuclear Cdc6 is unstable. In a separate experiment nuclei were prepared from S-phase cells at room temperature in the presence and absence of the phosphatase inhibitor okadaic acid. In both preparations the fast migrating form is the most abundent while the slower migrating form is only apparent when prepared in the presence of inhibitor. (C) Modification of exogenous Xenopus Cdc6 protein by cytosolic extracts from synchronised HeLa cells. S- and G2-phase cytosols, but not G1-phase cytosol, modify exogenous Cdc6 to a slower migrating form. This modification is prevented by inclusion of the cdk inhibitor olomoucine. Soluble extract from nocodazole-arrested M-phase cells produced a greater degree of retardation, which may be due to more extensive phosphorylation. (D) HeLa cells arrested at the beginning of S phase by thymidine block were released into medium containing nocodazole. Nuclei were prepared at 0, 3, 6, 9 and 11 hours. Flow cytometry profiles for the different populations are overlayed to demonstrate progression through S phase. By 11 hours approximately 30% of the population was arrested in mitosis; these cells were discarded to produce a late G2 population. (E) Approximately 1×105 nuclei from the 0, 3, 6, 9 and 11 hour populations shown in D were denatured, seperated by SDS-PAGE and blotted for Cdc6 and actin. (F) Cdc6 and actin band intensities were quantified using NIH image software. Cdc6 levels were normalised against actin and plotted (open circles) on the same axes as the percentage of elongating nuclei for each population (closed circles). Nuclei that were actively replicating in vivo at the time of isolation continue to elongate their DNA in vitro, when incubated in a buffered mix of nucleotides and energy regenerating system. This provides an independent measure of progression through S phase. Cdc6 levels fell by half upon entry into S phase and then remained almost constant. By 9 hours S phase was completely finished yet Cdc6 protein levels remained high, until just before mitosis. population that stain positively for nuclear p21 are also positive for epitope-tagged human Cdc6 (Saha et al., 1998). These data suggest that one of the roles of p21 in vivo in G1 phase is to protect Cdc6 from modification and subsequent destruction, so facilitating assembly of replication complexes. Chromatin-bound endogenous Cdc6 persists in S phase We have monitored the level of endogenous human Cdc6 protein in HeLa cell cytosols and nuclei prepared under in vitro replication conditions, using a previously characterised affinity-purified polyclonal antibody raised against recombinant human Cdc6 (Stoeber et al., 1998). We see persistent Cdc6 in S-phase nuclei and also in G2 phase nuclei, which typically include around 30% late S-phase cells (Fig. 5A). Quantification of nuclear Cdc6 band intensity (Fig. 5E) in synchronised populations taken 0, 3, 6, 9 and 11 hours after release from thymidine arrest reveals that Cdc6 levels fall to approximately one half (relative to actin) by 3 hours into S phase, and then remain constant until just before mitosis. At 11 hours after release a large proportion of cells were arrested in mitosis due to the presence of nocodazole. Mitotic cells were removed by vigorous shaking, leaving a late G2 population in which Cdc6 levels had fallen again. Cdc6 band intensities were normalised against actin levels and plotted using arbitrary units on the same axes as the number of elongating nuclei in each population (Fig. 5F). In vitro elongation assays and flow cytometry profiles (Fig. 5D) provide independent measures of progression through S phase, allowing the firm conclusion that at 9 hours after release from thymidine arrest DNA replication has completely finished, yet Cdc6 remains in the nucleus. Consistent with expression of Cdc6 at G1-S (Saha et al., 1998), the level of Cdc6 in post-G1 nuclei is higher than that in early G1 nuclei (Fig. 5A). Our analysis also reveals cell cycle changes in Cdc6 isoforms. We see a single fast migrating form in G1 and two equally abundant isoforms in S phase and G2. The upper form is unstable in isolated S-phase nuclei at room temperature, but recovery is improved in the presence of the phophatase inhibitor okadaic acid, suggesting that the retarded form is phosphorylated (Fig. 5B). Consistent with this instability, human Cdc6 has recently been reported to interact with a regulatory subunit of protein phosphatase 2A (Yan et al., 2000). S-phase phosphorylation of Cdc6 is observed directly when exogenous Xenopus Cdc6 protein is incubated with soluble extracts prepared from synchronised cells, in the absence of nuclei. S cytosol, but not G1 cytosol, modifies Cdc6 so that it migrates more slowly (Fig. 5C). Furthermore, this modification is prevented by olomoucine, suggesting that cdk-dependent phosphorylation causes the reduced mobility. G2 cytosol also generates a reduced mobility form of Cdc6, and M-phase extract causes an even greater degree of retardation, which probably reflects phosphorylation at more of the ten potential phosphorylation sites in Xenopus Cdc6 (Coleman et al., 1996). 1936 D. Coverley and others DISCUSSION Regulated destruction of Cdc6 In human cells the behaviour of Cdc6 is regulated in the cell cycle by several different mechanisms. Cdk consensus sites near the N-terminal nuclear localisation signal (NLS) are believed to influence nuclear protein import (Takei et al., 1999), while inhibition of nuclear protein export suggests that overexpressed Cdc6 migrates from the nucleus in S phase (Jiang et al., 1999). By using a finite amount of exogenous reporter protein we provide evidence for another level of control, regulated proteolysis, which rapidly destroys free Cdc6 in S phase. In yeast, analysis of cdc18/CDC6 mRNA and protein levels has produced a coherent picture of its regulation in the cell cycle. Transcription begins in mitosis and ends in S phase, and protein levels are regulated by proteolysis. Cdc18 protein is targeted for destruction in mitosis by cdc2-mediated phosphorylation and only begins to accumulate when mitotic kinase activity falls. Later, the rise of S-phase cdk activity again phosphorylates cdc18/CDC6, resulting in rapid destruction (Baum et al., 1998; Jallepalli et al., 1997; Nishitani and Nurse, 1995). Phosphorylation of cdc18/CDC6 in S phase seems to be central to the mechanism that restricts DNA replication to one round per cell cycle. Expression of a mutant form which lacks consensus cdk phosphorylation sites or overexpression of the wild-type protein results in unregulated initiation (Baum et al., 1998; Jallepalli et al., 1997; Liang and Stillman, 1997; Nishitani and Nurse, 1995). Our results on regulated proteolysis of Cdc6 are partly similar to the picture that has emerged from studies in yeasts, but instead of seeing degradation of all Cdc6 in S phase, we see degradation of only the free pool. Endogenous, chromatin-bound Cdc6 is conserved but, we believe, modified in a way that some antibodies fail to recognise (see later). Rapid proteolysis of free Cdc6 in S phase is dependent upon import into the nucleus and upon a nuclear-associated, p21sensitive protein kinase. Cyclin A-cdk2 can completely substitute for the nucleus activating Cdc6 destruction in G1 cytosol. This suggests that nuclear cyclin A-cdk2 is the major trigger for Cdc6 destruction in vivo. Cyclin A-cdk2 is known to be activated at G1-S phase (Carbonaro-Hall et al., 1993; Girard et al., 1991; Pagano et al., 1992; Pines and Hunter, 1990) and has been found at sub-nuclear sites of DNA replication (Cardoso et al., 1993). It is possible therefore that destruction of Cdc6 in vivo is initiated in S phase by phosphorylation of Cdc6 at sites of replication. Whether or not phosphorylation of Cdc6 is coupled to replication, the consequence is elimination of excess protein from the nucleus. Our data is consistent with this being a negative regulatory event that contributes to the prevention of re-initiation by preventing Cdc6 from rebinding to chromatin after initiation. Nuclear membrane permeabilization and the stimulation of DNA replication Extensive experimental evidence argues that the nuclear membrane plays an important role in regulating the formation and activation of replication complexes, by partitioning key proteins to the cytoplasm or the nucleus (Blow and Laskey, 1988; Coverley et al., 1993; Hua et al., 1997; Leno et al., 1992; Leno and Munshi, 1994; Madine et al., 1995a; Walter et al., 1998). In one model it serves to concentrate factors that simultaneously block replication complex assembly and activate initiation, while in another model it restricts access of essential initiation factors to chromatin. These models are not mutually exclusive. Recent work with an in vitro mammalian initiating system, from this laboratory, adds to the current body of evidence about the regulatory role of the nuclear membrane. Stimulation of in vitro initiation by recombinant Cdc6 protein was found to be dependent on artificial permeabilization of the nuclear membrane (Stoeber et al., 1998), adding weight to the theory that Cdc6 might be the Xenopus Mcm ‘loading factor’ whose access to the nucleus is restricted when the nuclear membrane is intact (Madine et al., 1995a). Early reports suggested that Cdc6 does not posses an NLS as mutations in the conserved putative bipartite NLS of the human protein resulted in no detectable alteration in nuclear localisation (Saha et al., 1998). This implied that access to the nucleus might occur by association with another protein or experimentally through a permeabilized nuclear membrane. However, recent analysis has shown that the amino acids altered in these experiments overlap with the cyclin binding domain (Petersen et al., 1999). Total deletion of this domain results in increased nuclear localisation of Cdc6 in S phase rather than the reduced nuclear localisation that might be expected from mutation of an NLS. More recently, mutations in another N-terminal conserved putative NLS abolished nuclear localisation of Cdc6 (Takei et al., 1999), and the two serine residues that flank the site were found to be important for regulating its function (Petersen et al., 1999; Takei et al., 1999). We present strong evidence that Cdc6 is actively transported into G1 nuclei, consistent with reports of a functional NLS. This would argue against Cdc6 as the Xenopus ‘loading factor’. Our current observations about regulated proteolysis of Cdc6 in S phase provide a trivial explanation for why nuclear membrane permeabilization might allow Cdc6 to stimulate in vitro replication. Most of the high level of Cdc6 protein used by Stoeber et al. (1998) to supplement S-phase cytosol would have been destroyed upon addition of nuclei. Permeabilization could allow a proportion of the input protein to bind to G1 chromatin and escape immediate destruction. Consistent with this, detergent-resistant Cdc6 was detected only if the nuclear membrane was permeabilized (Stoeber et al., 1998). This explanation is analogous to the proposal that the nuclear membrane serves to generate an intranuclear environment which inhibits replication complex assembly (Hua et al., 1997). However, our work does not exclude the possibility that permeabilization allows additional replication factors to gain access to the nucleus. In fact efficient pre-loading of Cdc6 into intact G1 nuclei in vitro did not increase the number of initiating nuclei, upon transfer to S-phase cytosol (D. C. and S. T., unpublished observations). This suggests that nuclear membrane permeabilization is required for productive binding of Cdc6 in this system. Endogenous human Cdc6 Finally and importantly, this study looks at the status of endogenous human Cdc6 in cytosolic replication type extracts and isolated nuclei from HeLa cells. Consistent with three of the reports in the literature (Fujita et al., 1999; Stoeber et al., Cdc6 in S phase in human cells 1937 1998; Williams et al., 1998), endogenous nuclear Cdc6 persists in S phase. Quantification of protein levels reveals a distinct drop in nuclear Cdc6 after initiation, followed by almost constant levels in S and G2, and a sharp fall just before mitosis. Early analysis of yeast mutants showed that cdc18/Cdc6 is part of the checkpoint control that prevents mitosis from occurring until DNA synthesis is complete (Kelly et al., 1993). A full investigation of the nuclear associated Cdc6 fraction in human G2 nuclei will be of interest with regard to this function. The immunofluorescence experiments, in which overexpressed Cdc6 disappeared from the nucleus in S phase, suggested that Cdc6 is displaced from chromatin in a manner similar to the Mcm proteins, and then exported to the cytoplasm (or degraded) under the control of cyclin-dependent kinases (Jiang et al., 1999; Petersen et al., 1999; Saha et al., 1998; Takei et al., 1999). While we believe that free Cdc6 is regulated in this way, our data clearly show that some Cdc6 remains associated with the nucleus long after replication has finished. After G1, nuclear Cdc6 exists as two distinct isoforms, of which the slow migrating, phosphorylated form is unstable in isolated nuclei. This form is also antigenically different to the faster migrating form as it is not recognised by a second antihuman Cdc6 polyclonal antibody that was raised against amino acids 145-360 (not shown). If, as our data suggests, chromatinbound Cdc6 exists in S phase as the phosphorylated form, its altered antigenicity is one possible reason why some studies fail to detect it. We observed abundant free, undegraded protein in S-phase cytosols, although levels varied between different preparations (not shown). Given the very high capacity of all our S-phase nuclei/S-phase cytosol preparations to degrade exogenous Cdc6, this is slightly surprising. High endogenous Cdc6 levels might arise in S-phase cells as a consequence of the synchronisation procedure, as suggested previously (Petersen et al., 1999). Our cells were arrested and held for two consecutive cell cycles at the beginning of S phase, the point at which mRNA levels are known to peak in the unaltered cell cycle (Saha et al., 1998). This could produce unphysiological Cdc6 levels which, our data shows, could only be degraded after nuclear protein import and phosphorylation inside the nucleus. Our aim has been to better understand the regulation of Cdc6 as a step toward a broader goal of dissecting the mammalian cellular initiation process. In this study we present strong evidence that human Cdc6 persists in the nucleus beyond S phase. In addition we have analysed the regulatory forces acting upon free Cdc6 at the transition from G1 to S phase. Exogenous Cdc6 is eliminated from the nucleus at the start of S phase, by cyclin A-cdk2 regulated proteolysis. This mechanism could function in vivo to remove any Cdc6 not assembled into replication complexes, as well as the Cdc6, which is lost from chromatin after initiation. This could contribute to the prevention of reinitiation by blocking formation of new replication complexes. We are pleased to thank the following people for kindly providing reagents: P. Romanowski for anti-human Cdc6 antibody, K. Stoeber and G. Williams for isoform-specific anti-human Cdc6 polyclonal antibody, H. Laman for cyclin D-cdk6 and cyclin E-cdk2, W. Krek and C. Wirbelauer for cyclin A-cdk2, D. Gorlich’s laboratory for purified importin β dominant-negative mutant, Nobuo Furuno for purified p21 protein, Minoru Yoshida for leptomycin B, and P. Carpenter, T. Coleman, W. Dunphy for the baculovirus Xcdc6 expression construct. We are also grateful to M. Jackman for helpful discussions. This work was supported by the Cancer Research Campaign except for C.P., who was supported by a post-doctoral Human Frontiers Science Program fellowship. REFERENCES Baum, B., Nishitani, H., Yanow, S. and Nurse, P. (1998). Cdc18 transcription and proteolysis couple S phase to passage through mitosis. EMBO J. 17, 5689-5698. Blow, J. J. and Laskey, R. A. (1988). A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature 332, 546-548. Botchan, M. (1996). Coordinating DNA replication with cell division: Current status of the licensing concept. Proc. Natl. Acad. Sci. USA 93, 9997-10000. Carbonaro-Hall, D., Williams, R., Wu, L., Warburton, D., ZeichnerDavid, M., MacDougall, M., Tolo, M. and Hall, F. (1993). G1 expression and multistage dynamics of cyclin A in human osteosarcoma cells. Oncogene 8, 1649-1659. Cardoso, M. C., Leonhardt, H. and Nadal-Ginard, B. (1993). Reversal of terminal differentiation and control of DNA replication: Cyclin A and cdk2 specifically localise at subnuclear sites of DNA replication. Cell 74, 1-20. Carpenter, P. B., Mueller, P. R. and Dunphy, W. G. (1996). Role for a Xenopus Orc2-related protein in controlling DNA replication. Nature 379, 357-360. Chong, J. P. J., Mahbubani, H. M., Khoo, C.-Y. and Blow, J. J. (1995). Purification of an MCM-containing complex as a component of the replication licensing system. Nature 375, 418-421. Cocker, J. H., Piatti, S., Santocanale, C., Nasmyth, K. and Diffley, J. F. X. (1996). An essential role for the Cdc6 protein in forming the pre-replicative complexes of budding yeast. Nature 379, 180-182. Coleman, T. R., Carpenter, P. B. and Dunphy, W. G. (1996). The Xenopus Cdc6 protein is essential for the initiation of a single round of DNA replication in cell-free extracts. Cell 87, 53-56. Coverley, D., Downes, C. S., Romanowski, P. and Laskey, R. A. (1993). Reversible effects of nuclear membrane permeabilization: evidence for a positive licensing factor. J. Cell Biol. 122, 985-992. Coverley, D., Wilkinson, H. R., Madine, M. A., Mills, A. D. and Laskey, R. A. (1998). Protein kinase inhibition in G2 causes mammalian Mcm proteins to reassociate with chromatin and restores ability to replicate. Exp. Cell Res. 238, 63-69. Dahmann, C., Diffley, J. F. X. and Nasmyth, K. A. (1995). S-phase promoting cyclin dependent kinases prevent re-replication by inhibiting the transition of replication origins to a pre-replicative state. Curr. Biol. 5, 12571269. Diffley, J. F. X. (1996). Once and only once upon a time: specifying and regulating origins of DNA replication in eukaryotic cells. Genes Dev. 10, 2819-2830. Donovan, S., Harwood, J., Drury, L. S. and Diffley, J. F. (1997). Cdc6dependent loading of Mcm proteins onto pre-replicative chromatin in budding yeast. Proc. Natl. Acad. Sci. USA 94, 5611-5616. Fujita, M., Yamada, C., Goto, H., Yokayama, N., Kuzushima, K., Inagaki, M. and Tsurumi, T. (1999). Cell cycle regulation of human Cdc6 protein. J. Biol. Chem. 274, 25927-25932. Furuno, N., den Elzen, N. and Pines, J. (2000). Human cyclin A is required for mitosis until mid prophase. J. Cell Biol., in press. Girard, F., Strausfeld, U., Fernandez, A. and Lamb, N. J. (1991). Cyclin A is required for the onset of DNA replication in mammalian fibroblasts. Cell 67, 1169-1179. Harlow, E. and Lane, D. (1988). Antibodies: A Laboratory Manual. New York: Cold Spring Harbour Laboratory Press. Hateboer, G., Wobst, A., Petersen, B. O., Le Cam, L., Vigo, E., Sardet, C. and Helin, K. (1998). Cell cycle-regulated expression of mammalian CDC6 is dependent on E2F. Mol. Cell. Biol. 18, 6679-6697. Hua, X. H., Yan, H. and Newport, J. (1997). A role for Cdk2 kinase in negatively regulating DNA replication during S phase of the cell cycle. J. Cell Biol. 137, 183-192. Jallepalli, P. V., Brown, G. W., Muzi-Falconi, M., Tien, D. and Kelly, T. J. (1997). Regulation of the replication initiator protein p65 cdc18 by CDK phosphorylation. Genes Dev. 11, 2767-2779. 1938 D. Coverley and others Jiang, W., Wells, N. J. and Hunter, T. (1999). Multistep regulation of DNA replication by Cdk phosphorylation of HsCdc6. Proc. Natl. Acad. Sci. USA 96, 6193-6198. Kehlenbach, R. H., Dickmanns, A. and Gerace, L. (1998). Nucleocytoplasmic shuttling factors including Ran and CRM1 mediate nuclear export of NFAT in vitro. J. Cell Biol. 141, 863-874. Kelly, T. J., Martin, G. S., Forsburg, S. L., Stephen, R. J., Russo, A. and Nurse, P. (1993). The fission yeast cdc18 gene product couples S phase to START and mitosis. Cell 74, 371-382. Krude, T., Jackman, M., Pines, J. and Laskey, R. A. (1997). Cyclin/Cdkdependent initiation of DNA replication in a human cell-free system. Cell 88, 109-119. Krude, T., Musahl, C., Laskey, R. A. and Knippers, R. (1996). Human replication proteins hCdc21, hCdc46 and P1Mcm3 bind chromatin uniformly before S-phase and are displaced locally during DNA replication. J. Cell Sci. 109, 309-318. Kubota, K., Mimura, S., Nishimoto, S.-I., Takisawa, H. and Nojima, H. (1995). Identification of the yeast MCM-related protein as a component of Xenopus DNA replication licensing factor. Cell 81, 601-609. Kunagai, A. and Dunphy, W. G. (1995). Control of the Cdc2-cyclin B complex in Xenopus egg extracts arrested at the G2/M checkpoint with DNA synthesis inhibitors. Mol. Biol. Cell 6, 199-213. Kutay, U., Izaurralde, E., Bischoff, F. R., Mattaj, I. W. and Gorlich, D. (1997). Dominant-negative mutants of Importin-β block multiple pathways of import and export through the nuclear pore complex. EMBO J. 16, 11531163. Lee, D. H. and Goldberg, A. L. (1998). Proteosome inhibitors: valuable tools for cell biologists. Trends Cell Biol. 8, 397-403. Leno, G. H., Downes, C. S. and Laskey, R. A. (1992). The nuclear membrane prevents replication of human G2 nuclei but not G1 nuclei in Xenopus egg extract. Cell 69, 151-158. Leno, G. H. and Munshi, R. (1994). Initiation of DNA replication in nuclei from quiescent cells requires permeabilization of the nuclear membrane. J. Cell Biol. 127, 5-14. Liang, C. and Stillman, B. (1997). Persistent initiation of DNA replication and chromatin-bound MCM proteins during the cell cycle in cdc6 mutants. Genes Dev. 11, 3375-3386. Madine, M., Khoo, C.-Y., Mills, A. D., Musahl, C. and Laskey, R. A. (1995a). The nuclear envelope prevents reinitiation of replication by regulating the binding of MCM3 to chromatin in Xenopus egg extracts. Curr. Biol. 5, 1270. Madine, M., Khoo, C.-Y., Mills, T. and Laskey, R. A. (1995b). MCM3 complex is required for cell cycle regulation of DNA replication in vertebrate cells. Nature 375, 421-424. Mills, A. D., Blow, J. J., White, J. G., Amos, W. B., Wilcock, D. and Laskey, R. A. (1989). Replication occurs at discrete foci spaced throughout nuclei replicating in vitro. J. Cell Sci. 94, 471-477. Muzi-Falconi, M., Brown, G. W. and Kelly, T. J. (1996). DNA replication: Controlling initiation during the cell cycle. Curr. Biol. 6, 229-233. Newlon, C. S. (1997). Putting it all together: building a prereplicative complex. Cell 91, 717-720. Nishi, K., Yoshida, M., Fujiwara, D., Nishikawa, M., Horinouchi, s. and Beppu, T. (1994). Leptomycin B targets a regulatory cascade of crm1, a fission yeast nuclear protein, involved in control of higher order chromosom structure and gene expression. J. Biol. Chem. 269, 6320-6324. Nishitani, H. and Nurse, P. (1995). p65cdc18 plays a major role controlling the initiation of DNA replication in yeast. Cell 83, 397-405. Pagano, M., Pepperkok, R., Verde, F., Ansorage, W. and Dreatta, G. (1992). Cyclin A is required at two points in the human cell cycle. EMBO J. 11, 961-971. Petersen, B. O., Lukas, J., Sorensen, C. S., Bartek, J. and Helin, K. (1999). Phosphorylation of mammalian CDC6 by Cyclin A/CDK2 regulates its subcellular localization. EMBO J. 18, 396-410. Piatti, S., Bohm, T., Cocker, J. H., Diffley, J. F. X. and Nasmyth, K. (1996). Activation of S-phase-promoting CDKs in late G1 defines a ‘point of no return’ after which Cdc6 synthesis cannot promote DNA replication in yeast. Genes Dev. 10, 1516-1531. Piatti, S., Lengauer, C. and Nasmyth, K. (1995). Cdc6 is an unstable protein whose de novo synthesis in G1 is important for the onset of S-phase and for preventing a ‘reductional’ anaphase in the budding yeast Saccaromyces cerevisiae. EMBO J. 14, 3788-3799. Pines, J. and Hunter, T. (1990). Human cyclin A is adenovirus E1Aassociated protein p60 and behaves differently from cyclin B. Nature 346, 760-763. Romanowski, P. and Madine, M. A. (1996). Mechanisms restricting DNA replication to once per cell cycle: MCMs, pre-replicative complexes and kinases. Curr. Biol. 6, 184-188. Romanowski, P. and Madine, M. A. (1997). Mechanisms restricting DNA replication to once per cell cycle: the role of Cdc6 and ORC. Curr. Biol. 7, 9-10. Romanowski, P., Madine, M. A., Rowles, A., Blow, J. J. and Laskey, R. A. (1996). The Xenopus origin recognition complex is essential for DNA replication and MCM binding to chromatin. Curr. Biol. 6, 1416-1425. Rowles, A. and Blow, J. J. (1997). Chromatin proteins involved in the initiation of DNA replication. Curr. Opin. Genet. Dev. 7, 152-157. Rowles, A., Chong, J. P. J., Howell, M., Evan, G. I. and Blow, J. J. (1996). Interactions between the origin recognition complex and the replication licensing system in Xenopus. Cell 87, 287-296. Saha, P., Chen, J., Thome, K. C., Lawlis, S. J., Hou, Z., Hendricks, M., Parvin, J. D. and Dutta, A. (1998). Human CDC6/Cdc18 Associates with Orc1 and Cyclin-cdk and Is Selectively Eliminated from the Nucleus at the Onset of S Phase. Mol. Cell Biol. 18, 2758-2767. Stillman, B. (1996). Cell Cycle Control of DNA Replication. Science 274, 1659-1664. Stoeber, K., Mills, A. D., Kubota, Y., Krude, T., Romanowski, P., Marheineke, K., Laskey, R. A. and Williams, G. H. (1998). Cdc6 protein causes premature entry into S phase in a mammalian cell-free system. EMBO J. 17, 7219-7229. Takei, Y., Yamamoto, K. and Tsujimoto, G. (1999). Identification of the sequence responsible for the nuclear localisation of human Cdc6. FEBS lett. 447, 292-296. Tanaka, T., Knapp, D. and Nasmyth, K. (1997). Loading of an Mcm Protein onto DNA Replication Origins is Regulated by Cdc6p and CDKs. Cell 90, 649-660. Todorov, I. T., Attaran, A. and Kearsey, S. E. (1995). BM28, a human member of the MCM2-3-5 family, is displaced from chromatin during replictaion. J. Cell Biol. 129, 1433-1455. Walter, J., Sun, L. and Newport, J. (1998). Regulated Chromosomal DNA Replication in the Absence of a Nucleus. Mol. Cell 1, 519-529. Williams, G. H., Romanowski, P., Morris, L., Madine, M., Mills, A. D., Stoeber, K., Marr, J., Laskey, R. A. and Coleman, N. (1998). Improved cervical smear assessment using antibodies against proteins that regulate DNA replication. Proc. Natl. Acad. Sci. USA 95, 14932-14937. Williams, R. S., Shohet, R. V. and Stillman, B. (1997). A human protein related to yeast Cdc6. Proc. Natl. Acad. Sci. USA 94, 142-147. Wuarin, J. and Nurse, P. (1996). Regulating S Phase: CDKs, Licensing and Proteolysis. Cell 85, 785-787. Yan, Z., DeGregori, J., Shohet, R., Leone, G., Stillman, B., Nevins, J. R. and Sanders Williams, R. (1998). Cdc6 is regulated by E2F and is essential for DNA replication in mammalian cells. Proc. Natl. Acad. SCi. USA 95, 3603-3608. Yan, Z., Fedorov, S. A., Mumby, M. C. and Williams, R. S. (2000). PR48, a novel regulatory subunit of protein phosphatase 2A, interacts with cdc6 and modulates DNA replication in human cells. Mol. Cell Biol. 20, 10211029. Zhou, C., Huang, S. H. and Y., J. A. (1989). Molecular cloning of Saccaromyces cerevisiae CDC6 gene, identification and sequence analysis. J. Biol. Chem. 264, 9022-9029.