Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Point mutation wikipedia , lookup
Paracrine signalling wikipedia , lookup
Signal transduction wikipedia , lookup
Ancestral sequence reconstruction wikipedia , lookup
Gene expression wikipedia , lookup
Expression vector wikipedia , lookup
Magnesium transporter wikipedia , lookup
Biochemistry wikipedia , lookup
Interactome wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Metalloprotein wikipedia , lookup
Protein purification wikipedia , lookup
Structural alignment wikipedia , lookup
Homology modeling wikipedia , lookup
Western blot wikipedia , lookup
Two-hybrid screening wikipedia , lookup
Protein–protein interaction wikipedia , lookup
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
Lecture 6: Proteins: Tertiary and Quaternary Structure [PDF]
Reading: Berg, Tymoczko & Stryer, 6th ed., Chapter 2, pp. 44-53, pp. 61-62
Updated on: 1/23/07 at 5:00 pm
Key Concepts
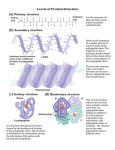
Tertiary and quaternary structures result from folding of primary structure (and secondary structural elements) in 3
dimensions.
Tertiary structure
Most proteins' 3 o structures are combinations of α helices, β sheets, and loops and turns.
Larger proteins often have multiple folding domains.
Folding of H2O-soluble, globular proteins into their native structures follows some basic rules/principles:
minimization of solvent-accessible surface area (burying hydrophobic groups)
maximization of intraprotein hydrogen bonds
chirality (right-handed twist and connectivity) of the polypeptide backbone
Quaternary Structure
Some proteins have multiple polypeptide chains (quaternary structure).
Arrangement of polypeptides in multimeric protein is generally symmetrical.
Quaternary structure can play important functional roles for multi-subunit proteins.
Objectives
Outline 3 principles guiding folding of water-soluble globular proteins and the generalizations about protein
structure resulting from those principles. Relate principles to real protein structures. Define the secondary
structure of a protein, and list examples of categories of secondary structure that occur in proteins.
Explain the term amphipathic, with an amphipathic protein a helix as an example.
Explain the terms supersecondary structure and tertiary structure. Recognize examples (ribbon diagrams) of
such supersecondary structures as coiled coils of α-helices, stacked β-sheets, βαβ elements, β-barrels, and β
saddles.
Define the terms domain and subunit as they relate to protein structure. Be able to recognize different domains
in a ribbon diagram of a single polypeptide chain with 2 or more domains.
Describe in general terms the structure of the polypeptide chain of myoglobin.
Describe the structure of the structure called the “immunoglobulin fold” (single domain).
Describe the general structure of an αβ barrel, including where in the structure you would expect to find
hydrophobic groups and where you would expect to find polar/charged groups.
Describe the general structure (arrangement of hydrophobic vs. polar R groups) of a globular protein that is
embedded in a lipid bilayer (membrane). Specifically, describe how the primary and secondary structures of a
bacterial porin relate to the tertiary structure (and function) of a single porin subunit.
Explain the term quaternary structure of a protein, and be able to describe a protein in terms like
"homotetramer", "heterodimer", etc.
Explain simple rotational symmetry for an oligomeric protein such as a homodimer like the Cro protein or a
heterotetramer like hemoglobin. Be able to use (correctly!) the terms "2-fold", "3-fold", etc. to refer to simple
rotational axes of symmetry and recognize that simple level of symmetry in a protein structure.
Tertiary Structure: 3-dimensional conformation of a whole polypeptide chain in its folded state (includes not only
positions of backbone atoms, but of all the sidechain atoms as well)
Most water-soluble and membrane proteins are globular: compact and roughly spherical
Secondary structures are further folded into compact tertiary structures.
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 1 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
From x-ray diffraction studies and solution nmr studies of proteins, we know the details of the three dimensional
structures of thousands of globular proteins.
Every protein has a unique three dimensional structure made up of a variety of helices, beta-sheets and non-regular
regions, which are folded in a specific manner.
Combinations (patterns) of secondary structural elements called motifs are often found in proteins. Some motifs have
functional significance, such as the helix-loop-helix (helix-turn-helix) DNA-binding motif or the EF hand calciumbinding motif. Others serve only a structural role.
Jmol routine: some structural motifs found in proteins (motif).
3 principles guiding folding of H2O-soluble, globular proteins & consequences of those principles -Generalizations about H2O-soluble, globular protein structure)
1 . *minimization of solvent-accessible surface area
2 . maximization of intra-protein hydrogen bonds
3 . chiral effect
*minimization of solvent-accessible surface area
burying as many hydrophobic groups as possible
Maximizing "hydrophobic interactions" really means getting hydrophobic groups OUT OF WATER.
probably the most important driving force in folding of water-soluble proteins
Hydrophobic R groups mostly cluster in interior, BUT
there can still be some hydrophobic groups in contact with H2O.
Globular protein structures generally tightly packed, compact units
Secondary structural elements (α-helices and β sheets) often amphipathic
R groups on one side hydrophobic (and face interior of protein)
R groups on other side hydrophilic (and face aqueous environment, outside)
Jmol routine showing locations of hydrophobic and hydrophilic side chains
Burial of hydrophobic R groups requires at least 2 interacting
secondary structural elements, e.g., 2 α helices, or a β-α-β loop
(uses α helix to connect 2 parallel β strands), or 2 β sheets, etc.
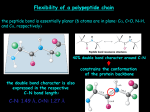
Berg, Tymoczko & Stryer, 6th ed. Fig. 2.44
How can 2 α helices get together to bury hydrophobic R groups,
if there's water around them?
amphipathic helices -- used to bury hydrophobic R groups
toward interior of protein on 1 side of helix while other side of
helix interacts with H2O
Fig. 16-30 from Stryer, Biochemistry, 4th ed.(1995) α-helical coiled coil (2 α helices coiled around each other) of a leucine zipper
motif (heptad repeat)
schematic diagram ("helical wheels" projections down helix axes)
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 2 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
Residues a and d of each strand
pack tightly together to form a
hydrophobic core.
If residues b, c, and f on periphery
are polar or charged, the helices are
amphipathic helices.
Note: Any protein α-helix will be amphipathic if one side of the helix is in a polar environment and the other side is in a
hydrophobic environment.
maximization of intra-protein hydrogen bonds
permits "burying" polar N-H and C=O groups of backbone
also sometimes burying polar side chains, if their polar groups are hydrogen-bonded
especially important in "driving"/stabilizing formation of secondary structures like α-helices and β sheets
makes burying polar protein backbone in interior of protein more favorable
polar backbone groups & side chains tend to be either
in contact with water (hydration) OR
hydrogen-bonded with OTHER PROTEIN GROUPS (e.g., in secondary structures like α-helices and β
sheets)
chiral effect
tendency of extended backbone structural arrangements to be right-handed as a result of having all L-amino acids
Consequences: twist & connectivity
twist:
α helices of L-amino acids tend to be right-handed.
β-conformation strands (and sheets) of L-amino acids tend to twist in a right-handed direction,
forming saddles or barrels.
connectivity: crossovers between adjacent secondary structural elements, e.g., in βαβ structure, are usually righthanded.
SUPERSECONDARY structural motifs (groupings of secondary structural elements)
bury some R groups in between
examples:
coiled coils of 2 or more α helices (αα)
stacks of β-sheets
βαβ elements
β barrels (β sheet folds/twists into a cylinder)
β saddles (twisted β sheet), etc.
DOMAINS: structurally independent folding units looking like separate globular proteins but all part of same polypeptide chain
(connected in same primary structure)
Larger proteins often have 2 or more domains.
Examples of tertiary structures of water-soluble globular proteins:
1. Myoglobin (Jmole structure of myoglobin)
binds O2 in muscle cells for storage and for intracellular transport, using a heme group (black in structure below,
with purple Fe2+)
mostly (70%) α-helical; rest mostly turns & loops (at surface)
first high-resolution crystal structure of a protein ever determined
very compact structure (almost no empty space inside)
very water-soluble
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 3 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
5 Pro residues, all in turns
8 α helices, designated by letters A - H, from N to C terminus
Berg, Tymoczko & Stryer, 6th ed. Fig. 2.48: Myoglobin (Mb) tertiary structure (A: ball & stick; B: ribbon diagram)
Mb binds O2 in muscle cells for storage and for intracellular transport, using a heme group (in black, with purple
Fe2+)
Berg, Tymoczko & Stryer Fig. 2.49: Distribution of Amino Acids in Myoglobin structure
(hydrophobic residues in yellow, charged residues in blue, others in white)
A. Surface view ; B. cross-sectional view showing interior of protein
NOTE: many charged residues on surface, none in interior
many hydrophobic residues in interior, but also a few on surface
The only polar residues inside are 2 His residues involved in binding the heme and O2.
2. Triose phosphate isomerase (an enzyme in the glycolytic pathway) Jmol structures of αβ proteins
Garrett & Grisham, Biochemistry, 2nd ed., Fig. 6-30: Some βαβ enzymes (triose phosphate isomerase, and one domain of
pyruvate kinase)
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 4 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
(a more detailed look at the center panel in figure
above)
Berg, Tymoczko & Stryer, 6th ed. Fig. 16.4, p. 439:
Triose phosphate isomerase, an αβ barrel (parallel
8-stranded β barrel on interior, surrounded by α
helices, a structural motif found in many different
enzymes)
3. Protein domains:
structurally independent folding units looking like separate globular proteins
but connected in same primary structure (part of a single polypeptide chain)
(Jmol structure of a large, multi-domain protein)
Nelson & Cox, Lehninger Principles of Biochemistry,
4th ed., 2004, Fig. 4-19: Structural domains in the
single polypeptide chain Troponin C
Berg, Tymoczko & Stryer, 6th ed. Fig. 2.52: a cell surface protein (CD4), with 4 similar domains (each in a different
color). The folding motif of each of the 4 domains is the same. (Each domain consists of 2 antiparallel β sheets, with
loops between β strands, a motif called the "immunoglobulin fold".)
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 5 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
4. Porins (one type of membrane protein structure)
found in outer membranes of many bacteria and in outer mitochondrial membranes
channel-forming proteins permitting passage of ions and small molecules across the membrane
globular, but their "solvent" is NOT water -- it’s a membrane
core of membrane like a very nonpolar solvent
structure of each chain of a porin is mainly a large β barrel (big antiparallel β sheet, 16 strands, folded
into a cylinder)
structure is sort of like "inside out" water-soluble protein
hydrophobic residues on outer surface, interacting with the hydrophobic lipid core of the membrane
inner side of the barrel has more hydrophilic (charged and polar) R groups (forming water-filled
channel across membrane)
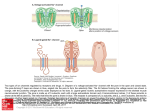
Berg, Tymoczko & Stryer, 6th ed. Fig. 2.50: Amino acid
distribution in structure of a bacterial porin (yellow =
hydrophobic residues, blue = charged/polar residues) -"inside out" compared to a water-soluble globular protein
See also Jmol routine: one subunit of a bacterial porin
Berg, Tymoczko & Stryer, Fig. 12.20, p. 338: Structure of one subunit of a bacterial porin (A: side view, in plane of
membrane; B: view from periplasmic space (from inside, looking out through pore in outer membrane)
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 6 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
Berg, Tymoczko & Stryer Fig. 12.21, p. 338: Amino acid sequence of a porin.
β strands are indicated, with diagonal lines indicating direction of hydrogen bonding along the β sheet
hydrophobic residues (F, I, L, M, V, W and Y) shown in yellow
Note the more or less alternating hydrophobic and hydrophilic residues in the β strands (adjacent R groups project
out from sheet on opposite sides).
Quaternary structure (4° structure):
3-dimensional relationship of the different polypeptide chains (subunits) in a multimeric protein, the way the subunits fit
together and their symmetry relationships (only in proteins with more than one polypeptide chain; proteins with only one chain
have no quaternary structure.)
Terminology:
Each polypeptide chain in a multichain protein = a subunit
2-subunit protein = a dimer, 3 subunits = trimeric protein, 4 = tetrameric, etc.
homo(dimer or trimer etc.): identical subunits
hetero(dimer or trimer etc.): more than one kind of subunit (chains with different amino acid sequences)
different subunits designated with Greek letters
e.g., subunits of a heterodimeric protein = the "α subunit" and the "β subunit".
NOTE: This use of the Greek letters to differentiate different polypeptide chains in a multimeric protein has
nothing to do with the names for the secondary structures α helix and β conformation.
Some protein structures have very complex quaternary structures; e.g., mitochondrial ATP synthase, and (even more complex) viral
capsids.
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 7 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
Examples of protein quaternary structure
Berg, Tymoczko & Stryer Fig. 2.53: The Cro protein of bacteriophage l , a homodimer (2 identical subunits)
Berg, Tymoczko & Stryer, 6th ed. Fig. 2.54: Hemoglobin, a heterotetramer (quaternary structure α 2β 2).
structure of the 2 identical α subunits (red) very similar to structure of the 2 identical β subunits (yellow)
both structures also very similar to
structure of the myoglobin polypeptide
chain (both primary and tertiary structure)
gene duplication of single ancestral gene
and subsequent divergent evolution of
sequences --> different globin genes
overall tertiary "fold" conserved
through evolution
Jmol structure of hemoglobin
Symmetry in quaternary structures
simplest kind of symmetry = rotational symmetry
Individual subunits can be superimposed on other identical subunits (brought into coincidence) by rotation
about one or more rotational axes.
If the required rotation = 180° (360°/2), protein has a 2-fold axis of symmetry (e.g., Cro repressor protein
above).
If the rotation = 120° (360°/3), e.g., for a homotrimer, the protein has a 3-fold symmetry axis.
Nelson & Cox, Lehninger Principles of Biochemistry, 4th ed. (2004), Fig. 4-24a: Rotational symmetry in
proteins. Cyclic symmetry: all subunits are related by rotation about a single n-fold rotation axis (C2 symmetry
has a 2-fold axis, 2 identical subunits; C 3 symmetry has a 3-fold axis, 3 identical subunits, etc.)
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 8 of 9
LEC6_ProteinTertQuatStruct
01/23/2007 05:11 PM
What type of rotational axis of symmetry is apparent in the HEMOGLOBIN structure above?
Why would a protein "want" to HAVE a quaternary structure (subunits)?
ease of folding of small vs. large polypeptide chains
signaling/communication ("division of labor", regulatory functions...)
use of same subunit in different multi-subunit proteins/complexes
[email protected] .edu
Department of Biochemistry & Molecular Biophysics
The University of Arizona
Copyright (©) 2007
All rights reserved.
http://www.biochem.arizona.edu/classes/bioc460/spring/460web/lectures/LEC6_ProteinTertQuatStruct/LEC6_ProteinTertQuatStr.html
Page 9 of 9