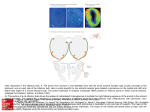
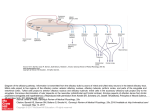
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Axon guidance wikipedia , lookup
Development of the nervous system wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Subventricular zone wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Synaptogenesis wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Sensory cue wikipedia , lookup
Signal transduction wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Optogenetics wikipedia , lookup
UMEÅ UNIVERSITY MEDICAL DISSERTATIONS New series No. 1047 ISSN 0346-6612 Molecular and functional anatomy of the mouse olfactory epithelium Viktoria Vedin Department of Molecular Biology Umeå University Umeå 2006 Department of Molecular Biology Umeå University S-901 87 Umeå, Sweden Copyright © 2006 by Viktoria Vedin ISBN 91-7264-138-X Printed by Solfjädern Offset AB, Umeå, Sweden, 2006 2 "Det är en god vana att då och då ifrågasätta saker och ting som man i åratal har tagit för givna." Tyskt ordspråk 3 4 TABLE OF CONTENTS ABSTRACT ........................................................................................................... 6 PAPERS IN THIS THESIS ................................................................................... 7 ABBREVIATIONS................................................................................................ 8 INTRODUCTION ................................................................................................ 9 Cell types of the olfactory epithelium ........................................................... 10 Proliferative capacity of the olfactory epithelium........................................ 12 Odorant receptors ........................................................................................... 14 Signal transduction in olfactory sensory neurons ........................................ 15 Odorant receptor expression patterns ........................................................... 18 Projections to the brain .................................................................................. 21 Initial steps in constructing a smell in the brain .......................................... 24 AIM OF THE THESIS ........................................................................................ 28 METHODS.......................................................................................................... 29 Studying odorant detection threshold........................................................... 29 RESULTS AND DISCUSSION ........................................................................... 33 Gene expression correlated to the zonal topography of ORs (paper I) ....... 33 Regionally restricted detection of odorants (papers II and III) ................... 37 Odorants induce c-Fos in OSNs in an OR-like pattern ............................ 37 Zonal ablation and effects on odorant detection....................................... 39 Regional differences in maintenance of the olfactory epithelium (papers III and IV) ....................................................................... 43 The olfactory epithelium can be divided into two major domains ............. 48 CONCLUSIONS.................................................................................................. 50 ACKNOWLEDGEMENTS ................................................................................. 51 REFERENSES...................................................................................................... 52 5 ABSTRACT The olfactory system is important for social behaviors, feeding and avoiding predators. Detection of odorous molecules is made by odorant receptors on specialized sensory neurons in the olfactory epithelial sheet. The olfactory sensory neurons are organized into a few regions or “zones” based on the spatially limited expression of odorant receptors. In this thesis the zonal division and functional specificity of olfactory sensory neurons have been studied in the mouse. We find that zones 2-4 show overlapping expression of odorant receptors while the border between the regions that express a zone 1 and a zone 2 odorant receptor, respectively, is sharp. This result indicates that zone 1 and zones 2-4 are inherently different from each other. In cDNA screens, aimed at finding genes whose expression correlate to the zonal expression pattern of odorant receptors, we have identified a number of signaling proteins implicated in neural-tissue organogenesis in other systems. The differential expression pattern of identified genes suggests that regional organization is maintained during the continuous neurogenesis in the olfactory epithelium as a result of counter gradients of positional information. We show that the gene c-fos is induced in olfactory sensory neurons as a result of cell activation by odorant exposure. A zonal and scattered distribution of c-Fos-positive neurons resembled the pattern of odorant receptor expression and a change of odorant results in a switch in which zone that is activated. Whereas earlier studies suggest that the odorant receptors are relatively broadly tuned with regard to ligand specificity, the restricted patterns of c-Fos induction suggests that low concentrations of odorous molecules activate only one or a few ORs. Studies on olfactory detection abilities of mice with zonal-restricted lesions in the olfactory epithelium show that loss of a zone has severe effects on the detection of some odorants but not others. These findings lend support to a hypothesis that odorant receptors are tuned to more limited numbers of odorants. Regional differences in gene expression and differences in response to toxic compounds between the zones indicate that there may be differences in tissue homeostasis within the epithelium. We have found that there are differences in proliferation and survival of olfactory sensory neurons in regions correlating to receptor expression zones. Identified differences with regard to gene expression, tissue homeostasis and odorant detection show that the olfactory epithelium is divided into regions that transduce different stimulus features. 6 PAPERS IN THIS THESIS The thesis is based on following articles and manuscripts, which will be referred to in the text by their Roman numerals (I-IV). I. Norlin EM, Alenius M, Gussing F, Hägglund M, Vedin V, Bohm S, (2001). Evidence for gradients of gene expression correlating with zonal topography of the olfactory sensory map. Mol Cell Neurosci. 18:283-295. II. Norlin EM, Vedin V, Bohm S, Berghard A, (2005). Odorantdependent, spatially restricted induction of c-fos in the olfactory epithelium of the mouse. J Neurochem. 93:1594-1602. III. Vedin V, Slotnick B, Berghard A, (2004). Zonal ablation of the olfactory sensory neuroepithelium of the mouse: effects on odorant detection. Eur J Neurosci. 20:1858-1864 IV. Vedin V, Berghard A, (2006). Regional differences in turnover of olfactory sensory neurons in the adult. Manuscript. Articles are reprinted with permission from the publishers. 7 ABBREVIATIONS ACIII AMP ATP CaM CAMKII cAMP cGMP CNG channel Dichlobenil GBC HBC Mash1 MRR NQO1 O/E OMP OR OSN PDE NCAM2 Npn2 RA RALDH2 Z1-4 8 adenylyl cyclase III adenosine monophosphate adenosine triphosphate calmodulin calmodulin kinase II cyclic adenosine monophosphate cyclic guanine monophosphate cyclic nucleotide-gated channel 2,6-dichlorobenzonitrile globose basal cell horizontal basal cell mammalian achaete-scute homologue 1 molecular receptive range NADPH quinone oxidoreductase 1 olfactory/early B-cell factor olfactory marker protein odorant receptor olfactory sensory neuron phosphodiesterase neural cell adhesion molecule 2 neuropilin 2 retinoic acid retinaldehyde dehydrogenase 2 odorant receptor expression zones 1-4 INTRODUCTION “You cannot suppose that atoms of the same shape are entering your nostrils when stinking corpses are roasting as when the stage is freshly sprinkled with saffron of Cicilia and a nearby alter exhales the perfumes of the Orient” Lucretius The human nose can detect an almost unbelievable large number of smells or odors (400 000 distinct odors, (Mori et al., 2006) and references therein) even though our sense of smell is not our most important sense under normal circumstances. The sense of smell is very important to most animals in order to survive, find food, avoid predators and to find a mating partner. I have in this thesis studied different aspects of the functional anatomy of the olfactory epithelium, focusing on one model organism, the mouse. Figure 1. The olfactory epithelium covers the turbinates protruding into the nasal cavity. OSNs in the olfactory epithelium send their axons to target cells in the olfactory bulb which is located in the anterior part of the brain. 9 A perceived odor is made up of one or several distinct odorous molecules, or odorants. Odorants are typically small organic molecules that are detected by the olfactory system by specialized receptor cells in the olfactory epithelium located in the nasal cavity. The main primary olfactory system consists of the olfactory epithelium located in the roof of the posterior nasal cavity and the olfactory bulb located at the most anterior part of the telencephalon. The nasal cavity is in rodents highly convoluted with turbinates protruding into the nasal lumen (fig 1). The olfactory epithelium covers the turbinates, thus resulting in a larger surface area of epithelium specialized to detect odorants. The intricate structure of the turbinates also result in a decrease in air velocity as indicated by computer simulations of the airflow in the rat nasal cavity (Kimbell et al., 1997). It is possible that the decrease in air velocity facilitates sorbtivness of certain odorants. The odorant-containing air that enters the nasal cavity during inhalation is flushed over the epithelium and the odorant molecules stimulate specialized sensory cells, olfactory sensory neurons (OSNs). The OSNs project their axons through the cribiform plate up to the olfactory bulb where the axons make contact with second order neurons. The signal is here refined and transferred to higher areas of the brain where it is interpreted as a scent and the animal becomes aware of the sensation and can respond accordingly. Cell types of the olfactory epithelium The olfactory epithelium is a pseudostratified epithelium consisting of several cell types in which the nuclei form distinct layers; sustentacular cells apically, neuronal cells in the middle compartment and basal cells located closest to the basal lamina (fig 2) (Graziadei & Graziadei, 1979; Nomura et al., 2004). The basal cell layer population contains horizontal basal cells (HBC) and globose basal cells (GBC). The HBCs form a single layer of flat cells situated directly on the basal lamina with round GBCs located above the HBCs. The OSN cell layer consists of both immature 10 and mature OSNs where the mature OSNs constitute the majority of cells in the adult olfactory epithelium. The mature OSNs are located more closely to the nasal lumen compared to the immature OSNs (Graziadei & Graziadei, 1979). The OSN is a bipolar neuron with a single dendrite that terminates in a knob from which 10-20 fine cilia extend into the mucus that covers the epithelium (Menco, 1980; Mendoza, 1993). The cilia are the structures that detect odorant and it is also here the binding of an odorant to the odorant receptor (OR) is converted into electric activity by the intracellular signal transduction cascade. The mature OSN sends a single unmyelinated axon through the cribiform plate to the target in the brain, the olfactory bulb. The sustentacular cells are non-neuronal supporting cells forming a single row of cells occupying the full height of the epithelium although the cells are wider closer to the nasal lumen. The sustentacular cells have Figure 2. The olfactory epithelium consists of several cell types. Sustentacular cells are located closest to the nasal lumen while the middle layers contain mature and immature OSNs. The layers closest to the basal membrane contain basal cells (GBCs and HBCs) of which some cells are the progenitors to OSNs. The epithelium is covered by mucus produced by Bowman’s glands which ducts extend through the epithelium. 11 branched foot processes spreading on the basal lamina. These cells provide support to the OSNs and metabolize xenobiotic compounds and odorants (Ding & Coon, 1988; Chen et al., 1992; Yu et al., 2005). Sustentacular cells are also known to phagocytous dead OSNs (Suzuki et al., 1995). The sustentacular cells are connected with tight junctions resulting in separation of the receptor cell body from the nasal lumen (Miragall et al., 1994; Rash et al., 2005). Bowman’s glands are located in the mesenchyme underneath the olfactory epithelium and their ducts protrude the epithelium. Bowman’s glands produce mucus covering the olfactory epithelium and are like sustentacular cells rich in detoxifying enzymes (Yu et al., 2005). Proliferative capacity of the olfactory epithelium The mammalian olfactory system maintain neurogenesis throughout life (Graziadei & Graziadei, 1979) which makes it an effective model for studying principles of tissue renewal and neuronal cell differentiation in the adult as well as during development. The average life-span of an OSN has been reported to be about 90 days (Mackay-Sim & Kittel, 1991), even though OSNs have been reported to survive for more than 12 months (Hinds et al., 1984). The olfactory epithelium is, due to its unprotected position, easily damaged as a result of exposure to toxic compounds, viral infections or mechanical trauma. The olfactory epithelium is often functionally repopulated with OSNs after damage which is achieved by progenitor cell proliferation and differentiation. The nature of the stem cell of the olfactory epithelium is still unclear; possible candidates have been suggested to reside in the HBC compartment, in the GBC compartment as well as amongst the Bowman’s duct cells (Matulionis, 1975; Graziadei & Graziadei, 1979; Calof & Chikaraishi, 1989; Caggiano et al., 1994; Huard et al., 1998). HBCs proliferate at a low rate where about 2% of the actively dividing cells in the basal compartment of the normal epithelium are HBCs (Holbrook et al., 1995; Huard & Schwob, 1995). HBCs have been 12 indicated by in vitro experiments to give rise to both OSNs, glia cells and HBCs (Carter et al., 2004). Nevertheless, the general view is that the stem cell is a member of the GBC population (for review on regeneration of olfactory epithelium see (Schwob, 2002)). GBCs have been suggested to give rise to OSNs in vivo (Mackay-Sim & Kittel, 1991; Schwartz Levey et al., 1991; Caggiano et al., 1994; Gordon et al., 1995) and proliferation in the GBC layer is increased after damage to the primary olfactory system, for example by bulbectomy (Schwartz Levey et al., 1991; Suzuki & Takeda, 1991), axotomy (Suzuki & Takeda, 1991) or damage by compounds that are toxic to the olfactory epithelium. After injury to the olfactory epithelium by the gas methyl bromide are areas with GBCs repopulated with all cell types of the olfactory epithelium whereas areas where GBCs are absent will become an epithelium with respiratory epithelial character (Jang et al, 2003). Areas that are repopulated sometimes lack HBCs during an early phase of recovery (Schwob et al., 1995). Moreover, some GBCs exhibit multipotency since highly purified GBCs transplanted into methyl bromide lesioned epithelium will give rise to clonal clusters containing both neural (OSNs and GBCs) or non-neural cells (Bowman’s glands, sustentacular cells and cells of the respiratory epithelium) or clusters containing both non-neural cells and neural cells, indicating the existence of different GBC populations (Chen et al., 2004). A subset of GBCs remains quiescent during long time-periods as observed by retention of intensity of labeled genomic DNA and this subset may resume active cell cycling after damage by methyl bromide (MackaySim & Kittel, 1991). During regeneration as well as normal turnover of the olfactory epithelium, GBCs function as the immediate progenitors to the OSNs. The GBCs are the cells that have the highest proliferative rate of all cells in the olfactory epithelium. About 97,2% of the actively dividing cells in the basal compartment of the normal epithelium are GBCs (Huard & Schwob, 1995). When a progenitor in the GBC population divides it gives rise to immature OSNs expressing immature markers (e.g. SCG10 (Camoletto et al., 2001)), the immature OSNs will, in time, mature to 13 functional OSNs expressing mature markers (e.g. olfactory marker protein (OMP) (Farbman & Margolis, 1980)) that establish functional contact with the olfactory bulb. Odorant receptors Odorant molecules are bound to and detected by odorant receptors (ORs) located on the cilia of the OSNs. The ORs were first identified in the rat in 1991 (Buck & Axel, 1991) and were predicted to be G proteincoupled receptors that span the cell membrane seven times. Mice have roughly 1,300 OR genes of which about 20% are predicted to be pseudogenes (Zhang & Firestein, 2002; Godfrey et al., 2004). The importance of this sense for many animals may be mirrored by the abundance of ORs. The OR family is the largest gene superfamily in the mammalian genomes (Zhang & Firestein, 2002). Humans have, in comparison to the mouse, few OR coding genes and many genes are predicted to be pseudogenes (roughly 640 genes of which 46% are pseudogenes) (Malnic et al., 2004). The mouse OR genes are organized in 27 clusters containing multiple OR genes and clusters are located on all chromosomes except 12 and Y (Sullivan et al., 1996; Zhang & Firestein, 2002). The OR sequences separate in two broad classes, class I and class II receptors (Zhang & Firestein, 2002). Class I ORs was first identified in fish (Ngai et al., 1993b) and was later identified in humans (Glusman et al., 2001) and mouse (Zhang & Firestein, 2002). All class I ORs are located in a cluster on chromosome 7 (Tsuboi et al., 2006). In the mouse, there are 147 class I OR genes of which 120 are potentially functional (Zhang & Firestein, 2002). The OR protein is encoded by a gene with a single coding intronless exon of about 1 kilobase pairs (e.g. (Sosinsky et al., 2000)). The OR protein has seven transmembrane regions where transmembrane domains 4 and 5, and to a lesser extent transmembrane domain 3, are 14 the most variable regions (Pilpel & Lancet, 1999). Computer models of two ORs indicate that the recognition of an odorant by its OR is mediated by amino acids in the transmembrane domains 3 to 6 (Singer & Shepherd, 1994; Katada et al., 2005), where point mutations can change ligand specificity (Katada et al., 2005). Signal transduction in olfactory sensory neurons The detection of an odorant starts with odorant binding to the G protein-coupled OR. The activation of ORs by odorants starts an intracellular signal transduction cascade in the tiny volume of the cilium, which in the end results in an action potential in the OSN (fig 3). Odorant binding to the OR results in the activation of a heterotrimeric G protein. The Gα-subunit Gαolf is present at high levels in the olfactory cilia (Jones & Reed, 1989) and stimulates the transmembrane adenylyl cyclase III (ACIII) (Pace et al., 1985; Bakalyar & Reed, 1990) to convert adenosine triphosphate (ATP) to cyclic adenosine monophosphate (cAMP). Elevation of intraciliary cAMP opens cyclic nucleotide-gated (CNG) channels (Nakamura & Gold, 1987) that are non-selective cation channels (Balasubramanian et al., 1995; Dzeja et al., 1999). The opening of the CNG channels results in an increase in intraciliary Ca2+ and Na+ ions (Leinders-Zufall et al., 1997) resulting in a slight depolarization of the ciliary membrane. The increase in Ca2+ in turn opens Ca2+-activated Cl--channels (Kleene, 1993; Reisert et al., 2003) driving Cl- to exit the cell due to its high intracellular concentration and its release hence further depolarize the cell (Reuter et al., 1998). The resulting receptor potential is then converted to trains of action potentials propagating to the axon terminals. There is a general consensus that the vast majority of OSNs react to odorant exposure by activating ORs that are linked to stimulation of adenylyl cyclase by Gαolf, since single knock-outs of either of the transduction components Gαolf, ACIII or the CNG channel subunit 15 Figure 3. Schematic picture of the signal transductions cascade in OSNs in response to odorant binding to the OR. Adaptation is mediated by feed-back mechanisms on several levels of the transduction cascade provided by increased levels of intracellular calcium. CNGA2 result in animals that are anosmic for a wide range of odorants (Brunet et al., 1996; Belluscio et al., 1998; Wong et al., 2000). However, there might be exceptions since mice genetically null for CNGA2 still detect some odorants (Lin et al., 2004). There have been reports of OSNs not expressing either Gαolf, ACIII or CNGA2 but that instead express guanylyl cyclase-D, cyclic guanine monophosphate (cGMP)-stimulated phosphodiesterase and a cGMP-selective cyclic nucleotide gated channel (Juilfs et al., 1997; Meyer et al., 2000). Inositol-1,4,5trisphosphate and diacylglycerol produced by phospholipase C have also been implicated in odorant signal transduction (reviewed in (Elsaesser & Paysan, 2005)). The roles of these alternative signaling pathways are currently unresolved. Six CNG channel genes have been identified in the mammalian genome, CNGA1-4, CNGB1 and CNGB3 (for a review of CNG channels see (Kaupp & Seifert, 2002)). CNG channels in OSNs are most likely 16 heterotetramers (Liu et al., 1996) and have a fixed stoichiometry of two CNGA2 subunits, one CNGA4 and one CNGB1b (Zheng & Zagotta, 2004). CNGA2 forms functional homomeric channels when expressed in a heterologous system while CNGA4 and CNGB1b only form functional channels as heteromers with CNGA2 suggesting that CNGA2 is the functional channel and CNGA4 and CNGB1 have roles in modulating the channel properties (Kaupp et al., 1989; Bradley et al., 1994; Liman & Buck, 1994; Zheng & Zagotta, 2004). Feed-back mechanisms likely ensure that the transduction cascade is terminated and may result in OSNs adjusting the sensitivity, adapting, to different stimulus intensities to prevent saturation of the transduction machinery. The process of signal termination and adaptation occurs already at the level of signal transduction by feedback of Ca2+ that has modulatory roles at various steps in the transduction cascade (adaptation reviewed in (Zufall & Leinders-Zufall, 2000)). Ca2+ is important both in signal termination and adaptation where the adaptive actions of Ca2+ can be divided into two categories; those on the CNG channel itself and those on other components in the transduction cascade. The principal mechanism underlying signal termination and adaptation is modulation of the CNG channel by Ca2+ feedback (Kurahashi & Menini, 1997). This inhibition is a result of a complex between Ca2+ and calmodulin (CaM) where, at high Ca2+ concentrations, Ca2+-CaM reduces the sensitivity of the CNG-channel to cAMP 10-fold (Chen & Yau, 1994; Trudeau & Zagotta, 2003). Negative modulation by Ca2+ through CaM on CNGs requires the subunits CNGB1b and CNGA4 (Bradley et al., 2001; Munger et al., 2001; Bradley et al., 2004). It has been suggested that CNGA4 null mice exhibit slower Ca2+-CaMmediated channel desensitization (Munger et al., 2001) and have problems with detecting odorants in a background of another odorant, suggesting that CNGA4 is important for adaptation (Kelliher et al., 2003). The inhibitory effect of Ca2+ on the CNG channel is fast and this can be achieved by constitutive binding of CaM to the CNG channel (Bradley et al., 2004). In addition of regulating the sensitivity of the CNG channel to cAMP, Ca2+-CaM also activates a phosphodiesterase 17 (PDE) resulting in breakdown of cAMP to adenosine monophosphate (AMP) and thus reduces the level of cAMP (Borisy et al., 1992; Yan et al., 1995). Ca2+-CaM also influences the level of cAMP by inhibiting adenylyl cyclase by the action of the activated kinase calmodulin kinase II (CamKII) that phosphorylates ACIII (Wei et al., 1996; Wei et al., 1998; Leinders-Zufall et al., 1999). The effect of Ca2+-CaM on the cAMP level in the cilia is somewhat slower than the effect on the CNG channel. The response of an odorant in the OSN can be influenced by other adaptive processes like phosphorylation of the receptor via G-proteincoupled receptor kinase 3 which is mediated by the βγ subunits (Boekhoff et al., 1994; Peppel et al., 1997). The amount of cGMP is, as well as the level of cAMP, increased after odorant stimulation. The rise in cGMP concentration after odorant stimulation is both slower and more sustained than the increase in cAMP (Zufall & Leinders-Zufall, 1997; Moon et al., 2005). Odorant receptor expression patterns Shortly after the cloning of the first ORs it was evident that each OR is expressed in a small fraction of OSNs, starting at embryonic day 11.5 (Strotmann et al., 1992; Ressler et al., 1993; Vassar et al., 1993; Sullivan et al., 1995). Each OSN expresses only one receptor (Chess et al., 1994; Malnic et al., 1999) in a monoallelic and mutually exclusive manner (Chess et al., 1994; Serizawa et al., 2000). Exception to the singular expression of ORs exist as an OSN have been reported to express two different ORs (Rawson et al., 2000). Several mechanisms behind the mutual exclusive expression of the ORs have been suggested, including irreversible DNA changes like DNA recombination or gene conversion as well as reversible mechanisms. By cloning mice from post mitotic OSNs two studies have shown that irreversible DNA changes in the nuclei of OSNs is not the mechanism 18 behind OR gene selection (Eggan et al., 2004; Li et al., 2004). The resulting cloned mice do not display monoclonal expression of ORs but rather have an olfactory epithelium expressing several types of ORs, in a similar manner as control mice. A cis-acting DNA region, called the Hregion, was suggested to activate expression of OR genes in a specific OR gene cluster and to restrict the expression to only one out of the three OR genes present in a transgenic mouse line (Serizawa et al., 2003). The H-region was later implied to mediate monoallelic expression of all ORs (Lomvardas et al., 2006). The H-region exists in two copies in each cell and one allele is methylated in OSNs. The methylation is implied in inactivation of one allele in order to achieve one functional H-region and expression of only one OR gene copy. A functional OR protein might stabilize expression of the chosen OR and Figure 4. The expression areas of four ORs in one nostril are shown by radioactive in situ hybridizations, positive signal in white. ORs are expressed in restricted areas of the olfactory epithelium in scattered cells where the most dorsomedial expression area is called zone 1 (Z1) and the most ventrolateral is called zone 4 (Z4). 19 exclude expression of additional ORs as the expression of a pseudogene fail to inhibit expression of a functional OR gene (Serizawa et al., 2003) and the presence of an untranslatable OR coding sequence in the mRNA is not sufficient to exclude expression of a second OR (Lewcock & Reed, 2004). OSNs expressing a given OR are not spread over the entire epithelial sheet but are rather restricted to broad areas of the epithelium. A zonal topography is evident at embryonic day 13.5 when enough cells are expressing ORs to be able to discern a pattern (Sullivan et al., 1995). The olfactory epithelium is classically divided into four zones based on the expression pattern of a few ORs (Ressler et al., 1993; Vassar et al., 1993). The OSNs expressing a particular OR are intermingled with other OSNs expressing other ORs within such an expression zone (Ressler et al., 1993). The zones are distributed in a dorsomedial to ventrolateral orientation where zone 1 (Z1) is the most dorsomedial zone and zone 4 (Z4) is the most ventrolateral zone (fig 4). Other patterns of expression have also been identified (Kubick et al., 1997; Strotmann et al., 1999). Expression differences can be observed between class I and class II ORs where class I ORs are expressed predominantly in Z1 (Tsuboi et al., 2006), class II ORs, on the other hand, are expressed in any of the zones, but are in Z1 intermingled with OSNs expressing class I ORs (Zhang et al., 2004; Tsuboi et al., 2006). Small regions upstream of the transcription start site of OR genes have been implicated in the regulation of the zonal specific expression (Qasba & Reed, 1998; Serizawa et al., 2000; Rothman et al., 2005). These regions contain homeobox and olfactory/early B-cell factor (O/E) binding sites where a minimal promoter containing one homeobox and one O/E site faithfully mimic the spatial distribution of one OR studied (Rothman et al., 2005). The number and order of motifs (e.g. homeobox and O/E sites) may regulate the pattern of OR expression by modulating the probability of expression of different ORs differentially across the olfactory epithelium (Rothman et al., 2005; Hoppe et al., 2006). 20 Projections to the brain In the olfactory epithelium axons of OSNs form loose bundles between the "feet" of HBCs, traveling in a rostro-caudal direction. As they cross the basal lamina and exit the epithelium, processes of olfactory ensheating cells tightly enwrap the axon bundles (Holbrook et al., 1995). Ensheating cells can be observed forming tunnels containing hundreds of OSN axons traveling to make synaptic contacts with target neurons in the olfactory bulb (Li et al., 2005). The axons pass the cribiform plate and form, together with glial cells, the nerve layer surrounding the olfactory bulb. The axons defasciculate upon reaching the olfactory bulb and target different regions of the bulb (Whitesides & LaMantia, 1996). The OSN axons terminate in anatomically distinct structures called glomeruli (Graziadei & Graziadei, 1979) where Figure 5. OSNs expressing the same OR send their axons to the same glomeruli and make synaptic contacts with target cells e.g. mitral cells. Mitral cells send the olfactory signal further up to the olfactory cortex. Granule cells make dendrodendritic contacts between different mitral cells and regulate mitral cell output to the olfactory cortex. The olfactory bulb has distinct layers formed by different cell types. 21 synaptic connections are made with e.g. principal projection neurons (fig 5). The mouse olfactory bulb contains about 1800 glomeruli (Royet et al., 1989). Neurons expressing the same OR converge upon the same glomeruli, usually one medial and one lateral (first noticed by (Ressler et al., 1994; Vassar et al., 1994)). The zonal organization is maintained in the olfactory bulb (Alenius & Bohm, 1997; Yoshihara et al., 1997; Gussing & Bohm, 2004; Miyamichi et al., 2005) where the position of OR expression in the olfactory epithelium determines the dorsal/ventral positioning of target glomeruli in the olfactory bulb (Miyamichi et al., 2005). The glomerulus targeted by a particular OR is located approximately in the same position between individuals (fig 6) (Strotmann et al., 2000), forming a stereotype map of projections from the epithelium to the olfactory bulb. OSNs expressing highly homologous receptors from the mOR37 subfamily project to glomeruli that are situated close to each other in the olfactory bulb (Strotmann et al., 2000). In the fruit fly Drosophila, OSNs expressing similar ORs are not clustered in the periphery but OSN expressing the same OR project to the same glomeruli and OSNs expressing similar ORs project to closely positioned glomeruli (Couto et al., 2005). Each glomerulus receives input from thousands of OSNs which in the glomerulus make synaptic connections with second order neurons, i.e. mitral and tufted cells that in turn project to higher olfactory centras. Each glomerulus contains the dendrites of several mitral and tufted cells and each mitral cell innervates one single glomerulus ((Schoppa & Urban, 2003) and references therein). There are two classes of interneurons in the olfactory bulb; the periglomerular cells and the granule cells. Cells of both types are added throughout life of the animal (Kaplan et al., 1985) and modulate the activity of OSNs as well as the mitral and tufted neurons. Periglomerular cell synapses are mostly restricted to one glomerulus where they mediate intraglomerular interactions. Granule cells make synaptic connections between mitral cells associated with different glomeruli. The synaptic organization of the glomerulus is reviewed in (Schoppa & Urban, 2003; Kosaka & Kosaka, 2005). 22 The axons from the olfactory bulb constitute the lateral olfactory tract projecting to the olfactory cortex. Olfactory cortex is defined as the cortical areas receiving direct input from the olfactory bulb and includes, among several areas, the anterior olfactory nucleus, the entorhinal cortex, the piriform cortex and the olfactory tubercle. A transgenic mouse line expressing Barley lectin under the control of specific promoters made it possible to chart neural circuits due to the ability of the lectin to transfer transneuronally (Horowitz et al., 1999). It was indicated, by expressing Barley lectin under the control of two different OR promoters, that the projections from the olfactory bulb to cortical areas display some similarity between individuals. The inputs originating from ORs, diverge in the olfactory cortex to clusters of thousands of neurons overlapping with clustered inputs from other ORs (fig 6) (Zou et al., 2001). These findings are supported by a study using c-Fos as a neuronal activity marker where single odorants activate a small subset of cells in the anterior piriform cortex. The different odorants had different but overlapping representations of activated cells (Zou et al., 2005). Figure 6. OSNs expressing the same OR project to the same glomerulus and the zonal topography of OR expression in the olfactory epithelium is maintained in the projections to the olfactory bulb. In contrast, the projections to the olfactory cortex are overlapping between axons coming from different areas of the olfactory bulb. The projections from the olfactory epithelium to the bulb and cortex are similar between individuals. 23 Initial steps in constructing a smell in the brain In the environment, thousands of chemical molecules exists that need to be detected and identified in order for the animal to be able to interpret the world. Large numbers of structurally diverse odorant molecules interact with the ORs. It is likely that more chemicals can be smelled than there are ORs for them, making a combinatorial representation necessary. Computer models indicate that air entering the rat nasal cavity first flows through dorsomedial areas and then courses more peripherally (Kimbell et al., 1997; Zhao et al., 2006). The uptake of odorants in the mucus may be changed by changing airflow velocity over the epithelium so that efficient adsorption of odorants is achieved in areas expressing ORs most sensitive to the odorant ((Schoenfeld & Cleland, 2005) and references therein). The odorant is, after being adsorbed in the mucus covering the olfactory epithelium, identified by ORs located on the surface of the OSNs. An OSN expresses only one out of the thousand ORs in the mouse genome and it is hypothesized that an OR recognizes a particular chemical structure or molecular feature, for example a specific polar group or molecules with a certain carbon chain length (Malnic et al., 1999). Hence an OSN can be activated by one particular chemical structure; the structure identified by the OR. An odorant can, because it often contains several chemical structures, be detected by several ORs (Araneda et al., 2000) and thus several different types of OSNs. The range of odorants that activates an OR can be called the ORs molecular receptive range (MRR). In vitro studies have indicated that ORs that are >60% identical has a high probability of being stimulated by odorants with related structures (Malnic et al., 1999; Kajiya et al., 2001). The structure of an odorant might thus be coded by activation of a specific subset of OSNs (Sato et al., 1994; Malnic et al., 1999; Ma & Shepherd, 2000; Kajiya et al., 2001). The perceived quality of an odorant can change when the odorant concentration is changed. The explanation for this may be that the ORs 24 have different affinities to odorants and at higher odorant concentration a recruitment of additional receptors occurs (Amoore, 1982; Ma & Shepherd, 2000; Kajiya et al., 2001). The olfactory information that is extracted from the environment by the OSN is influenced by the OR repertoire of the animal. Comparison of human and mouse OR subfamilies shows that the two species have many, but not all, subfamilies in common. However, mouse subfamilies are usually larger than the human counterparts. This finding suggests that humans and mice recognize many of the same odorant structural motifs, but mice may be superior in odor discrimination and may be superior in identifying different odorants (Godfrey et al., 2004). The inability to detect specific odorants, so called specific anosmia, has been described both in humans and rodents. One example is the mouse strain C57Bl/6 that is anosmic to the odorant isovaleric acid (Wysocki et al., 1977). The anosmia was later found to be associated with a locus containing an OR cluster, indicating that loss of ORs may be the cause of the specific anosmia (Griff & Reed, 1995; Zhang & Firestein, 2002). The identification of ligands for ORs has been hampered by the difficulty to express ORs at the surface in heterolougous cell lines and up to date a few proteins have been identified that facilitate cell surface localization of some of the ORs (Saito et al., 2004; Neuhaus et al., 2006). The first in vivo identification of a ligand to an OR was made in the rat when the I7 receptor was expressed in OSNs normally not expressing that OR by employing adenovirus mediated delivery. The OSNs displayed physiological responses to the odorant octanal (Zhao et al., 1998). Since then several identifications of ligands have been made in cell culture. Some of the mouse ORs, identified as having ligands such as aliphatic acids, alcohols and aldehydes, are known to belong to class I ORs (Malnic et al., 1999; Saito et al., 2004). All OSNs expressing a certain OR converge to the same glomeruli. The response profile of a glomerulus is determined by the OR expressed by the OSNs projecting to it (Bozza et al., 2002) and the MRR of the glomerulus is thus the same as the MRR of the OSNs that innervate the 25 glomerulus. The odorant stimuli received by the olfactory epithelium thus appear to be converted to a topographic map of activated glomeruli in the olfactory bulb (Ngai et al., 1993a; Ressler et al., 1994; Vassar et al., 1994; Malnic et al., 1999; Rubin & Katz, 1999; Uchida et al., 2000; Fried et al., 2002). Imaging studies demonstrate that different odorants elicit defined patterns of glomerular activation, revealing a functional representation of the anatomical map (Rubin & Katz, 1999; Uchida et al., 2000; Wachowiak & Cohen, 2001). OSNs expressing a limited number of highly homologous receptors project to glomeruli that are closely juxtaposed in the olfactory bulb (Strotmann et al., 2000) which is consistent with clustered glomerular responses to odorants with similar structure (reviewed in (Mori et al., 2006)) and the fact that highly homologous receptors detect molecules with related structures (Malnic et al., 1999; Kajiya et al., 2001). Imaging studies of a limited area of the olfactory bulb have suggested that the response patterns after activation with natural complex odors are highly specific and that the number of glomeruli activated by a complex odor are well predicted by the sum of activated glomeruli after stimulation with the individual components of the complex odor (Lin et al., 2006). The OSN axons make synaptic contacts with mitral and tufted cells in the glomeruli, which then project to the olfactory cortex. The signals in the glomeruli are processed by local interneurons like periglomerular cells and granule cells. Mitral cells connected to glomeruli that respond to a wide range of related odorants receive inhibitory inputs from neighboring glomerular units via lateral inhibition. Lateral inhibition is mediated by granule cells via reciprocal dendrodendritic connections between mitral cells from different glomeruli, thereby regulating mitral cell axonal output (Rall et al., 1966; Isaacson & Strowbridge, 1998; Margrie et al., 2001). Moreover, strongly activated glomeruli suppress transmitter release in surrounding glomeruli via excitatory actions on periglomerular cells (Aungst et al., 2003; Vucinic et al., 2006). The glomerulus that is exhibiting the strongest activation thereby inhibits the other activated glomeruli. The MRR of a mitral cell is thus not the same as the MRR of the OSN targeting the glomerulus, but is modified 26 by interactions with other glomeruli. The olfactory bulb thus functions to sharpen the signal coming from the olfactory epithelium before it is sent to the olfactory cortex (Mori et al., 1999). After processing in the olfactory bulb, the signal is transferred to olfactory cortex where mitral cells axons show overlapping patterns of innervations. Two mitral cell responding to different odorants may make contact with the same olfactory cortex neuron and individual neurons in the olfactory cortex can respond to multiple odorants (Haberly, 1969; Tanabe et al., 1975). Increasing the odorant concentration results in higher number of activated cells. Neurons activated by a certain odorant are scattered in cortex (Cattarelli et al., 1988) and neurons projecting from two different glomeruli have overlapping termination sites (Zou et al., 2001). There have also been reports on cells that act as coincidence detectors, cells that are activated by binary mixtures of odorants but not activated by the individual odorant components (Zou et al., 2005; Zou & Buck, 2006). These studies suggest integration of signals from different ORs at the level of the olfactory cortex which may be part of the processing that eventually leads to an odor percept (Zou et al., 2005). Olfactory cortex in turn projects to subcortical, limbic and neocortical regions involved in motivated behavior, autonomic reflexes and hormonal modulation. 27 AIM OF THE THESIS The reason for the spatially restricted expression of ORs and the division of the epithelium into several zones that express unique ORs are up to date not known. I have in this thesis studied the olfactory epithelium of the mouse focusing on molecular and anatomical differences between OR expression zones of the olfactory epithelial sheet. Specific aims were to: - Identify genes whose expression pattern corresponds to the spatial expression of ORs in order to find a mechanism used in the specification of the OSNs. - Study zonal activation of OSNs after odorant exposure in order to examine the response properties of the ORs. - Explore differences in tissue homeostasis over the olfactory epithelium in intact as well as in damaged epithelium. 28 METHODS Studying odorant detection threshold A number of behavior assays have been used to measure olfactory detection in mice and rats. One of the most simple is the hidden cookie test were a piece of food is hidden in the bedding of the cage or another arena. The animal is then introduced into the arena and allowed to explore the area and to find the hidden snack. A course estimate of the animals olfactory abilities can then be based on the time it takes for the animal to achieve the task. This test contains a lot of parameters that is not related to olfaction like activity and attention of the animal. A test with a little better resolution is the two choice paradigm where the animal is conditioned to an odorant by simultaneous presentation of a reward during the training phase. During the test period the animal is presented with the test odorant and an unrelated odorant, both without the reward. A positive answer would here be if the animal chooses to go to the conditioned odorant. The concentration of the odorants can be altered to get a course estimate of the detection threshold. Although these methods are simple and do not require any special equipment they are either too unspecific or require much work in order to obtain higher resolution of detection threshold values. Automated operant conditioning enables the analysis of behavior in a standardized way with relatively little time required for handling the animals compared to the quality of the result obtained. Slotnick and colleagues have worked on automated operant conditioning for odorant detection and then mainly in the rat (e.g. (Slotnick & Bodyak, 2002; Slotnick et al., 2004)). Almost all of the mammalian research of the molecular biology of olfaction has been done in the mouse which is an animal that has not been widely used in operant conditioning studies of sensory functions. The increasing use of gene targeted mice in 29 Figure 7. The olfactometer is used to analyze detection or discrimination of odorants. This setting was used in paper III to analyze for effects on odorant detection thresholds after selective ablation of OSNs. In the lower picture a mouse is inserting the nose in the odor sampling port, trying to detect an odorant in order to get a reward in form of a drop of water. molecular biology and the need to measure olfactory capabilities in such strains motivated Slotnick and co-worker to test mice in a modified olfactometer originally designed for rats (Bodyak & Slotnick, 1999). The test showed that mice do perform well on a go-no go task that can be used to determine the detection thresholds of some odorants in wild type mice. We have analyzed the detection threshold for a few odorants in adult mice subjected to OSN ablation as described in paper III. The olfactometer used in our study and by (Bodyak & Slotnick, 1999) is an 30 eight-odor olfactometer (see fig 7). Air is passed through a carbon filter and over the surface of diluted odorants placed in odor saturator bottles. The air is then presented to the mouse at a sampling port. The odorant that reaches the mouse is 2.5% of the concentration in the headspace above the odorant in liquid. The initial training is aimed at teaching the mouse to lick at a reinforcement tube in the sampling port when an odorant is presented. The lick is what is measured and automatically recorded. The first stage trains the mouse to lick at the tube for a drop of water. The mouse is kept at a restricted water schedule in order to ensure that the mouse is motivated during the experiment. The second stage aims at training the mouse to withdraw the snout before a new trial can be initiated. In the third stage odorant is introduced and the animal receives the reward after sampling the odorant. When a mouse is properly trained, it responds to odorant presentation by licking at the reinforcement tube only when odorant is presented. Odorant and clean air is presented in random order in blocks of 20 trials during testing. The mouse is run in a total of five consecutive blocks (a session). Licking when odorant is presented and not licking when there is no odorant are considered as correct responses. Not licking when odorant is presented or licking when there is no odorant are considered as incorrect responses. If the mouse achieves 90% correct responses in a single block it is considered to have reached criterion performance and is tested in a new session with a lower concentration of the odorant investigated. The mice were in paper III tested for three sessions per day where a maximum of two sessions were at the same concentration to ensure that the mouse does not fail to show that it detects a certain odorant concentration due to inter daily fluctuations in factors beyond our control. When the mouse has failed to detect the odorant of a certain concentration in three sessions the test is terminated and the mouse is assigned the preceding concentration as the detection threshold. One has to make a number of precautions in order to be sure that the mouse actually responds as a consequence of sensing the odorant and 31 not due to e.g. sound cues from the valves or from odor cues from tubing or the odor saturator bottles. For example, we use “distraction valves” that open randomly when the odor valves are operated. The positions of the saturator bottles for one particular concentration are changed, both within and between sessions, so that the mouse does not easily learn to distinguish between the sounds of opening or closing of odor valves. Moreover, we frequently change the tubing and odor saturator bottles. The odor saturator bottles are changed daily and washed in ethanol and stored at 70°C. The tubing can easily be pinched above the odor saturator tubes in order to close the odor stream which is yet another way to test if the mouse responds to the odorant and not to other cues. The most reliable way to test for cues is to run a trained mouse. Cues exist if the mouse is able to get high number of correct responses when no odorant or a concentration below detection threshold is used. Since the publication of our study, Clevenger & Restrepo have reported on an alternative protocol for determining a odorant detection threshold using the olfactometer (Clevenger & Restrepo, 2006). Instead of using descending concentrations of odorants they used random presentation of odorant concentrations to “home in” on the detection threshold. Alternating presentation of different odorant concentrations also decrease the number of trials needed. 32 RESULTS AND DISCUSSION Gene expression correlated to the zonal topography of ORs (paper I) ORs are expressed in limited regions of the olfactory epithelium. The epithelium has classically been divided into four zones based on OR expression (Ressler et al., 1993; Vassar et al., 1993). We could in paper I, however, show that Z2-4 ORs have overlapping areas of expression while Z1 ORs appear not to overlap thus forming a distinct border between Z1 and Z2. Later studies confirm and extend our findings on the overlapping expression of ORs in Z2-4 (Iwema et al., 2004; Miyamichi et al., 2005). It was suggested, by studying a relatively large number of ORs, that each OR located in Z2-4 has its own unique expression area which is overlapping between different ORs, forming broad but overlapping bands of OR expression (Miyamichi et al., 2005). The possibility exists that Z1 is inherently different from the other zones as this zone is the only zone that expresses both class I and class II ORs (Tsuboi et al., 2006). Z1 and Z2-4 are different from each other with regard to expression of other genes as well, e.g. expression of NADPH quinone oxidoreductase 1 (NQO1) in Z1 (Gussing & Bohm, 2004) and neural cell adhesion molecule 2 (NCAM2) in Z2-4 (Alenius & Bohm, 1997; Yoshihara et al., 1997). The function of the zonal topography and the mechanisms that control OSN zonal specification and specific zone projection are up to date not completely understood. OR zones may be established during early development when OSNs are generated as the zonal topography is evident already at embryonic day 13.5 (Sullivan et al., 1995). New OSN are, however, continuously generated throughout life and the zonal arrangement maintained. Specification of OSNs, i.e. OR choice, is independent of the target neurons (Sullivan et al., 1995; Fan & Ngai, 2001). Conceivably, information within and/or in proximity of the 33 olfactory epithelium may continuously contribute to restrict OSNs to express ORs and axon guidance proteins according to the cell's zonal localization (e.g. (Alenius & Bohm, 1997), paper I). The fact that Z2-4 ORs display overlapping expression areas indicate that the regulation of OR choice may not be regulated by zonally distinct transcription factors but may rather be regulated by gradients of instructive factors. In a screen aimed at identifying genes displaying expression patterns correlated to the expression of ORs we have identified several genes. Msx1, Alk6, retinaldehyde dehydrogenase 2 (RALDH2) and Neuropilin2 (Npn2) are expressed in a graded manner in or in close proximity to the olfactory epithelium (fig 8). Zone-specific expression of ORs is in part regulated by DNA regions upstream of the OR genes. The correct expression pattern of an OR (M71) has been implied to depend on only a small DNA region close to Figure 8. OR expression areas are overlapping in Z2-4 but not in Z1. Alk6 are expressed in sustentacular cells (sus) in a graded manner with high expression in Z1 and low expression in Z4. Reverse gradients of Npn2, Msx1 and RALDH2 was observed in OSNs, basal cells (bc) and in the lamina propria, respectively. 34 the transcription start site. This minimal promoter contains an O/E binding site and a homeobox binding site (Rothman et al., 2005). The identification of the same common regulatory regions in several OR genes expressed in different zones indicates that common regulatory proteins may be involved in the choice of which OR an OSN expresses. If so, it is unlikely that expression of regulatory factors would follow zonal borders but may instead by graded over the olfactory epithelium. A candidate is the homeobox protein Msx1, identified in paper I to be expressed in a graded manner in the basal cell layer of the epithelium as this cell layer contains the progenitors of the OSNs. Retinoic acid (RA) is a molecule crucial to olfactory development (Mic et al., 2002) that has been implicated in Msx1 regulation (Shen et al., 1994; Wang & Sassoon, 1995; Song et al., 2004). RA is a possible candidate for regulating the low dorsomedial to high ventrolateral expression of Msx1 as the rate-limiting enzyme in RA synthesis, RALDH2, is expressed in mesenchymal cells as a gradient that follows the Msx1 gradient. Our data correlates with previous work as cells responding to RA are located in lateral areas of the olfactory epithelium (Whitesides et al., 1998). Moreover, RA is suggested to be important for the formation of lateral structures of the olfactory epithelium during early embryogenesis (LaMantia et al., 2000). Mature OSNs are dependent on RA for their survival (Hägglund et al., 2006) and RA deficiency has been shown to increase cell proliferation in the olfactory epithelium in rat (Asson-Batres et al., 2003). The identification of genes with graded expression in the sustentacular cells, basal cells and in cells in the mesenchyme makes it likely that graded expression can be observed in OSNs as well. The fact that the localization of OSNs expressing a given OR strongly influences the projection sites in the olfactory bulb so that the OR location in the epithelium determines the dorsal-ventral position of the glomeruli (Miyamichi et al., 2005) suggests that there may be gradients of molecules regulating projections to the target glomeruli. We found that Npn2, involved in axon guidance, is expressed in a gradient in OSNs. It 35 can be hypothesized that different levels of Npn2 may help in guiding the axons towards their correct glomeruli. Npn2 is however not a major factor in the zonal topography of projections to the olfactory bulb as mice without a functional Npn2 gene do not display gross defects in the guidance of the axons to their correct glomeruli (Walz et al., 2002). The Npn2 mutant mice however have axons that overshoot their target glomeruli and instead extend into deeper layers of the olfactory bulb. Target cells in the olfactory bulb express the ligands Sema3B and Sema3F (paper I) which mediate repulsive actions when interacting with Npn2 and may function to stop axons in the right compartment of the olfactory bulb. Nevertheless, reasons for the graded expression of Npn2 are currently unknown. 36 Regionally restricted detection of odorants (papers II and III) A number of ligand-receptor matches have been made in heterologous as well as in homologous in vitro systems, e.g. (Zhao et al., 1998; Malnic et al., 1999). These results suggest that the ORs are relatively broadly tuned, i.e. a receptor can be activated by quite a range of odorant molecules and one odorant can activate several different receptor types. These studies are however made for a limited number of odorants and ORs and do not take the spatially restricted expression of OR genes in the olfactory epithelium or the number of responding cells into account. Electro-olfactograms have shown that the response to odorant exposure may vary over the olfactory epithelial sheet but if such variation is correlated to zonal topography has not been addressed (Thommesen & Doving, 1977; Scott et al., 2000). In papers II and III we have aimed to analyze for topography of odorant-activated OSNs in their proper context, i.e. the olfactory epithelium of behaving mice. We show that this is possible by using c-Fos as an activity marker after odorant exposure (paper II) and by measuring odorant detection thresholds of mice subjected to selective ablation of OR zones (paper III). Odorants induce c-Fos in OSNs in an OR-like pattern We have exposed live mice to a number of odorants in order to activate ORs and hence OSNs. The epithelium was analyzed for changes in expression of genes known to be regulated by neural activity in order to be able to identify odorant-activated OSNs. c-fos is an immediate-early gene that has been used as an activity marker in other systems in response to stimuli (reviewed in (Herdegen & Leah, 1998)). Normally the epithelium is negative for c-Fos but in paper II we have found that c-Fos is detectable in the olfactory epithelium after exposure to odorant. c-Fos is readily distinguished 20 minutes after odorant exposure and both mRNA and protein can be detected several hours after odorant 37 exposure, indicating that long-lasting changes in gene-regulation occur in the OSNs as a result of odorant exposure. Preferentially mature OSNs respond since 90% of cells positive for c-Fos are also OMP positive. Several odorants with varying chemical structure, such as pyridine, pyrazine, trimethylphosphite, cyclohexanone and octanal, α-ionone, benzene and benzaldehyde could induce c-Fos. It was soon evident that c-Fos positive OSNs are stochastically distributed in the olfactory epithelium. The expression frequency of a particular OR is often between 0.1-1% (Ressler et al., 1993). The number of cells showing induced c-Fos after odorant is similar to the number of OSNs positive for one OR. OSNs responding with c-Fos induction are restricted to limited areas of the epithelial sheet, e.g. benzaldehyde induces c-Fos in areas also expressing Z1 ORs and octanal induce c-Fos in areas with more laterally expressed ORs (Z4). The zonal response patterns of benzaldehyde (Z1) and octanal (Z4) are in accordance with data that M71, expressed in Z1 is responding to benzaldehyde, and that I7, expressed in Z4 is activated by octanal (Ressler et al., 1993; Zhao et al., 1998; Bozza et al., 2002; Araneda et al., 2004). Stimulations of rat olfactory epithelium with benzaldehyde or benzene result in electro-olfactogram responses in dorsal and medial areas, respectively (Scott-Johnson et al., 2000). We show that small changes in chemical structure can result in that the OSNs responding to odorants by inducing c-Fos are located in different OR expression zones. This shift in zonal activation can be expected if the odorants activate unique ORs, as ORs are expressed in a zonally limited manner. Figure 9A illustrates zones activated in response to odorants used in paper II. The similarities between the activation of OSNs after odorant exposure and the pattern of OR expression indicate that the induction of c-Fos is a result of activation of one or a few ORs by the odorant. A change in odorant concentration influences the level of c-Fos in the OSN rather than the number of responding cells, even if concentrations several orders of magnitude above the detection threshold are used in our 38 exposure paradigm. Neither is the spatial distribution of c-Fos induction altered with increasing concentrations of odorant which indicates that additional ORs are not recruited when odorant concentration is changed. The fact that induction of c-Fos after odorant exposure exhibits the same pattern as the expression of an OR suggests that the odorant used interacts with one or a few ORs. This is not in line with the view of ORs as broadly tuned but rather points to ORs as more narrow in their response to single odorant; at least with regard to long-term cellular changes such as expression of odorant induced genes. Zonal ablation and effects on odorant detection The relatively restricted activation of OSNs after odorant observed in paper II raises the question if removal of specific OR expression zones in mice will alter the ability to detect low levels of odorants. If ORs are relatively broadly tuned then removal of one or a few of the ORs interacting with the odorant would not result in a large difference for the animal when attempting to detect an odorant. If, on the other hand, the ORs are more narrow in their response properties as our c-Fos study indicates, then removal of specific OR expression zones would affect odorant detection. Acute effects of 2,6-dichlorobenzonitrile In paper III, we have used the herbicide 2,6-dichlorobenzonitrile (dichlobenil) to selectively destroy specific areas of the olfactory epithelium. Dichlobenil has previously been shown to damage the dorsomedial part of the olfactory epithelium in mice by irreversible binding to Bowman’s glands. Metabolism of dichlobenil by a P450dependent mechanism indirectly results in degeneration of the olfactory epithelium (Brandt et al., 1990; Brittebo, 1997; Mancuso et al., 1997). Our study confirms that dichlobenil induces OSN death primarily in the 39 dorsomedial part of the epithelium. In addition we find that a low dose of dichlobenil results in ablation of areas corresponding to OR expression Z1. A higher dose results in ablation of larger areas that also involves Z2 and parts of Z3. The distribution of a Z4 OR, however, was normal even shortly after the higher dose of dichlobenil. Effects on odorant detection by removal of OR zones The spatial restriction of OR expression and the limited MRR of each OR may have consequences for the animal, e.g. in the case of localized injury as a result of physical injury or encounters to toxic substances. In order to analyze for the effects of ablation of OR expression zones on odorant detection we have analyzed mice treated with dichlobenil and compared their performance to that of untreated mice. Odorant detection thresholds of several odorants were measured using an olfactometer (see Methods section). Both treated and untreated mice readily learn to perform the task to respond correctly to the stimulus presented. Dichlobenil treated mice make as few errors as controls when analyzed for their ability to detect benzene and ethyl acetate. Dichlobenil treated mice have the same detection threshold as controls which suggests that they do not have difficulties in learning and memory, a fact previously shown in rat subjected to dichlobenil treatment as assessed in a Morris water maze spatial task (Genter et al., 1996). We show that when testing mice for detection of n-decyl alcohol and pyridazine, then dichlobenil treated mice are less sensitive than control mice. ORs activated by n-decyl alcohol are thus likely located in Z1 and Z2 as mice treated with a high dose are almost anosmic to the odorant while mice treated with a low dose still can detect n-decyl alcohol although a higher concentration is needed than for control mice. Pyridazine on the other hand is detected by ORs located in Z1 as well as ORs in Z3 and/or Z4 since dichlobenil treatment results in an increase in detection threshold but does not result in a severe inability to detect pyridazine. The detection threshold for benzene and ethyl acetate were not affected by removal of Z1 and Z2, suggesting that detection at threshold concentrations of these odorants occur in Z3 40 and/or Z4. Figure 9B shows probable location of high affinity ORs for the odorants tested in the olfactometer. Zonally restricted activation of OSNs We have, in two independent studies, shown that activation of OSNs by odorants is mediated by a limited number of ORs in vivo. OSNs responding to odorants are located in all four zones where no zone appears to display higher sensitivity to odorants in general. Our studies show that there may be substantial differences between zones in their ability to mediate detection of the same odorant. Although ORs are Figure 9. (A) Summary of zones containing OSNs activated by different odorants as observed by induction of c-Fos. The location of identified ORs for benzaldehyde (M71) and octanal (I7) is located in Z1 and Z4 respectively. (B) Location of high affinity ORs as determined by studies on detection threshold after zone-specific removal of OSNs. 41 broadly tuned in heterologous systems it appears that ORs with lower affinity for a given odorant may not be able to compensate for the loss of high affinity ORs when attempting to detect odorants at low concentrations. 42 Regional differences in maintenance of the olfactory epithelium (papers III and IV) The life-long function of the olfactory system is dependent on continuous production of new OSNs to replace damaged or worn-out neurons. The OSN progenitors, the GBCs, reside in the basal compartment of the olfactory epithelium. This is most readily observed when the olfactory epithelium is physically damaged as an increase in proliferation of the GBCs (e.g. (Schwartz Levey et al., 1991; Suzuki & Takeda, 1991)). The immature cells produced by the GBCs mature into OSNs to replace dying neurons with functional neurons able to respond to olfactory cues. The studies made on proliferation in the rodent olfactory epithelia have not taken the different zonal identities or differences in milieu between the zones into account. One may hypothesize that the intricate structure of the turbinates protruding into the nasal lumen result in differences in the milieu for the neurons and hence differences in proliferation and survival. We have examined proliferation in the mouse olfactory epithelium by utilizing bromodeoxyuridine incorporation in paper IV. We demonstrate that the basal cell proliferation is uneven over the epithelial sheet in the adult and that Z1 has a lower rate of proliferation compared to Z2-4. The ratio of mature and immature OSNs is not the same in Z1 as in Z4. Z1 has fewer immature neurons compared to Z4 in adult mice. The number of immature neurons is in agreement with the lower proliferation in Z1 compared to Z4 at this age. The epithelium is however thicker in Z1 and the major part of the Z1 neuronal layer is composed of mature, OMP positive OSNs. The more mature nature of Z1 indicates that OSNs are retained longer in the epithelium and pulsechase studies show that the newly born OSNs survive longer in Z1 compared to more lateral areas. RA has been suggested to promote survival of mature OSNs and to decrease proliferation of progenitor cells. The rate-limiting enzyme in RA production, RALDH2, is expressed in a low level in Z1 and a high level in Z4 (paper I) which 43 does not correlate with the high level of proliferation in lateral zones or the shorter life-span of Z4 OSNs. Intriguingly, the adult pattern of proliferation in the olfactory epithelium appears at a time period where olfactory ensheating cells acquire their adult proliferation rate and the start of elimination of mistargeted axons occurs (e.g. (Zou et al., 2004; Watanabe et al., 2006)). It can be speculated that ongoing neurogenesis is an advantage for an animal in a changing environment where birth of new sensory cells may help the olfactory system to regulate the sensitivity to new odorants. It has previously been shown that mice exposed to certain odorants can become more sensitive to these odorants as determined by a behavioral assay (Wang et al., 1993). OSNs that are activated have been suggested to have a strong survival advantage over non-activated OSNs (e.g. (Zhao & Reed, 2001)). It is thus plausible that activated OSN are selected for in the olfactory epithelium. By exposing young mice to odorants we find that newly produced OSNs responding to the specific odorants are not enriched in the epithelium, suggesting that the sensitization to odorant occurs at the level of the individual cell e.g. by up-regulation of components of the signaling transduction cascade. The period of odorant exposure was approximately three weeks which corresponds to a time period during which OSNs in Z4 are turned over but Z1 neurons have slower turn over times. It is thus still possible that a longer exposure period will result in selection of responding OSNs in Z1. Regeneration after dichlobenil treatment In paper III we show that parts of the olfactory epithelium are able to regenerate after dichlobenil treatment during a time period of a few weeks. Some areas do however not recover even after 8 months. The non-repopulated areas correspond to the areas expressing ORs specific for the most dorsomedial zone, Z1, as shown using in situ hybridization. The damage to Z2 and the following regeneration as well as the lack of regeneration in Z1 suggest that there may be differences between the 44 most dorsomedial zone and the other zones with regard to ability of cells to respond to the damage and to repopulate the epithelium. The inability of Z1 to regenerate after dichlobenil is not an effect of lack of proliferation in Z1 in the adult. Z1 has a small number of proliferating cells that in time should be able to repopulate the epithelium. It is instead possible that cells responsive for repopulation are missing in Z1 after dichlobenil treatment. Mammalian achaete-scute homologue 1 (Mash1) is expressed in a subpopulation of proliferative cells of the olfactory epithelium (Cau et al., 1997). Mice lacking a functional Mash1 gene almost completely lack OSNs which suggests that Mash1 is necessary for OSN formation (Guillemot et al., 1993). When analyzing for Mash1 expression in dichlobenil treated mice by in situ hybridization, we have observed that areas not recovering from dichlobenil treatment are devoid of Mash1 positive cells. Cells expressing Mash1 are however present in Z2 (fig 10). It is not known whether the cells that repopulate the epithelium in e.g. Z2 are spared from the toxic effects of dichlobenil. It is not unlikely that cells from more lateral, undamaged areas migrate to repopulate Figure 10. A population of cells located in the basal part of the epithelium is positive for Mash1 as seen by in situ hybridization. The area marked by the dotted line is devoid of Mash1 positive cells and is unable to regenerate after dichlobenil treatment. The arrow heads point to Mash1 expressing cells. 45 areas that regenerate but that these fail to migrate over the Z1/2 border. Cells from the Z2-4 areas may not be able to survive in the region of Z1 OR expression as the sharp boundary between regenerated and nonregenerated areas indicates that there are differences in local signals/milieu between Z1 and the other zones. During regeneration of the epithelium some ORs appear to be expressed in spatially expanded regions compared to control mice. This can be seen in fig 11 as an expansion of Z3 and Z4 ORs up to the Z1/2 border as assessed by in situ hybridization using OR probes. This is evident early during the regeneration process after a high dose of dichlobenil. Figure 11. The distribution of OR expression in Z3 and Z4 is expanded in epithelium of mice treated with dichlobenil as seen with radioactive in situ hybridization using OR probes, signal in white. Lines demarcate the Z1/Z2 border and the dorsal extent of Z4, respectively. 46 The graded expression of genes with possible instructive roles for OSNs (paper I) may be perturbed early during the regeneration process leading to expanded regions of OR expression. Later, however, the normal pattern of OR expression in Z2-4 is restored. OSNs expressing ORs outside their normal expression area likely project to ectopic glomeruli that in time are eliminated, aided by sensory experience (Nakatani et al., 2003; St John & Key, 2003; Kerr & Belluscio, 2006). 47 The olfactory epithelium can be divided into two major domains Throughout our studies we noticed differences between epithelial Z1 and the other zones with respect to gene expression (paper I), effect of a toxic compound (paper III) and tissue homeostasis (paper IV). In addition we describe an overlap of OR expression between Z2-4 suggesting that these zones are not strictly separated from each other (paper I). I have in fig 12 illustrated this by listing some of our findings together with findings made by others. Genes expressed in all cell layers of the epithelium, as well in the mesechyme, can display expression areas limited by the Z1/Z2 border. Such genes include e.g. metabolizing enzymes, axon guidance molecules and transcription factors (Miyawaki et al., 1996; Alenius & Bohm, 1997; Yoshihara et al., 1997; Norlin & Berghard, 2001; Oka et al., 2003; Gussing & Bohm, 2004). It is likely that life-span and maturity of the OSNs in the olfactory epithelium follow the same zonal pattern. About 50% of all ORs are expressed in Z1 (Zhang et al., 2004). Combined with the fact that Z1 has a higher number of mature OSNs compared to more lateral zones suggests greater diversity of OSNs located in Z1. The large number of differences between Z1 and the other zones make it likely that there is a division of the olfactory epithelium into two major domains and this may have a functional role. 48 Figure 12. Z1 is different from the other zones in regard to expression of ORs and other genes, response to toxic compound and tissue homeostasis indicating that Z1 might be inherently different from the other zones. The location of e.g. gene expression is color coded as insert. 49 CONCLUSIONS ORs, as well as other genes, display regional limitations of expression in the olfactory epithelium. The zonally restricted pattern of expression exists in all cell types of the olfactory epithelium of the mouse. Reasons for the zonal anatomy are still elusive although the answer is hopefully not far ahead. The adult pattern of OR expression is apparent from the time point where spatial OR expression are first evident in the embryo. The expression of Z2-4 ORs is overlapping while Z1 ORs appear not to overlap with ORs from other zones. Zonal arrangement must be maintained throughout life as there is continuous neurogenesis and gradients of expression of genes with instructive roles may be involved in specifying OSNs and thus directly or indirectly be important for correct zonal OR expression. An odorant is detected by limited numbers of ORs as suggested by the induction of the activity marker c-Fos after odorant exposure. In the adult mice localized injury may result in specific inability to detect odorants at their normal detection threshold concentration and thus ORs with lower affinity are not able to compensate for the lack of high affinity ORs. The damaged epithelium is, after regeneration, displaying the same zonally limited expression of Z2, Z3 and Z4 ORs as before damage. This may be a result of gradients of instructive gene expression that is observed in the olfactory epithelium. It can be concluded that Z1 OSNs are different from Z2-4 with regard to toxic responses, gene expression, proliferation and OSN survival which may open for different functions of Z1 and Z2-4. 50 ACKNOWLEDGEMENTS 51 REFERENSES Alenius, M. and Bohm, S. (1997). "Identification of a novel neural cell adhesion molecule-related gene with a potential role in selective axonal projection." J Biol Chem 272: 26083-26086. Amoore, J. E. (1982). "Odor theory and odor classification." Fragance Chemistry Academic Press, Inc.: pp 1-76. Araneda, R., Peterlin, Z., Zhang, X., Chesler, A. and Firestein, S. (2004). "A pharmacological profile of the aldehyde receptor repertoire in rat olfactory epithelium." J Physiol 555: 743-56. Araneda, R. C., Kini, A. D. and Firestein, S. (2000). "The molecular receptive range of an odorant receptor." Nature Neurosci 3: 1248-1255. Asson-Batres, M., Zeng, M., Savchenko, V., Aderoju, A. and McKanna, J. (2003). "Vitamin A deficiency leads to increased cell proliferation in olfactory epithelium of mature rats." J Neurobiol 54: 539-54. Aungst, J. L., Heyward, P. M., Puche, A. C., Karnup, S. V., Hayar, A., Szabo, G. and Shipley, M. T. (2003). "Centre-surround inhibition among olfactory bulb glomeruli." Nature 426: 623-629. Bakalyar, H. A. and Reed, R. R. (1990). "Identification of a specialized adenylate cyclase that may mediate odorant detection." Science 250: 14031406. Balasubramanian, S., Lynch, J. W. and Barry, P. H. (1995). "The permeation of organic cations through cAMP-gated channels in mammalian olfactory receptor neurons." J Membr Biol 146: 177-91. Belluscio, L., Gold, G. H., Nemes, A. and Axel, R. (1998). "Mice deficient in Golf are anosmic." Neuron 20: 69-81. 52 Bodyak, N. and Slotnick, B. (1999). "Performance of mice in an automated olfactometer: odor detection, discrimination and odor memory." Chem Senses 24: 637-645. Boekhoff, I., Inglese, J., Schleicher, S., Koch, W. J., Lefkowitz, R. J. and Breer, H. (1994). "Olfactory desensitization requires membrane targeting of receptor kinase mediated by βγ-subunits of heterotrimeric G proteins." J Biol Chem 269: 37-40. Borisy, F. F., Ronnett, G. V., Cunningham, A. M., Juilfs, D., Beavo, J. and Snyder, S. H. (1992). "Calcium/calmodulin-activated phosphodiesterase expressed in olfactory receptor neurons." J Neuroscience 12: 915-923. Bozza, T., Feinstein, P., Zheng, C. and Kauer, J. S. (2002). "Odorant receptor expression defines functional units in the mouse olfactory system." J Neurosci 22: 3033-3043. Bradley, J., Bonigk, W., Yau, K.-W. and Frings, S. (2004). "Calmodulin permanently associates with rat olfactory CNG channels under native conditions." Nat Neurosci 7: 705-710. Bradley, J., Li, J., Davidson, N., Lester, H. A. and Zinn, K. (1994). "Heteromeric olfactory cyclic nucleotide-gated channels: a subunit that confers increased sensitivity to cAMP." Proc Natl Acad Sci 91: 8890-8894. Bradley, J., Reuter, D. and Frings, S. (2001). "Facilitation of CalmodulinMediated Odor Adaptation by cAMP-Gated Channel Subunits." Science 294: 2176-2178. Brandt, I., Brittebo, E. B., Feil, V. J. and Bakke, J. E. (1990). "Irreversible binding and toxicity of the herbicide dichlobenil (2,6-dichlorobenzonitrile) in the olfactory mucosa of mice." Toxicol Appl Pharmacol 103: 491-501. Brittebo, E. B. (1997). "Metabolism-dependent activation and toxicity of chemicals in nasal glands." Mutat Res 380: 61-75. 53 Brunet, L. J., Gold, G. H. and Ngai, J. (1996). "General anosmia caused by a targeted disruption of the mouse olfactory cyclic nucleotide-gated cation channel." Neuron 17: 681-693. Buck, L. and Axel, R. (1991). "A novel multigene family may encode odorant receptors: a molecular basis for odor recognition." Cell 65: 175-187. Caggiano, M., Kauer, J. S. and Hunter, D. D. (1994). "Globose basal cells are neuronal progenitors in the olfactory epithelium: a linage analysis using a replication-incompetent retrovirus." Neuron 13: 339-352. Calof, A. L. and Chikaraishi, D. M. (1989). "Analysis of neurogenesis in a mammalian neuro-epithelium: proliferation and differentiation of an olfactory neuron precursor in vitro." Neuron 3: 115-127. Camoletto, P., Colesanti, A., Ozon, S., Sobel, A. and Fasolo, A. (2001). "Expression of stathmin and SCG10 proteins in the olfactory neurogenesis during development and after lesion in the adulthood." Brain Res Bull 54: 1928. Carter, L. A., MacDonald, J. L. and Roskams, A. J. (2004). "Olfactory Horizontal Basal Cells Demonstrate a Conserved Multipotent Progenitor Phenotype." J Neurosci 24: 5670-5683. Cattarelli, M., Astic, L. and Kauer, J. S. (1988). "Metabolic mapping of 2deoxyglucose uptake in the rat piriform cortex using computerized image processing." Brain Research 442: 180-184. Cau, E., Gradwohl, G., Fode, C. and Guillemot, F. (1997). "Mash1 activates a cascade of bHLH regulators in olfactory neuron progenitors." Development 124: 1611-21. Chen, T.-Y. and Yau, K.-W. (1994). "Direct modulation by Ca2+-calmodulin of cyclic nucleotide-activated channel of rat olfactory receptor neurons." Nature 368: 545-548. 54 Chen, X., Fang, H. and Schwob, J. E. (2004). "Multipotency of purified, transplanted globose basal cells in olfactory epithelium." J Comp Neurol 469: 457-74. Chen, Y., Getchell, M. L., Ding, X. and Getchell, T. V. (1992). "Immunolocalization of two cytochrome P450 isozymes in rat nasal chemosensory tissue." Neuroreport 3: 749-52. Chess, A., Simon, I., Cedar, H. and Axel, R. (1994). "Allelic inactivation regulates olfactory receptor gene expression." Cell 78: 823-834. Clevenger, A. C. and Restrepo, D. (2006). "Evaluation of the Validity of a Maximum Likelihood Adaptive Staircase Procedure for Measurement of Olfactory Detection Threshold in Mice." Chem Senses 31: 9-26. Couto, A., Alenius, M. and Dickson, B. J. (2005). "Molecular, Anatomical, and Functional Organization of the Drosophila Olfactory System." Current Biology 15: 1535-1547. Ding, X. X. and Coon, M. J. (1988). "Purification and characterization of two unique forms of cytochrome P-450 from rabbit nasal microsomes." Biochemistry 27: 8330-7. Dzeja, C., Hagen, V., Kaupp, U. B. and Frings, S. (1999). "Ca2+ permeation in cyclic nucleotide-gated channels." EMBO J 18: 131-44. Eggan, K., Baldwin, K., Tackett, M., Osborne, J., Gogos, J., Chess, A., Axel, R. and Jaenisch, R. (2004). "Mice cloned from olfactory sensory neurons." Nature 428: 44-9. Elsaesser, R. and Paysan, J. (2005). "Morituri te salutant? Olfactory signal transduction and the role of phosphoinositides." J Neurocytol 34: 97-116. Fan, J. and Ngai, J. (2001). "Onset of odorant receptor gene expression during olfactory sensory neuron regeneration." Dev Biol 229: 119-127. 55 Farbman, A. I. and Margolis, F. L. (1980). "Olfactory marker protein during ontogeny: immunohistochemical localization." Dev Biol 74: 205-215. Fried, H. U., Fuss, S. H. and Korsching, S. I. (2002). "Selective imaging of presynaptic activity in the mouse olfactory bulb shows concentration and structure dependence of odor responses in identified glomeruli." Proc Natl Acad Sci U S A 99: 3222-3227. Genter, M. B., Owens, D. M., Carlone, H. B. and Crofton, K. M. (1996). "Characterization of olfactory deficits in the rat following administration of 2,6-dichlorobenzonitrile (dichlobenil), 3,3'-iminodipropionitrile, or methimazole." Fund Appl Toxicol 29: 71-77. Glusman, G., Yanai, I., Rubin, I. and Lancet, D. (2001). "The complete human olfactory subgenome." Genome Res 11: 685-702. Godfrey, P. A., Malnic, B. and Buck, L. B. (2004). "The mouse olfactory receptor gene family." Proc Natl Acad Sci USA 101: 2156-2161. Gordon, M. K., Mumm, J. S., Davis, R. A., Holcomb, J. D. and Calof, A. L. (1995). "Dynamics of MASH1 expression in vitro and in vivo suggest a nonstem cell site of MASH1 action in the olfactory receptor neuron lineage." Mol Cell Neurosci 6: 363-79. Graziadei, P. P. and Graziadei, G. A. (1979). "Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons." J Neurocytol 8: 1-18. Griff, I. C. and Reed, R. R. (1995). "The genetic basis for specific anosmia to isovaleric acid in the mouse." Cell 83: 407-14. Guillemot, F., Lo, L.-C., Johnson, J. E., Auerbach, A., Anderson, D. J. and Joyner, A. L. (1993). "Mammalian achaete-scute homolog 1 is required for the early development of olfactory and autonomic neurons." Cell 75: 463-476. 56 Gussing, F. and Bohm, S. (2004). "NQO1 activity in the main and the accessory olfactory systems correlates with the zonal topography of projection maps." Eur J Neurosci 19: 2511-8. Haberly, L. B. (1969). "Single unit responses to odor in the prepyriform cortex of the rat." Brain Res 12: 481-4. Herdegen, T. and Leah, J. D. (1998). "Inducible and constitutive transcription factors in the mammalian nervous system: control of gene expression by jun, fos and krox, and CREB/ATF proteins." Brain Res Rev 28: 370-490. Hinds, J. W., Hinds, P. L. and McNelly, N. A. (1984). "An autoradiographic study of the mouse olfactory epithelium: evidence for long-lived receptors." Anat Rec 210: 375-83. Holbrook, E. H., Szumowski, K. E. and Schwob, J. E. (1995). "An immunochemical, ultrastructural, and developmental characterization of the horizontal basal cells of rat olfactory epithelium." J Comp Neurol 363: 12946. Hoppe, R., Breer, H. and Strotmann, J. (2006). "Promoter motifs of olfactory receptor genes expressed in distinct topographic patterns." Genomics 87: 71123. Horowitz, L. F., Montmayeur, J. P., Echelard, Y. and Buck, L. B. (1999). "A genetic approach to trace neural circuits." Proc Natl Acad Sci U S A 96: 1949. Huard, J. M. and Schwob, J. E. (1995). "Cell cycle of globose basal cells in rat olfactory epithelium." Dev Dyn 203: 17-26. Huard, J. M. T., Youngentob, S. L., Goldstein, B. J., Luskin, M. B. and Schwob, J. E. (1998). "Adult olfactory epithelium contains multipotent progenitors that give rise to neurons and non-neuronal cells." J Comp Neurol 400: 469-486. 57 Hägglund, M., Berghard, A., Strotmann, J. and Bohm, S. (2006). "Retinoic acid receptor-dependent survival of olfactory sensory neurons in postnatal and adult mice." J Neurosci 26: 3281-91. Isaacson, J. S. and Strowbridge, B. W. (1998). "Olfactory reciprocal synapses: dendritic signaling in the CNS." Neuron 20: 749-761. Iwema, C. L., Fang, H., Kurtz, D. B., Youngentob, S. L. and Schwob, J. E. (2004). "Odorant Receptor Expression Patterns Are Restored in LesionRecovered Rat Olfactory Epithelium." J Neurosci 24: 356-369. Jones, D. T. and Reed, R. R. (1989). "Golf: an olfactory neuron specific Gprotein involved in odorant signal transduction." Science 244: 790-795. Juilfs, D. M., Fülle, H.-J., Zhao, A. Z., Houslay, M. D., Garbers, D. L. and Beavo, J. A. (1997). "A subset of olfactory neurons that selectively express cGMP-stimulated phosphodiesterase (PDE2) and guanylyl cyclase-D define a unique olfactory signal transduction pathway." Proc Natl Acad Sci 94: 33883395. Kajiya, K., Inaki, K., Tanaka, M., Haga, T., Kataoka, H. and Touhara, K. (2001). "Molecular bases of odor discrimination: reconstitution of olfactory receptors that recognize overlapping sets of odorants." J Neurosci 21: 60186025. Kaplan, M. S., McNelly, N. A. and Hinds, J. W. (1985). "Population dynamics of adult-formed granule neurons of the rat olfactory bulb." J Comp Neurol 239: 117-25. Katada, S., Hirokawa, T., Oka, Y., Suwa, M. and Touhara, K. (2005). "Structural basis for a broad but selective ligand spectrum of a mouse olfactory receptor: mapping the odorant-binding site." J Neurosci 25: 180615. Kaupp, U. B., Niidome, T., Tanabe, T., Terada, S., Bonigk, W., Stuhmer, W., Cook, N. J., Kangawa, K., Matsuo, H., Hirose, T., Miyata, T. and Numa, S. (1989). "Primary structure and functional expression from complementary 58 DNA of the rod photoreceptor cyclic GMP-gated channel." Nature 342: 762766. Kaupp, U. B. and Seifert, R. (2002). "Cyclic Nucleotide-Gated Ion Channels." Physiol Rev 82: 769-824. Kelliher, K. R., Ziesmann, J., Munger, S. D., Reed, R. R. and Zufall, F. (2003). "Importance of the CNGA4 channel gene for odor discrimination and adaptation in behaving mice." Proc Natl Acad Sci 100: 4299-4304. Kerr, M. A. and Belluscio, L. (2006). "Olfactory experience accelerates glomerular refinement in the mammalian olfactory bulb." Nat Neurosci 9: 484-486. Kimbell, J. S., Godo, M. N., Gross, E. A., Joyner, D. R., Richardson, R. B. and Morgan, K. T. (1997). "Computer Simulation of Inspiratory Airflow in All Regions of the F344 Rat Nasal Passages." Toxicology and Applied Pharmacology 145: 388-398. Kleene, S. J. (1993). "Origin of the chloride current in olfactory transduction." Neuron 11: 123-132. Kosaka, K. and Kosaka, T. (2005). "synaptic organization of the glomerulus in the main olfactory bulb: compartments of the glomerulus and heterogeneity of the periglomerular cells." Anat Sci Int 80: 80-90. Kubick, S., Strotmann, J., Andreini, I. and Breer, H. (1997). "Subfamily of Olfactory Receptors Characterized by Unique Structural Features and Expression Patterns." J Neurochem 69: 465-475. Kurahashi, T. and Menini, A. (1997). "Mechanism of odorant adaptation in the olfactory receptor cell." Nature 385: 725-9. LaMantia, A.-S., Bhasin, N., Rhodes, K. and Heemskerk, J. (2000). "Mesenchymal/epithelial induction mediates olfactory pathway formation." Neuron 28: 411-425. 59 Leinders-Zufall, T., Ma, M. and Zufall, F. (1999). "Impaired odor adaptation in olfactory receptor neurons after inhibition of Ca2+/calmodulin kinase II." J Neurosci 19: RC19 (1-6). Leinders-Zufall, T., Rand, M. N., Shepherd, G. M., Greer, C. A. and Zufall, F. (1997). "Calcium entry through cyclic nucleotide-gated channels in individual cilia of olfactory receptor cells: spatiotemporal dynamics." J Neurosci 17: 4136-48. Lewcock, J. W. and Reed, R. R. (2004). "A feedback mechanism regulates monoallelic odorant receptor expression." Proc Natl Acad Sci U S A 101: 1069-1074. Li, J., Ishii, T., Feinstein, P. and Mombaerts, P. (2004). "Odorant receptor gene choice is reset by nuclear transfer from mouse olfactory sensory neurons." Nature 428: 393-399. Li, Y., Field, P. M. and Raisman, G. (2005). "Olfactory ensheathing cells and olfactory nerve fibroblasts maintain continuous open channels for regrowth of olfactory nerve fibres." Glia 52: 245-251. Liman, E. R. and Buck, L. B. (1994). "A second subunit of the olfactory cyclic nucleotide-gated channel confers high sensitivity to cAMP." Neuron 13: 611621. Lin, D. Y., Shea, S. D. and Katz, L. C. (2006). "Representation of Natural Stimuli in the Rodent Main Olfactory Bulb." Neuron 50: 937-949. Lin, W., Arellano, J., Slotnick, B. and Restrepo, D. (2004). "Odors detected by mice deficient in cyclic nucleotide-gated channel subunit A2 stimulate the main olfactory system." J Neurosci 24: 3703-3710. Liu, D. T., Tibbs, G. R. and Siegelbaum, S. A. (1996). "Subunit Stoichiometry of Cyclic Nucleotide-Gated Channels and Effects of Subunit Order on Channel Function." Neuron 16: 983-990. 60 Lomvardas, S., Barnea, G., Pisapia, D. J., Mendelsohn, M., Kirkland, J. and Axel, R. (2006). "Interchromosomal Interactions and Olfactory Receptor Choice." Cell 126: 403-413. Ma, M. and Shepherd, G. M. (2000). "Functional mosaic organization of mouse olfactory receptor neurons." Proc Natl Acad Sci 97: 12869-12874. Mackay-Sim, A. and Kittel, P. (1991). "Cell dynamics in the adult mouse olfactory epithelium: a quantitative autoradiographic study." J Neurosci 11: 979-984. Malnic, B., Godfrey, P. A. and Buck, L. B. (2004). "The human olfactory receptor gene family." Proc Natl Acad Sci U S A 101: 2584-9. Malnic, B., Hirono, J., Sato, T. and Buck, L. B. (1999). "Combinatorial receptor codes for odors." Cell 96: 713-723. Mancuso, M., Giovanetti, A. and Brittebo, E. B. (1997). "Effects of dichlobenil on ultrastructural morphology and cell replication in the mouse olfactory mucosa." Tox Pathol 25: 186-194. Margrie, T. W., Sakmann, B. and Urban, N. N. (2001). "Action potential propagation in mitral cell lateral dendrites is decremental and controls recurrent and lateral inhibition in the mammalian olfactory bulb." Proc Natl Acad Sci U S A 98: 319-324. Matulionis, D. H. (1975). "Ultrastructural study of mouse olfactory epithelium following destruction by ZnSO4 and its subsequent regeneration." Am J Anat 142: 67-89. Menco, B. P. M. (1980). "Qualitative and quantitative freeze-fracture studies on olfactory and nasal respiratory epithelial surfaces of frog, ox, rat, and dog III. Tight-junctions." Cell Tissue Res 211: 361-373. Mendoza, A. S. (1993). "Morphological studies on the rodent main and accessory olfactory systems: the regio olfactoria and vomeronasal organ." Ann Anat 175: 425-446. 61 Meyer, M. R., Angele, A., Kremmer, E., Kaupp, B. and Muller, F. (2000). "A cGMP-signaling pathway in a subset of olfactory sensory neurons." Proc Natl Acad Sci 97: 10595-10600. Mic, F., Haselbeck, R., Cuenca, A. and Duester, G. (2002). "Novel retinoic acid generating activities in the neural tube and heart identified by conditional rescue of Raldh2 null mutant mice." Development 129: 2271-82. Miragall, F., Krause, D., de Vries, U. and Dermietzel, R. (1994). "Expression of the tight junction protein ZO-1 in the olfactory system: presence of ZO-1 on olfactory sensory neurons and glial cells." J Comp Neurol 341: 433-48. Miyamichi, K., Serizawa, S., Kimura, H. M. and Sakano, H. (2005). "Continuous and Overlapping Expression Domains of Odorant Receptor Genes in the Olfactory Epithelium Determine the Dorsal/Ventral Positioning of Glomeruli in the Olfactory Bulb." J Neurosci 25: 3586-3592. Miyawaki, A., Homma, H., Tamura, H., Matsui, M. and Mikoshiba, K. (1996). "Zonal distribution of sulfotransferase for phenol in olfactory sustentacular cells." EMBO J 15: 2050-2055. Moon, C., Simpson, P. J., Tu, Y., Cho, H. and Ronnett, G. V. (2005). "Regulation of intracellular cyclic GMP levels in olfactory sensory neurons." J Neurochem 95: 200-209. Mori, K., Nagao, H. and Yoshihara, Y. (1999). "The olfactory bulb: coding and processing of odor molecule information." Science 286: 711-715. Mori, K., Takahashi, Y. K., Igarashi, K. M. and Yamaguchi, M. (2006). "Maps of Odorant Molecular Features in the Mammalian Olfactory Bulb." 86: 409433. Munger, S. D., Lane, A. P., Zhong, H., Leinders-Zufall, T., Yau, K.-W., Zufall, F. and Reed, R. R. (2001). "Central role of the CNGA4 channel subunit in Ca2+-calmodulin-dependent odor adaptation." Science 294: 21722175. 62 Nakamura, T. and Gold, G. H. (1987). "A cyclic nucleotide-gated conductance in olfactory receptor cilia." Nature 325: 442-444. Nakatani, H., Serizawa, S., Nakajima, T., Imai, T. and Sakano, H. (2003). "Developmental elimination of ectopic projection sites for the transgenic OR gene that has lost zone specificity in the olfactory epithelium." Eur J Biochem 18: 2425-2432. Neuhaus, E. M., Mashukova, A., Zhang, W., Barbour, J. and Hatt, H. (2006). "A Specific Heat Shock Protein Enhances the Expression of Mammalian Olfactory Receptor Proteins." Chem Senses 31: 445-452. Ngai, J., Chess, A., Dowling, M. M., Necles, N., Macagno, E. R. and Axel, R. (1993a). "Coding of olfactory information: topography of odorant receptor expression in the catfish olfactory epithelium." Cell 72: 667-680. Ngai, J., Dowling, M. M., Buck, L., Axel, R. and Chess, A. (1993b). "The family of genes encoding odorant receptors in the channel catfish." Cell 72: 657-666. Nomura, T., Takahashi, S. and Ushiki, T. (2004). "Cytoarchitecture of the normal rat olfactory epithelium: Light and scanning electron microscopic studies." Arch Histol Cytol 67: 159-170. Norlin, E. M. and Berghard, A. (2001). "Spatially restricted expression of regulators of G-protein signaling (RGS) in primary olfactory neurons." Mol Cell Neurosci 17: 872-882. Oka, Y., Kobayakawa, K., Nishizumi, H., Miyamichi, K., Hirose, S., Tsuboi, A. and Sakano, H. (2003). "O-MACS, a novel member of the medium-chain acyl-CoA synthetase family, specifically expressed in the olfactory epithelium in a zone-specific manner." Eur J Biochem 270: 1995-2004. Pace, U., Hanski, E., Salomon, Y. and Lancet, D. (1985). "Odorant-sensitive adenylate cyclase may mediate olfactory reception." Nature 316: 255-258. 63 Peppel, K., Boekhoff, I., McDonald, P., Breer, H., Caron, M. G. and Lefkowitz, R. J. (1997). "G protein-coupled receptor kinase 3 (GRK3) gene disruption leads to loss of odorant receptor desensitization." J Biol Chem 272: 25425-25428. Pilpel, Y. and Lancet, D. (1999). "The variable and conserved interfaces of modeled olfactory receptor proteins." Protein Sci 8: 969-977. Qasba, P. and Reed, R. R. (1998). "Tissue and zonal-specific expression of an olfactory receptor transgene." J Neurosci 18: 227-236. Rall, W., Shepherd, G. M., Reese, T. S. and Brightman, M. W. (1966). "Dendrodendritic synaptic pathway for inhibition in the olfactory bulb." Experimental Neurology 14: 44-56. Rash, J., Davidson, K., Kamasawa, N., Yasumura, T., Kamasawa, M., Zhang, C., Michaels, R., Restrepo, D., Ottersen, O., Olson, C. and Nagy, J. (2005). "Ultrastructural localization of connexins (Cx36, Cx43, Cx45), glutamate receptors and aquaporin-4 in rodent olfactory mucosa, olfactory nerve and olfactory bulb." J Neurocytol 35: 307-41. Rawson, N. E., Eberwine, J., Dotson, R., Jackson, J., Ulrich, P. and Restrepo, D. (2000). "Expression of mRNAs encoding for two different olfactory receptors in a subset of olfactory receptor neurons." J Neurochem 75: 185195. Reisert, J., Bauer, P. J., Yau, K. W. and Frings, S. (2003). "The Ca-activated Cl channel and its control in rat olfactory receptor neurons." J Gen Physiol 122: 349-63. Ressler, K. J., Sullivan, S. L. and Buck, L. B. (1993). "A zonal organization of odorant receptor gene expression in the olfactory epithelium." Cell 73: 597609. Ressler, K. J., Sullivan, S. L. and Buck, L. B. (1994). "Information coding in the olfactory system: evidence for a stereotyped and highly organized epitope map in the olfactory bulb." Cell 79: 1245-1256. 64 Reuter, D., Zierold, K., Schröder, W. H. and Frings, S. (1998). "A depolarizing chloride current contributes to chemoelectrical transduction in olfactory sensory neurons in situ." J Neurosci 18: 6623-6630. Rothman, A., Feinstein, P., Hirota, J. and Mombaerts, P. (2005). "The promoter of the mouse odorant receptor gene M71." Mol Cell Neurosci 28: 535-46. Royet, J. P., Jourdan, F., Ploye, H. and Souchier, C. (1989). "Morphometric modifications associated with early sensory experience in the rat olfactory bulb: II. Stereological study of the population of olfactory glomeruli." J Comp Neurobiol 289: 594-609. Rubin, B. D. and Katz, L. C. (1999). "Optical imaging of odorant representations in the mammalian olfactory bulb." Neuron 23: 499-511. Saito, H., Kubota, M., Roberts, R. W., Chi, Q. and Matsunami, H. (2004). "RTP family members induce functional expression of mammalian odorant receptors." Cell 119: 679-691. Sato, T., Hirono, J., Tonoike, M. and Takebayashi, M. (1994). "Tuning specificities to aliphatic odorants in mouse olfactory receptor neurons and their local distribution." J Neurophysiol 72: 2980-2989. Schoenfeld, T. and Cleland, T. (2005). "The anatomical logic of smell." Trends Neurosci. 28: 620-7. Schoppa, N. and Urban, N. (2003). "Dendritic processing within olfactory bulb circuits." Trends Neurosci 26: 501-6. Schwartz Levey, M., Chikaraishi, D. M. and Kauer, J. S. (1991). "Characterization of potential precursor populations in the mouse olfactory epithelium using immunohistochemistry and auroradiography." J Neurosci 11: 3556-3564. 65 Schwob, J. E. (2002). "Neural regeneration and the peripheral olfactory system." Anat Rec 269: 33-49. Schwob, J. E., Youngentob, S. L. and Mezza, R. C. (1995). "Reconstitution of the rat olfactory epithelium after methyl bromide-induced lesion." J Comp Neurol 359: 15-37. Scott-Johnson, P., Blakley, D. and Scott, J. (2000). "Effects of air flow on rat electroolfactogram." Chem Senses. 25: 761-8. . Scott, J. W., Brierley, T. and Schmidt, F. H. (2000). "Chemical determinants of the rat electro-olfactogram." J. Neurosci. 20: 4721-4731. Serizawa, S., Ishii, T., Nakatani, H., Tsuboi, A., Nagawa, F., Asano, M., Sudo, K., Sagakami, J., Sakano, H., Ijiri, T., Matsuda, Y., Suzuki, M., Yamamori, T., Iwakura, Y. and Sakano, H. (2000). "Mutually exclusive expression of odorant receptor transgenes." Nature Neurosci 3: 687-693. Serizawa, S., Miyamichi, K., Nakatani, H., Suzuki, M., Saito, M., Yoshihara, Y. and Sakano, H. (2003). "Negative Feedback Regulation Ensures the One Receptor-One Olfactory Neuron Rule in Mouse." Science 302: 2088-2094. Shen, R., Chen, Y., Huang, L., Vitale, E. and Solursh, M. (1994). "Characterization of the human MSX-1 promoter and an enhancer responsible for retinoic acid induction." Cell Mol Biol Res 40: 297-312. Singer, M. S. and Shepherd, G. M. (1994). "Molecular modeling of ligandreceptor interactions in the OR5 olfactory receptor." Neuroreport 5: 12971300. Slotnick, B. and Bodyak, N. (2002). "Odor discrimination and odor quality perception in rats with disruption of connections between the olfactory epithelium and olfactory bulb." J Neurosci 22: 4205-4216. Slotnick, B., Cockerham, R. and Pickett, E. (2004). "Olfaction in olfactory bulbectomized rats." J Neurosci 24: 9195-200. 66 Song, Y., Hui, J., Fu, K. and Richman, J. (2004). "Control of retinoic acid synthesis and FGF expression in the nasal pit is required to pattern the craniofacial skeleton." Dev Biol 276: 313-29. Sosinsky, A., Glusman, G. and Lancet, D. (2000). "The genomic structure of human olfactory receptor genes." Genomics 70: 49-61. St John, J. A. and Key, B. (2003). "Axon mis-targeting in the olfactory bulb during regeneration of olfactory neuroepithelium." Chem Senses 28: 773-779. Strotmann, J., Conzelmann, S., Beck, A., Feinstein, P., Breer, H. and Mombaerts, P. (2000). "Local permutations in the glomerular array of the mouse olfactory bulb." J Neurosci 15: 6927-6938. Strotmann, J., Hoppe, R., Conzelmann, S., Feinstein, P., Mombaerts, P. and Breer, H. (1999). "Small subfamily of olfactory receptor genes: structural features, expression pattern and genomic organization." Gene 236: 281-291. Strotmann, J., Wanner, I., Krieger, J., Raming, K. and Breer, H. (1992). "Expression of odorant receptors in spatially restricted subsets of chemosensory neurones." Neuroreport 3: 1053-1056. Sullivan, S. L., Adamson, M. C., Ressler, K. J., Kozak, C. A. and Buck, L. B. (1996). "The chromosomal distribution of mouse odorant receptor genes." Proc Natl Acad Sci USA 93: 884-888. Sullivan, S. L., Bohm, S., Ressler, K. J., Horowitz, L. F. and Buck, L. B. (1995). "Target-independet pattern specification in the olfactory epithelium." Neuron: 779-789. Suzuki, Y., Schafer, J. and Farbman, A. I. (1995). "Phagocytic Cells in the Rat Olfactory Epithelium after Bulbectomy." Experimental Neurology 136: 225233. Suzuki, Y. and Takeda, M. (1991). "Basal cells in the mouse olfactory epithelium after axotomy: immunohistochemical and electron-microscopic studies." Cell Tissue Res 266: 239-45. 67 Tanabe, T., Iino, M. and Takagi, S. F. (1975). "Discrimination of odors in olfactory bulb, pyriform-amygdaloid areas, and orbitofrontal cortex of the monkey." J Neurophysiol 38: 1284-1296. Thommesen, G. and Doving, K. B. (1977). "Spatial distribution of the EOG in the rat; a variation with odour quality." Acta Physiol Scand 99: 270-280. Trudeau, M. C. and Zagotta, W. N. (2003). "Calcium/Calmodulin Modulation of Olfactory and Rod Cyclic Nucleotide-gated Ion Channels." J Biol Chem 278: 18705-18708. Tsuboi, A., Miyazaki, T., Imai, T. and Sakano, H. (2006). "Olfactory sensory neurons expressing class I odorant receptors converge their axons on an antero-dorsal domain of the olfactory bulb in the mouse." Eur J Neurosci 23: 1436-44. Uchida, N., Takahashi, Y. K., Tanifuji, M. and Mori, K. (2000). "Odor maps in the mammalian olfactory bulb: domain organization and odorant structural features." Nature Neurosci 3: 1035-1043. Wachowiak, M. and Cohen, L. B. (2001). "Representation of odorants by receptor neuron input to the mouse olfactory bulb." Neuron 32: 723-735. Walz, A., Rodriguez, I. and Mombaerts, P. (2002). "Aberrant sensory innervation of the olfactory bulb in neuropilin-2 mutant mice." J Neurosci 22: 4025-4035. Wang, H.-W., Wysocki, C. J. and Gold, G. H. (1993). "Induction of olfactory receptor sensitivity in mice." Science 260: 998-1000. Wang, Y. and Sassoon, D. (1995). "Ectoderm-mesenchyme and mesenchymemesenchyme interactions regulate Msx-1 expression and cellular differentiation in the murine limb bud." Dev Biol 168: 374-82. 68 Vassar, R., Chao, S. K., Sitcheran, R., Nunez, J. M., Vosshall, L. B. and Axel, R. (1994). "Topographic organization of sensory projections to the olfactory bulb." Cell 79: 981-992. Vassar, R., Ngai, J. and Axel, R. (1993). "Spatial segregation of odorant receptor expression in the mammalian olfactory epithelium." Cell 74: 309318. Watanabe, K., Kondo, K., Takeuchi, N., Nibu, K. and Kaga, K. (2006). "Agerelated changes in cell density and the proliferation rate of olfactory ensheathing cells in the lamina propria of postnatal mouse olfactory mucosa." Brain Res doi:10.1016/j.brainres.2006.07.124 Wei, J., Wayman, G. and Storm, D. R. (1996). "Phosphorylation and Inhibition of Type III Adenylyl Cyclase by Calmodulin-dependent Protein Kinase II in Vivo." J Biol Chem 271: 24231-24235. Wei, J., Zhao, A. Z., Chan, G. C. K., Baker, L. P., Impey, S., Beavo, J. A. and Storm, D. R. (1998). "Phosphorylation and inhibition of olfactory adenylyl cyclase by CaM kinase II in neurons: a mechanism for attenuation of olfactory signals." Neuron 21: 495-504. Whitesides, J., Hall, M., Anchan, R. and LaMantia, A.-S. (1998). "Retinoid signaling distinguishes a subpopulation of olfactory receptor neurons in the developing and adult mouse." J. Comp. Neurol. 394: 445-461. Whitesides, J. G. r. and LaMantia, A. S. (1996). "Differential adhesion and the initial assembly of the mammalian olfactory nerve." J Comp Neurol 373: 24054. Wong, S. T., Trinh, K., Hacker, B., Chan, G. C. K., Xia, Z., Gold, G. H. and Storm, D. R. (2000). "Disruption of the type III adenylyl cyclase gene leads to peripheral and behavioral anosmia in transgenic mice." Neuron 27: 487-497. Vucinic, D., Cohen, L. B. and Kosmidis, E. K. (2006). "Interglomerular Center-Surround Inhibition Shapes Odorant-Evoked Input to the Mouse Olfactory Bulb In Vivo." J Neurophysiol 95: 1881-1887. 69 Wysocki, C. J., Whitney, G. and Tucker, D. (1977). "Specific anosmia in the laboratory mouse." Behav Genet 7: 171-188. Yan, C., Zhao, A. Z., Bentley, J. K., Loughney, K., Ferguson, K. and Beavo, J. A. (1995). "Molecular cloning and characterization of a calmodulindependent phosphodiesterase enriched in olfactory sensory neurons." Proc Natl Acad Sci 92: 9677-9681. Yoshihara, Y., Kawasaki, M., Tamada, A., Fujita, H., Hayashi, H., Kagamiyama, H. and Mori, K. (1997). "OCAM: a new member of the neural cell adhesion molecule family related to zone-to-zone projection of olfactory and vomeronasal axons." J Neurosci 17: 5830-5842. Yu, T. T., McIntyre, J. C., Bose, S. C., Hardin, D., Owen, M. C. and McClintock, T. S. (2005). "Differentially expressed transcripts from phenotypically identified olfactory sensory neurons." J Comp Neurol 483: 251-262. Zhang, X. and Firestein, S. (2002). "The olfactory receptor gene superfamily of the mouse." Nature Neurosci 5: 124-133. Zhang, X., Rogers, M., Tian, H., Zhang, X., Zou, D.-J., Liu, J., Ma, M., Shepherd, G. M. and Firestein, S. J. (2004). "High-throughput microarray detection of olfactory receptor gene expression in the mouse." Proc Natl Acad Sci U S A 101: 14168-14173. Zhao, H., Ivic, L., Otaki, J. M., Hashimoto, M., Mikoshiba, K. and Firestein, S. (1998). "Functional expression of a mammalian odorant receptor." Science 279: 237-242. Zhao, H. and Reed, R. R. (2001). "X inactivation of the OCNC1 channel gene reveals a role for activity-dependent competition in the olfactory system." Cell 104: 651-660. 70 Zhao, K., Dalton, P., Yang, G. and Scherer, P. (2006). "Numerical modeling of turbulent and laminar airflow and odorant transport during sniffing in the human and rat nose." Chem Senses. 31: 107-18. Zheng, J. and Zagotta, W. N. (2004). "Stoichiometry and Assembly of Olfactory Cyclic Nucleotide-Gated Channels." Neuron 42: 411-421. Zou, D.-J., Feinstein, P., Rivers, A. L., Mathews, G. A., Kim, A., Greer, C. A., Mombaerts, P. and Firestein, S. (2004). "Postnatal refinement of peripheral olfactory projections." Science 304: 1976-1979. Zou, Z. and Buck, L. (2006). "Combinatorial effects of odorant mixes in olfactory cortex." Science 311: 1477-81. Zou, Z., Horowitz, L. F., Montmayeur, J.-P., Snapper, S. and Buck, L. B. (2001). "Genetic tracing reveals a stereotyped sensory map in the olfactory cortex." Nature 414: 173-179. Zou, Z., Li, F. and Buck, L. B. (2005). "Odor maps in the olfactory cortex." Proc Natl Acad Sci U S A 102: 7724-7729. Zufall, F. and Leinders-Zufall, T. (1997). "Identification of a Long-Lasting Form of Odor Adaptation that Depends on the Carbon Monoxide/cGMP SecondMessenger System." J Neurosci 17: 2703-2712. Zufall, F. and Leinders-Zufall, T. (2000). "The cellular and molecular basis of odor adaptation." Chem Senses 25: 473-481. 71