Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Non-coding DNA wikipedia , lookup
Community fingerprinting wikipedia , lookup
RNA polymerase II holoenzyme wikipedia , lookup
Genome evolution wikipedia , lookup
Epitranscriptome wikipedia , lookup
Non-coding RNA wikipedia , lookup
Eukaryotic transcription wikipedia , lookup
Gene expression profiling wikipedia , lookup
List of types of proteins wikipedia , lookup
Molecular evolution wikipedia , lookup
Gene expression wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Gene regulatory network wikipedia , lookup
Biochemistry wikipedia , lookup
Promoter (genetics) wikipedia , lookup
Transcriptional regulation wikipedia , lookup
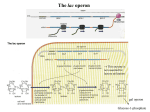
The lactose (lac) operon – an example for prokaryotic gene regulation SLIDE 1 Nobel Prize Two French researchers, Francois Jacob and Jacques Monod, working at the Pasteur Institute in Paris, for their work in elucidating the regulatory mechanism of lac operon, received the Nobel Prize in Physiology or Medicine in 1965. They examined two mutations: induced mutations of coding and regulator regions. SLIDE 2 Structure of the lac operon-I The lac operon is the best-known genetic regulation unit of a human enteric bacterium: Escherichia (E.) coli. With the presentation of the function of lac operon, we will present the logic of a prokaryotic gene regulation. The lac operon consists of: (1) a promoter, (2) an operator and (3) three adjacent structural genes. The promoter consist of 2 domain: (a) RNApolymerase-binding domain and (b) CAP (catabolite activator protein)-binding domain. The three structural genes are: lacZ gene encoding β-galactosidase, lacY encoding permease enzyme, and lacA encoding transacetylase. The function of β-galactosidase is the cleavage of the disaccharide lactose into glucose and galactose molecules. Permease, a membrane-bound transport protein pumps lactose into the cell. The accurate function of transacetylase is not known. There is a terminator sequence at the end of the polycistronic DNA (consist of more than one gene under the control of a single promoter). Terminator marks the end of operon of genomic DNA for transcription. The lacI gene encoding the repressor is not part of lac operon (in fact, it can be considered as a specific operon containing a single gene). SLIDE 3 Structure of the lac operon-II There are two initiation sites at a DNA sequence: (1) the transcription start site, which is within the operator region; and (2) the translation start site (it is also called as Shine-Dalgarno sequence in bacteria). The repressor binds to the initiation site: it physically blocks the RNA-polymerase and thus, the transcription of the mRNA is not possible. SLIDE 4 Function of the lac operon - repression The repressor is produced at a very low level, but it bounds to the operator with high efficiency when lactose (milk sugar) is absent in the cytoplasm. So, if the substrate (lactose) is missing from the growth medium, the function of the operon is blocked by the repressor, and through its blocking, the RNA polymerase cannot start the transcription. The logic of the system is based on the economy: few repressors block the expression of three genes, which work intensively. SLIDE 5 Function of the lac operon - induction When cells are grown in the presence of lactose: lactose binds to the repressor which is unable to bind to the operator, allowing RNA-polymerase to transcribe the polycistronic mRNA and thereby leading to high levels of the encoded proteins. βgalactosidase cleaves the lactose into glucose and galactose. Effectively, the sense of the operation of lac operon, is the production of glucose, if it is not present in the environment. Glucose is the most important source of carbon and energy of the living world. SLIDE 6 Operation of the lac operon - activation – Cell can take up glucose from the surroundings (the simpler solution) or can get it by cleavage of lactose. When glucose is absent, the amount of cAMP (cyclic adenosine monophosphate) is high (see later for the reason). cAMP binds the so called catabolite activator protein (CAP; not to be confused with the cap on the 5’-ends of mRNAs!), and in turn, this complex binds to the CAP site of the promoter. Binding of CAP is a prerequisite of binding of RNA polymerase. In the absence of glucose, the prevalence of cAMP, the binding of the CAP to the DNA significantly increases the production of β-galactosidase, enabling the cell to digest the lactose needed to produce glucose. Conversely, in the presence of glucose the lac operon remains silent since no cAMP-CAP complex is formed. SLIDE 7 Summary: operation of lac operon is regulated by two factors: absence or presence of lactose and glucose. The logic of the system: 1 Absence of glucose: enzymatic cleavage of lactose into glucose (and galactose) Presence of glucose: stop the operation of the lac operon Catabolite repression: a process when the end product of the reaction stops the reaction. cAMP is synthesized from ATP by adenylate cyclase enzyme. Under conditions of high glucose content, a glucose breakdown product inhibits the enzyme adenylate cyclase, preventing the conversion of ATP into cAMP: more glucose less cAMP (and vice versa). The lactose is a stimulating and the glucose is an inhibitor of the lac operon. SLIDE 8 Mutant -gal Created mutant lacZ gene RNA-polymerase proceeds to transcribe all three genes (lacZ, lacY and lacA) into mRNA. -galactosidase molecules will not function lac operon functions in the presence of lactose and absence of glucose, but lactose is not cleaved Catabolite repression will not occur SLIDE 9 Transcriptional stop in the genes Jacob and Monod’s observations: there is a mutation, which stops the translation, in the structural genes: Mutation in lacA gene: lacZ and lacY transcribed normally, lacA is not transcribed Mutation in lacY gene: lacZ transcribed, but lacA and lacY are not transcribed!!! Mutation in lacZ gene = transcriptional stop: lacA, lacY and lac Z are not transcribed Conclusion: the order of the genes: lacZ, lacY, lacA. SLIDE 10 The Oc mutation of the operon Lac operon was active, all the three structure genes were transcribed and translated in the absence of lactose (and absence of glucose!). Because of this mutation occurred in the operator region, repressor was not able to bind to the structure region. SLIDE 11 The I- mutation of the repressor Due to this mutation, repressor did not work: lac operon was active in the absence of glucose and lactose. SLIDE 12 The IS mutation of the repressor Repressor could not bind lactose because of this type of the mutation of the repressor. There was lactose in the cytoplasm (there wasn’t glucose), but it did not induce to operate the operon, as a result of this mutation. SLIDE 13 Mutant CAP-I CAP was not able to bind cAMP lac operon did not operate (nevertheless, absence of glucose, presence of cAMP). The reason: RNA polymerase didn’t recognize the promoter, because of the absence of CAP SLIDE 14 Mutant CAP-II CAP bound to the promoter without binding of the cAMP due to another type of CAP mutations. In this case, the lac operon operated in the presence of glucose. SLIDE 15 Too many repressors We take many of the copies of the repressor gene into the cell (E. coli) repressors inhibit the transcription of the lac operon in the presence of lactose. The reason: the system works dynamically: the repressor-DNA and the repressor-lactose binding are continuously disintegrating and establishing contacts. If there are enough repressors, the low amount of lactose cannot remove efficiently the repressor molecules from the operator region. SLIDE 16 Too many operators In the absence of lactose, the operation of the lac operon is repressed. We put many copies of the operator sequence into the cell, therefore the repressor will be unable to inhibit the lac operon. SLIDE 17 Partial diploids - I The technological opportunities were not as good in Jacob and Monod 's time as today. It was necessary to clear it up, in a first step, that the regulator elements are cis-acting elements (they are found on the DNA strand), or trans-factors (they float freely in the cytoplasm). The function of the operator was established as follows: Jacob and Monod used a lacY and Oc mutant of E. coli, which didn’t block the transcription of lacA and lacZ genes. This type of mutant bacteria produced normal lacZ and lacA products, in the presence of lactose (and in the absence of glucose). If we take a lacZ mutant lac operon with F-plasmid into this E. coli (operator is normal) in the absence of lactose, product of lacY and lacA will be produced, but the product of lacZ gene will not. If we add lactose to 2 induce the system, -galactosidase (lacZ gene product) will appear. Conclusion: operator is a cis-factor (for explanation listen to the lecturers). SLIDE 18 Partial diploids - II In another experiment Jacob and Monod used a lacY and lacI- mutant bacteria strain. The same lacZ mutant F-plasmid was used, as in the previous experiment, plasmid contained only a single intact repressor. Operon of plasmid DNA and bacteria were not active in absence of lactose. All the three proteins appeared due to the lactose induction. Conclusion: repressor floats freely in the cytoplasm, and therefore it is a trans-acting factor. MINIMAL REQUIREMENTS: SLIDES 1 - 14 EXTRA REQIREMENTS: SLIDES 15 - 18 3