Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Development of the nervous system wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Synaptic gating wikipedia , lookup
Caridoid escape reaction wikipedia , lookup
Embodied language processing wikipedia , lookup
End-plate potential wikipedia , lookup
Proprioception wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Electromyography wikipedia , lookup
Central pattern generator wikipedia , lookup
Synaptogenesis wikipedia , lookup
Microneurography wikipedia , lookup
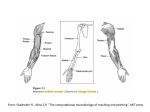
Organization of motor system Muscles Spinal reflexes Squire Ch 29 Rosenbaum Ch 3 pp 46-61 Pathways for signals to muscles Red nucleus Hierarchical Organization of the motor system Somatosensory ctx Planning/ sequences Supplementary motor ctx Selection of trajectory Activity prior to movement Target selection Posterior parietal ctx Pre-motor cortex M1 Primary motor ctx eyes ?? Muscle commands Cortico-spinal tract Initiation of movement “Efferent copy” Visual consequences of movement Smoothness, timing Learning new skills Monitor feedback 80 msec Spinal feedback: 30-50 msec Proprioceptive signals (muscles, joints) Somatosensory ctx Pre-motor ctx Reach (unfolded) Primary motor ctx FEF Intra-parietal sulcus Grasp V5(MT/MST) V1 A “motor unit” is defined as one motor neuron And all the muscle fibers it innervates. In mammals, each muscle fiber is innervated by only one motor neuron, but one motor neuron innervates many muscle fibers as its axon branches in the muscle. Hand: 100 muscle fibres per motor unit Leg: 1000 muscle fibres per motor unit Figure 34-1 A typical muscle consists of many thousands of muscle fibers working in parallel and organized into a smaller number of motor units. A motor unit consists of a motor neuron and the muscle fibers that it innervates. The motor neurons innervating one muscle are usually clustered into an elongated motor nucleus within the ventral spinal cord that may extend over 1-4 segments. In the example shown here the motor neuron A1 plus other motor neurons innervating muscle A form motor nucleus A. Muscle B is innervated by motor neurons lying in a separate motor nucleus B. Motor Unit Intrafusal fibre connected to a single extrafusal fibre Contracts Intrafusal Contracts Extrafusal Alpha and gamma motor neurons co-activate 1a afferents respond to a difference in length between The intra and extrafusal fibres Golgi (Ib) thresholds regulated by central signals. Activity of Ia afferents corresponds to awareness of stretch Golgi tendon organ is located at the transition between muscle and tendon. It senses any active tension produced by the muscle fibers, being in series between muscle and tendon. The muscle spindle is located in parallel with the extrafusal muscle fibers. It signals the muscle length and dynamic changes in muscle length. Both muscle spindle and Golgi tendon organ have fast conducting afferent nerve fibers in the range of ∼100 m/s. Mechanism of muscle contraction sarcomere myosin actin myasthenia gravis, caused by antibodies that bind to AChRs at the neuromuscular junction B. The sarcomere is the functional unit of the muscle. It contains contractile proteins, the thick and thin filaments, bounded by thin Z disks, from which the thin filaments arise. Thick and thin filaments overlap, creating alternating dark bands that give skeletal muscle its characteristic striated appearance. This banded pattern changes as the overlap between the thin and thick filaments changes during shortening or lengthening of the muscle fiber. Contraction is produced by cyclical attachment and detachment of myosin heads on adjacent thin filaments. Figure 34-5 The amount of active contractile force developed during contraction depends on the degree of overlap of thick and thin filaments. When the sarcomere is stretched beyond the length at which the thick and thin filaments overlap (length a), no active force develops because the myosin heads are not near any binding sites and thus cross bridges cannot form. As the filaments overlap (lengths a-b) the force that can develop increases linearly as length decreases because of the progressive increase in the number of binding sites for myosin heads. Around the muscle's optimal length (L0, between lengths b-c) the level of force remains constant because the central portion of the thick filaments is devoid of myosin heads. With further reductions in length (lengths c-d) the progressive overlap of thin filaments with each other occludes potential attachment sites and the force begins to fall. Once the thick filaments abut the Z disks (lengths d-e), they act like compression springs opposing the active force generated by the cross bridges. Passive force exists in muscle regardless of activation, starting at about L0 and rising at first exponentially and then linearly as progressive lengthening of the muscle stretches the connectin filaments that tether the thick filaments between the Z disks. Total force is the sum of active and passive force. Muscle can exert most force when joints are in the mid-range. At extreme joint angles the muscle is less capable of exerting force. Different kinds of muscle fiber Red muscles - type I fibers type IIA fibers type IIB fibers Slow rise fatigue resistant Smaller, exert less force Exert more force- use different form of myosin When a motor nucleus begins to be activated by peripheral or descending inputs, individual motor neurons begin firing at a slow regular rate (5-10 impulses per second in humans). This results in a partially fused train of contractions in the target muscle fibers. As the net excitatory synaptic input in the nucleus increases, the firing rate of the cells increases and other, slightly larger motor neurons reach their threshold for firing, adding their force as well. In this way the mean level of force produced in the muscle gradually increases (Figure 34-12). The overall force of a contraction depends on both the number and size of active motor units and their individual firing rates. Spinal Cord Feedback from muscle receptors to motoneurons: the stretch reflex. The muscle spindle activates a-motoneurons directly, which causes the muscle fibers to contract. The sensitivity of the muscle spindle can be actively regulated by g-motoneurons, which are more slowly conducting. The muscle spindle provides negative feedback. If the muscle with its muscle spindle is lengthened, the afferent activity from the muscle spindle increases, exciting the a-motoneurons and leading to an increased muscle contraction, which in turn counteracts the lengthening. This is called the stretch reflex. The Golgi tendon organ provides force feedback. The more the muscle contracts, the more the Golgi tendon organ and its afferent are activated. In the diagram the interneuron between afferent nerve fiber and motoneuron is inhibitory. Thus increased muscle force leads to inhibition of the a-motoneuron, which results in a decrease of muscle force. The efficacy of length and force feedback can be regulated independently in the spinal cord and via g-motoneurons. Thus their respective contributions can vary considerably between different patterns of motor behavior. Role of gamma motor neurons is to prevent the spindle sensory fiber from falling silent when the muscle shortens as a result of active contraction, enabling it to signal length changes over the full range of muscle lengths. Reciprocal inhibition in the spinal cord Modulation of spinal reflexes by descending innervation from M1 and brainstem. A. The Ia inhibitory interneuron allows higher centers to coordinate opposing muscles at a joint through a single command. This inhibitory interneuron mediates reciprocal innervation in stretch reflex circuits. In addition, it receives inputs from corticospinal descending axons, so that a descending signal that activates one set of muscles automatically leads to relaxation of the antagonists. Other descending pathways make excitatory and inhibitory connections to this interneuron. When the balance of input is shifted to greater inhibition of the Ia inhibitory interneuron, reciprocal inhibition will be decreased and co-contraction of opposing muscles will occur. B. Renshaw cells produce recurrent inhibition of motor neurons. These spinal interneurons are excited by collaterals from motor neurons and then inhibit those same motor neurons. This negative feedback system regulates motor neuron excitability and stabilizes firing rates. Renshaw cells also send collaterals to synergist motor neurons (not shown) and Ia inhibitory interneurons. Thus, descending inputs that modulate the excitability of the Renshaw cell adjust the excitability of all the motor neurons around a joint. Renshaw cells modulate alpha motor neurons Short-latency and Long-latency responses: M1 = short, M2 = long Central Pattern Generations: neural networks for control of rhythmic movements Central Pattern Generators A. Transection of the spinal cord of a cat at the level of b-b¢ isolates the hind limb segments of the cord, but but the hind limbs are still able to step on a treadmill B. Depending on the exact level of the transection of the brain stem, locomotion either occurs spontaneously (cut 1) or can be initiated by electrical stimulation of the mesencephalic locomotor region (MLR) after a more caudal transection (cut 2). Thal = thalamus; SC = superior colliculus; MB = mammillary body. C. The spinal cord is removed from a neonatal animal (0-5 days of age) and placed in a saline bath. Addition of N-methyl- D-aspartate (NMDA) and serotonin (5-hydroxytryptamine, or 5-HT) to the bath elicits rhythmic bursting in the motor neurons supplying leg muscles. Complex behavior in the spinal frog. Spinal cord must be able to relate the sensory and motor maps.