Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Point mutation wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Human genome wikipedia , lookup
Non-coding DNA wikipedia , lookup
Oncogenomics wikipedia , lookup
Extrachromosomal DNA wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
Epigenetics of human development wikipedia , lookup
X-inactivation wikipedia , lookup
No-SCAR (Scarless Cas9 Assisted Recombineering) Genome Editing wikipedia , lookup
Genomic imprinting wikipedia , lookup
Public health genomics wikipedia , lookup
Genomic library wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Mir-92 microRNA precursor family wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Minimal genome wikipedia , lookup
Genome evolution wikipedia , lookup
Genome editing wikipedia , lookup
Designer baby wikipedia , lookup
Microevolution wikipedia , lookup
Genetic engineering wikipedia , lookup
Genome (book) wikipedia , lookup
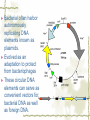
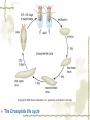
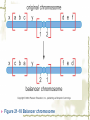
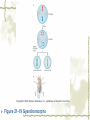
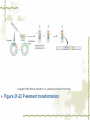
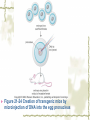
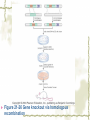
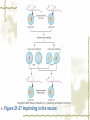
Model Organisms Model Organism Important features of all model organisms Short lifespan Small, easy and inexpensive to maintain Produce large numbers of offspring Development external as well as internal Availability of mutants History/previous experiments and discoveries Genome is sequenced Homologues for large % of human disease genes Exhibit complex behaviors Few ethical concerns The choice of a model organism depends on what question is being asked. Specific species Uniform from research lab to research lab Ability to apply new knowledge to other organisms Advance our understanding of Cellular function Development Disease Model Organisms E. coli Drosophila Xenopus Zebrafish Mouse C. elegans Yeast Arabidopsis The Nematode Worm Caenorhabditis elegans In 1965, Sydney Brenner settled on the small nematode worm Caenorhabditis elegans to study the important questions of development and the molecular basis of behavior, because of their suitable characteristics. Due to its simplicity and experimental accessibility, it is now one of the most completely understood metazoans. What is unique to this organism is that wild-type individuals contain a constant 959 cells. The position of cells is constant as is the cell number. If the 6th chromosome pair is XX, then C. elegans will be a hermaphrodite. A XO combination in the 6th chromosome pair will produce a male. Hermaphrodites can self-fertilize or mate with males but cannot fertilize each other. In nature, hermaphrodites are the most common sex. C. elegans has a very rapid life cycle C.elegans is transparent. It is easy to track cells and follow cell lineages. The genome size of C. elegans is about a hundred million base pairs. This is approximately 20X bigger than that of E. coli and about 1/30 of that of human. At 25℃, fertilized embryos of C. elegans complete development in 12 hours and hatch into free-living animals capable of complex behaviors. The first stage juvenile(L1) passes through four juvenile stages(L1-L4) over the course of 40 hours to become a sexually mature adult. The life cycle of the worm, C.elegans Under stressful conditions, the L1 stage animal can enter an alternative developmental stage in which it forms what is called a dauer. Dauers are resistant to environmental stresses and can live many months while waiting for environmental conditions to improve. C. elegans’s cell lineages C. elegans has a simple body plan. Its cell lineages are relatively few and well studied. Among C.elegans genes are components of highly conserved receptor tyrosine kinase signaling pathways that control cell proliferation. Many of the mammalian homologs of these genes are oncogenes and tumorsupressor genes that when altered can lead to cancer. The cell death pathway was discovered in C. elegans The most notable achievement to date in C. elegans research has been the elucidation of the molecular pathway that regulates apoptosis or cell death. Analysis of the ced mutants showed that, in all but one case, developmentally programmed cell death is cell autonomous, that is, the cell commits suicide. Cell death is as important as cell proliferation in development and disease and is the focus of intense research to develop therapeutics for the control of cancer and neurodegenerative diseases. RNAi was discovered in C. elegans In 1998 a remarkable discovery was announced. The introduction of dsRNA into C. elegans silenced the gene homologous to the dsRNA. It is significant in two respects. One is that RNAi appears to be universal since introduction of dsRNA into nearly all animal, fungal, or plant cells leads to homology-directed mRNA degradation. The second was the rapidity with which experimental investigation of this mysterious process revealed the molecular mechanisms. Bacteria The attraction of bacteria such as E. coli or B. subtilis as experimental systems is that they are relatively simple cells and can be grown and manipulated with comparative ease. Molecular biology owes its origin to experiments with bacterial model systems. Assays of bacterial growth Bacterial cells are large enough, about 2µm, in length to scatter light, allowing the growth of a bacterial culture to be measured conveniently in liquid culture by the change in optical density. The number of bacteria can be determined by diluting the culture and plating the cells on solid (agar) medium in a petri dish. Knowing how many colonies are on the plate and how much the culture was diluted makes it possible to calculate the concentration of cells in the original culture. Bacterial often harbor autonomously replicating DNA elements known as plasmids. Evolved as an adaptation to protect from bacteriophages These circular DNA elements can serve as convenient vectors for bacterial DNA as well as foreign DNA. BAKER’S YEAST Saccharomyces cerevisiae Unicellular eukaryotes offer many advantages as experimental model systems. The best studied unicellular eukaryote is the budding yeast S. cerevisiae. These cell types can be manipulated to perform a variety of genetic assays. The genetic analysis of S. cerevisiae is further enhanced by the availability of techniques used to precisely and rapidly modify individual genes. Generating precise mutations in yeast is easy recombinational transformation in yeast The ability to make such precise changes in the genome allows very detailed questions concerning the function of particular genes or their regulatory sequences to be pursued with relative ease. Because of its rich history of genetic studies and its relatively small genome, S. cerevisiae was chosen as the first eukaryotic organism to have its genome entirely sequenced. This landmark was accomplished in 1996. S. cerevisiae cells change shape as they grow Simple microscopic observation of S. cerevisiae cell shape can provide information about the events occurring inside the cell. A cell that lacks a bud has yet to start replicating its genome. A cell with a very large bud is almost always in the process of executing chromosome segregation. The Fruit Fly Drosophila melanogaster Drosophila has a rapid life cycle The salient features of the Drosophila life cycle are a very rapid period of embryogenesis, followed by periods of larval growth prior to metamorphosis. The Drosophila life cycle One of the key processes that occurs during larval development is the growth of the imaginal disks, which arise from invaginations of the epidermis in mid-stage embryos. Imaginal disks differentiate into their appropriate adult structures during metamorphosis (or putation). Figure 21-16 Imaginal disks in Drosophila The first genome maps were produced for Drosophila Morgan’s lab studies on Drosophila (1910) led to two major discoveries: genes are located on chromosomes, and each gene is composed of two alleles that assort independently during meiosis; genes located on separate chromosomes segregate independently, whereas those linked on the same chromosome do not. Hermann J. Muller provided the first evidence that environmental factors can cause chromosome rearrangements and genetic mutations. Bridges used the polytene chromosomes to determine a physical map of the Drosophila genome (the first produced for any organism). Figure 21-17 Genetic maps, polytene chromosome, and deficiency mapping A variety of additional genetic methods were create to establish the fruit fly as the premiere model organism for studies in animal inheritance. For example, balancer chromosomes were created that contain a series of inversions relative to the organization of the native chromosome. Figure 21-18 Balancer chromosome Embryos that contain two copies of the balancer chromosome die because some of the inversions produce recessive disruptions in critical genes. In addition, embryos that contain two copies of the normal chromosome die because they are homozygous for the eve null mutation. Genetic mosaics permit the analysis of lethal genes in adult flies Mosaics are animals that contain small patches of mutant tissue in a generally “normal” genetic background. The analysis of genetic mosaics provided the first evidence that Engrailed is required for subdividing the appendages and segments of flies into anterior and posterior compartments. The most spectacular genetic mosaics are gynandromorphs. Figure 21-19 Gyandromorphs These are flies that are literally half male female. The X instability occurs only at the first division. And the “line” separating the male and female tissues is random. Its exact position depends on the orientation of the two daughter nuclei after the first cleavage. The yeast FLP recombinase permits the efficient production of genetic mosaics Drosophila possesses several favorable attributes for molecular studies and wholegenome analysis. Most notably, the genome is relatively small. The frequency of mitotic recombination was greatly enhanced by the use of the FLP recombinase from yeast. Figure 21-20 FLP-FRT This method is quite efficient. In fact, short pulse of heat shock are often sufficient to produce enough FLP recombinase to produce large patches of zˉ/zˉ tissue in different regions of an adult fly. It is easy to create transgenic fruit flies that carry foreign DNA P-elements are transposable DNA segments that are the causal agent of a genetic phenomenon called hybrid dysgenesis. Figure 21-21 Hybrid dysgenesis P-element excision and insertion is limited to the pole cells, the progenitors of the gametes (sperm in males and eggs in females). P-elements are used as transformation vectors to introduce recombinant DNAs into otherwise normal strains of flies. Figure 21-22 P-element transformation This method of P-element transformation is routinely uses to identify regulatory sequences such as those governing eve stripe 2 expression. In addition, this strategy is used to examine protein coding genes in various genetic backgrounds. The House Mouse, Mus musculus The mouse enjoys a special status due to its exalted position on the evolutionary tree: it is a mammal and, therefore, related to humans. The mouse provides the link between the basic principles, discovered in simpler creatures like worms and flies, and human disease. Mouse Embryonic Development Depends on Stem Cells Their small size prohibits grafting experiments of the sort done in zebrafish and frogs, but microinjection methods have been developed for introducing. Figure 21-23 shows an overview of mouse embryogenesis. Figure 21-23 Overview of mouse embryogenesis It Is Easy to Introduce Foreign DNA into the Mouse Embryo DNA is injected into the egg pronucleus, and the embryos are places into the oviduct of a female mouse and allowed to implant and develop. The injected DNA integrates at random positions in the genome Figure 21-24 Creation of transgenic mice by microinjection of DNA into the egg pronucleus Figure 21-25 In situ expression patterns of embryos obtained from transgenic mice Homologous Recombination Permits the Selective Ablation of Individual The single most Genes powerful method of mouse transgenesis is the ability to disrupt, or “knock out,” single genetic loci. This permits the creation of mouse models for human disease. Gene disruption experiments are done with embryonic stem (ES) cells Figure 21-26 Gene knockout via homologous recombination Mice Exhibit Epigenetic Inheritance Studies on manipulated mouse embryos led to the discovery of a very peculiar mechanism of non-Mendelian, or epigenetic, inheritance. This phenomenon is known as parental imprinting. Figure 21-27 Imprinting in the mouse The basic idea is that only one of the two alleles for certain genes is active. It has been suggested that imprinting has evolved to protect the mother from her own fetus.