Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Optogenetics wikipedia , lookup
Development of the nervous system wikipedia , lookup
Synaptogenesis wikipedia , lookup
Subventricular zone wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Signal transduction wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
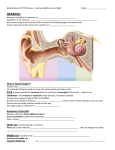
The Senses We have 5 senses: touch (including pressure) smell taste hearing vision Each sense has specialized receptor cells associated with it. Receptor cells are depolarized (channels open to allow sodium to rush in) by the stimulus. The depolarization (a generator potential) leads to synaptic activity where the receptor cell connects to a sensory neuron. Like the post-synaptic potentials that sum to determine whether a nerve impulse will arise, the generator potentials from receptors sum to determine whether (and at what frequency) nerve impulses will arise in sensory neurons. Generator potentials thus determine one aspect of neural coding – frequency of impulses (frequency coding) as an indication of intensity of stimulation. The other aspect of coding is called line labeling. Your brain knows what kind of stimulus it is by which neurons are delivering the information. Errors can arise in line labeling: close your eyes; press gently a finger gently against your eyelid. When you press and when you release the pressure most of us will ‘see’ what we interpret as colour. The pressure caused some generator potentials in receptor cells of the retina. We ‘misinterpret’ the stimulus as if it were coloured light. Now let’s consider each of the senses individually: Touch (and other skin senses) What is classed within this ‘sense’ are: touch, pain, temperature (both hot and cold) and pressure. There are different sensory cells for these ‘senses’. Touch and pain are both sensed by ‘free’ nerve endings in the skin and some touch by enclosed endings (Meissner’s corpuscles). What differs is their sensitivity. Touch sensors can be exquisitely sensitive, e.g. on the face, lips and fingertips. Free nerve endings are frequently wrapped around the base of hair follicles. All touch sensors are distorted and become leaky by the stimulus. For free endings around hair follicles, amount of bending determines the size of the generator potential, and, thus, the frequency of nerve impulses. Pain sensors, looking essentially identical, are insensitive. Most of their response is the result of physical damage to the nerve ending. Temperature sensors are generally enclosed nerve endings (bulbs of Krause). The enclosure is simple. The last type is the pressure sensors. They are enclosed in an onion-like leyered structure called a Pacinian corpuscle). The layering has the effect of ‘averaging out’ the pressure over a small region. Chemoreceptors (taste and smell) Taste and smell are closely related. Most tastes and many smells are compound responses involving both senses. There is a greater diversity of types (what they respond to) among smell receptors than among taste ones. Four different taste receptors are generally distinguished: sweet – near the tip of the tongue salty – along the sides, but nearer the front sour – along the sides but toward the rear bitter – the rear surface of the tongue A taste bud (shown below) looks somewhat like an orange, with a number of cells clustered together like segments of the orange. At the top of the cells are microvilli at an opening, a pit in the tongue. Chemicals dissolved in saliva bind to receptor proteins in the cell membranes of microvilli and stimulate the receptor cells. Taste receptors are not neuronal, and are replaced by other bud cells (segments of the orange). There is continuing cell division in each bud, and segments ‘mature’ to become active sensory receptors. Smell Sensory cells for olfaction are located in the olfactory bulb in the uppermost part of the nasal cavity. Here, each receptor is a neuron, whose axon passes through a spongy bit of bone (the ethmoid bone) and to the olfactory region of the cerebrum. The receptor cells have long cilia that extend into the nasal cavity. The cilia act as the receptive surface. Molecules dissolved or suspended in mucus stimulate the receptors by molecular shape-dependent binding. In many animals it may take only one or very few molecules of an odorant to produce action potentials in receptor axons. How many odors are there? There are probably only a few ‘categories’ – maybe 7or less, but multiple receptors respond to single, complex odors, so that a trained perfume sniffer can distinguish >10,000 different odors. Here is one categorization: etherial – small molecules like simple anaesthetics and solvents camphorous – like tiger balm musky – large, complex molecules like the secretions from mustelid (e.g. skunk) anal glands floral – also large molecules, but based on carbon chains and ring structures minty – different ring structures Hearing (and balance) We hear sound over a range of frequencies from ~20 Hz to 20,000 Hz (when we’re young, then parts of the cochlea stiffen, and we lose some response to the high frequencies). Velocity of sound propagation (~1100km/hr, or Mach 1) is distinct from frequency, measured in Hertz (Hz). The sensory receptors for hearing are hair cells located in the cochlea of the inner ear. For sound to stimulate those hair cells, there is a relatively complex apparatus that transmits vibration to the cochlea… The pinna (external ‘ear’) acts like an old-fashioned ear trumpet, collecting sound waves from the environment and passing them down the auditory canal. At the inner end of the canal is the eardrum, a thin membrane. Against it on the ‘inside’ is the first of a set of 3 tiny bones (the hammer, the anvil, and the stirrup, in order) that transmit the vibration (and amplify the amplitude of the waves) to the oval window. The oval window is the entry point into the cochlea for vibration. You can think of the cochlea as a little like a French horn played backwards. The oval window brings vibration to the cochlea at the cider end of the coil. However, the cochlea is not a tube with only a single channel… The cochlea actually has 3 channels, the vestibular canal (or scala vestibuli), the cochlear duct (or scala media) and the tympanic canal (scala timpani). The tympanic and vestibular canals are open to each other at the far (small) end of the cochlea. The oval window is at the large end of the vestibular canal. Vibrations pass down the vestibular canal and back up the tympanic canal. The vibration energy is dissipated at the round window at the head of the tympanic canal. So how does this vibration stimulate hearing? Lying on the bottom of the cochlear canal, which is also the top of the tympanic canal, is the receptor organ, the organ of Corti. The receptor cells are the hair cells. The hairs projecting from their upper surface lie against the tectorial membrane. It’s gelatinous and pretty much stationary. When sound waves pass through the tympanic canal, the organ of Corti vibrates up and down. The hairs of the receptors are bent, and produce generator potentials when they bend. O.K., so we can sense sound. How do we distinguish frequencies? The cochlea changes in diameter along its length. Only a specific region of the membrane at the top of the tympanic canal resonates to any given frequency. You can begin to see how if we look at the cochlea as if it were unrolled… High frequencies excite the part of the organ of Corti nearest the oval window, and low frequencies excite the region near the apex. Sound intensity is determined by the amplitude of the sound wave, the amplitude of movement of the floor of the tympanic canal, and finally the amount the hairs are bent. Sound can get too loud. We have a protective response; muscles attached to the middle ear bones dampen amplitude. However, the hairs can take only so much. Repeated exposure to very loud sound eventually begins to break off hairs; hearing loss occurs. Where, in an electron micrograph of a healthy ear, the hair cells look like a forest, some rock musicians (and others similarly exposed) look like the forest has been clearcut. Balance and orientation Above the oval window are the semicircular canals (3 of them) and the saccule and utricle. The saccule and utricle are open saces lined with hair cells, and with calcium carbonate (limestone) granules inside. See, you do have rocks in your head! The granules fall to the bottom of the sacs due to gravity, bending hairs of cells beneath. Orientation (right side up, upside down, lying on your side,…) is indicated by which cells are producing generator potentials. The 3 semicircular canals are oriented in the three possible planes. They are fluid filled, with hair cells lining them. The hairs move in the fluid ‘breeze’. Stand still, and there is no ‘breeze’. Start moving, and the fluid in the canals has inertia, it ‘falls behind’ and hairs are bent, causing generator potentials. Stop and the same inertia keeps the fluid moving – more generator potentials. Note that the big signals are sent when you begin or stop moving. The semicircular canals are sending signals indicating acceleration. If you spin for a while, the fluid catches up and signals stop. Then you stop spinning. Your eyes tell you you’ve stopped, but the semicircular canals are sending a conflicting signal. That’s believed to be what causes dizziness and motion sickness. Vision Most animals are sensitive to light. The simplest have photoreceptors that can cause an animal to move (a kinesis) until it reaches a dark area. This sort of response persists into flatworms and others that have eyecups. Higher invertebrates and vertebrates have camera-like eyes with a single lens. The only real difference between our eyes and those of a squid is the orientation of the light receptors in the retina: in the squid they are pointed toward the light source; in ours they are facing backwards. That makes the squid eye more sensitive in its low-light environment. The analogies to a camera: The sclera (the white of your eye) is the body of the camera The choroid is a black layer like the black paint inside the camera; it prevents reflections The lens focuses images onto your retina. It can get thicker or thinner due to contraction or relaxation of the ciliary body (muscle) The cornea is the clear covering in the front. Since it is a curved surface, it really does most of the focusing (at least for objects > 6m away), but is not adjustable. The iris controls how much light enters the eye Our ability to accommodate, to change the shape of the lens by making it thicker, rounder to focus on nearer objects, and thinner to focus on more distant objects, decreases with age. The lens loses the elasticity that allows it to thicken and round to focus on near objects. The problem is called presbyopia, which means “old eye” in Greek. A stiffer lens doesn’t thicken this much in old age. The final analogy is the retina. It functions as the ‘film’ in your camera-eye. At the front are ganglion cells; they are neurons whose axons form the optic nerve. They integrate information from a number of bipolar cells. In the middle are bipolar cells. They collect input from one or more receptor cells, and pass it on to one or more ganglion cells. At the back are the receptor cells. There are two types: Cones provide colour information in the central focus area of the retina, called the fovea. These cells are packed very tightly together, and contain one of three pigments, called photopsins, that respond to different colours of light. Rods are present in the remainder of the retina. They contain the pigment rhodopsin; provide only black-andwhite information. They are, however, far more sensitive to low light. The discs on the outer segments of the receptors contain the pigments. Light causes the pigments to change conformation, opening channels in the membrane and causing a generator potential. How does this become a visual perception? The ganglion cells are the first level of integration. They are excited by impulses from some bipolar cells, but inhibited by others. What results is what is called a “receptive field”, typically “on-center” and “offsurround”. On if this area is lit Off (inhibited) if this area is illuminated This is the information passed to the visual cortex. There, individual cells (the first level of integration in the brain) sum the information coming from particular groups of ganglion cells. In the cat, the groups added formed bars (lines) of an ‘image’. Here’s what happens when a ‘bar’ of light shines onto the retina at different angles: There are two more (and more complex) levels of visual integration in the cat. You can see how some simple perceptions might occur: a “V” is two simple cells whose bars are at angles to each other summed at the next level… More complex objects and perceptions are harder to explain mechanically. They work, and we recognize objects as complex as the human face of someone familiar. This, as well as other aspects of biology, sometimes seem almost magical.