Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Killer-cell immunoglobulin-like receptor wikipedia , lookup
Hedgehog signaling pathway wikipedia , lookup
Programmed cell death wikipedia , lookup
Protein–protein interaction wikipedia , lookup
Cannabinoid receptor type 1 wikipedia , lookup
Leukotriene B4 receptor 2 wikipedia , lookup
Lipid signaling wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Biochemical cascade wikipedia , lookup
Toll-like receptor wikipedia , lookup
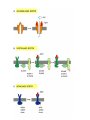
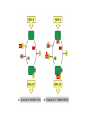
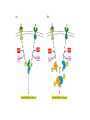
CELL SIGNALING Gap Junctions Allow Signaling Information to Be Shared by Neighboring Cells • These are specialized cell-cell junctions that can form between closely apposed plasma membranes, directly connecting the cytoplasms of the joined cells via narrow water-filled channels • The channels allow the exchange of small intracellular signaling molecules ( intracellular mediators), such as Ca2+ and cyclic AMP, but not of macromolecules, such as proteins or nucleic acids • Thus cells connected by gap junctions can communicate with each other directly without having to deal with the barrier presented by the plasma membranes Each Cell Is Programmed to Respond to Specific Combinations of Signaling Molecules • Any given cell in a multicellular organism is exposed to many perhaps hundreds - of different signals from its environment • Thus a cell may be programmed to respond to one set of signals by differentiating, to another set by proliferating, and to yet another by carrying out some specialized function • Most cells in higher animals, moreover, are programmed to depend on a specific set of signals simply for survival: when deprived of the appropriate signals a cell will activate a suicide program and kill itself - a process called programmed cell death • Different types of cells require different sets of survival signals and so are restricted to different environments in the body • Because signaling molecules generally act in combinations, an animal can control the behavior of its cells in highly specific ways using a limited diversity of such molecules: hundreds of such signals can be used in millions of combinations (Combinational Signaling) Different Cells Can Respond Differently to the Same Chemical Signal • TWO specific ways, a cell reacts to its environment 1. According to the set of receptor proteins that the cell possesses through which it is tuned to detect a particular subset of the available signals and, 2. According to the intracellular machinery by which the cell integrates and interprets the information that it receives • The neurotransmitter acetylcholine, for example, stimulates the contraction of skeletal muscle cells but decreases the rate and force of contraction in heart muscle cells • Difference may lie in different types of receptors and also different internal machinery (in case of identical receptors being used) The Concentration of a Molecule Can Be Adjusted Quickly Only If the Lifetime of the Molecule Is Short • It is important to consider what happens when a signal is withdrawn • Many intracellular proteins that are rapidly degraded have short half-lives, some surviving less than 10 minutes; in most cases these are proteins with key regulatory roles, whose concentrations are rapidly regulated in the cell by changes in their rates of synthesis • Likewise, any covalent modifications of proteins that occur as part of a rapid signaling process - most commonly the addition of a phosphate group to an amino acid side chain - must be continuously removed at a rapid rate to make such signaling possible Nitric Oxide Gas Signals by Binding Directly to an Enzyme Inside the Target Cell • Nitric oxide (NO), has recently been recognized to act as a signaling molecule in vertebrates • Acetylcholine acts indirectly by inducing the endothelial cells to make and release NO, which then signals the smooth muscle cells to relax ( nitroglycerine – treatment of angina) • NO is also produced as a local mediator by activated macrophages and neutrophils to help them kill invading microorganisms. • In addition, it is used by many types of nerve cells to signal neighboring cells • NO is made by the enzyme NO synthase by the deamination of the amino acid arginine. • Because it diffuses readily across membranes, the NO diffuses out of the cell where it is produced and passes directly into neighboring cells • It acts only locally because it has a short half-life about 5- 10 seconds • The effects of NO can be rapid, occurring within seconds • In many target cells, such as endothelial cells, NO reacts with iron in the active site of the enzyme guanylyl cyclase, stimulating it to produce the intracellular mediator cyclic GMP Steroid Hormones, Thyroid Hormones, Retinoids, and Vitamin D Bind to Intracellular Receptors That Are Ligand activated Gene Regulatory Proteins • Steroid hormones, thyroid hormones, retinoids, and vitamin D are small hydrophobic molecules that differ greatly from one another in both chemical structure and function • They diffuse directly across the plasma membrane of target cells and bind to intracellular receptor proteins. • Ligand binding activates the receptors, which then directly regulate the transcription of specific genes. • These receptors are structurally related and constitute the intracellular receptor superfamily (or steroid-hormone receptor superfamily) • Steroid hormones, including cortisol, the steroid sex hormones, vitamin D (in vertebrates), and the moulting hormone ecdysone (in insects), are all made from cholesterol • Besides the fundamental difference in the way they signal their target cells, most water-insoluble signaling molecules differ from water-soluble ones in the length of time that they persist in the bloodstream or tissue fluids. • Most water-soluble hormones are removed and/or broken down within minutes of entering the blood, and local mediators and neurotransmitters are removed from the extracellular space even faster - within seconds or milliseconds. • Steroid hormones, by contrast, persist in the blood for hours and thyroid hormones for days. • Consequently, water soluble signaling molecules usually mediate responses of short duration, whereas the water insoluble ones tend to mediate longer-lasting responses. • The intracellular receptors for the steroid and thyroid hormones, retinoids, and vitamin D all bind to specific DNA sequences adjacent to the genes that the ligand regulates • Ligand binding alters the conformation of the receptor protein, which then activates (or occasionally suppresses) gene transcription. • In many cases the response takes place in two steps: 1. The direct induction of the transcription of a small number of specific genes within about 30 minutes is known as the primary response; 2. The products of these genes in turn activate other genes and produce a delayed, secondary response. There Are Three Known Classes of Cell-Surface Receptor Proteins • Ion-channel-linked receptors, also known as transmitter-gated ion channels, are involved in rapid synaptic signaling between electrically excitable cells. • This type of signaling is mediated by a small number of neurotransmitters that transiently open or close the ion channel formed by the protein to which they bind, briefly changing the ion permeability of the plasma membrane and thereby the excitability of the postsynaptic cell. • The ion-channel linked receptors belong to a family of homologous, multipass transmembrane proteins • G-protein-linked receptors act indirectly to regulate the activity of a separate plasma-membrane bound target protein, which can be an enzyme or an ion channel. • The interaction between the receptor and the target protein is mediated by a third protein, called a trimeric GTP-binding regulatory protein (G protein) • The activation of the target protein either alters the concentration of one or more intracellular mediators (if the target protein is an enzyme) or alters the ion permeability of the plasma membrane (if the target protein is an ion channel). • The intracellular mediators act in turn to alter the behavior of yet other proteins in the cell. • All of the G protein- linked receptors belong to a large superfamily of homologous, seven-pass transmembrane proteins • Enzyme-linked receptors, when activated, either function directly as enzymes or are associated with enzymes • Most are single-pass transmembrane proteins, with their ligand binding site outside the cell and their catalytic site inside. • Compared with the other two classes, enzyme-linked receptors are heterogeneous, although the great majority are protein kinases, or are associated with protein kinases, that phosphorylate specific sets of proteins in the target cell Activated Cell-Surface Receptors Trigger PhosphateGroup Additions to a Network of Intracellular Proteins • Signals received at the surface of a cell by GPL and EL classes of receptors are often relayed to the nucleus, where they alter the expression of specific genes and thereby alter the behavior of the cell. • Elaborate sets of intracellular signaling proteins form the relay systems. The majority of these proteins are of one of two kinds: • Proteins that become phosphorylated by protein kinases, and • Proteins that are induced to bind GTP when the signal arrives. • In both cases the proteins gain one or more phosphates in their activated state and lose the phosphates when the signal decays • These proteins in turn generally cause the phosphorylation of downstream proteins as part of a phosphorylation cascade 1. Serine/threonine kinases, 2. Tyrosine kinases • It is estimated that about 1% of our genes encode protein kinases and that a single mammalian cell may contain more than 100 distinct kinds of these enzymes • Complex cell behaviors, such as survival or proliferation, are generally stimulated by specific combinations of signals rather than by a single signal acting alone • The cell has to integrate the information coming from separate signals so as to make a proper response to live or die, or to proliferate or stay quiescent. • The integration seems to depend on interactions between the various protein phosphorylation cascades that are activated by different extracellular signals SUMMARY Each cell in a multicellular animal is programmed during development to respond to a specific set of signals that act in various combinations to regulate the behavior of the cell and to determine whether the cell lives or dies and whether it proliferates or stays quiescent. Most of these signals mediate paracrine signaling, in which local mediators are rapidly taken up, destroyed, or immobilized, so that they act only on neighboring cells. In addition, centralized control is exerted both by endocrine signaling, in which hormones secreted by endocrine cells are carried in the blood to target cells throughout the body, and by synaptic signaling, in which neurotransmitters secreted by nerve cells act locally on the postsynaptic cells that their axons contact. Cell signaling requires both extracellular signaling molecules and a complementary set of receptor proteins in each cell that enable it to bind and respond to them in a programmed and characteristic way. Some small hydrophobic signaling molecules, including the steroid and thyroid hormones and the retinoids, diffuse across the plasma membrane of the target cell and activate intracellular receptor proteins, which directly regulate the transcription of specific genes. Some dissolved gases, such as nitric oxide and carbon monoxide, act as local mediators by diffusing across the plasma membrane of the target cell and activating an intracellular enzyme – usually guanylyl cyclase, which produces cyclic GMP in the target cell. But most extracellular signaling molecules are hydrophilic and are able to activate receptor proteins only on the surface of the target cell; these receptors act as signal transducers, converting the extracellular binding event into intracellular signals that alter the behavior of the target cell. There are three main families of cell-surface receptors, each of which transduces extracellular signals in a different way. Ionchannel- linked receptors are transmitter-gated ion channels that open or close briefly in response to the binding of a neurotransmitter. G-protein-linked receptors indirectly activate or inactivate plasma-membrane-bound enzymes or ion channels via trimeric GTP-binding proteins (G proteins). Enzymelinked receptors either act directly as enzymes or are associated with enzymes; the enzymes are usually protein kinases that phosphorylate specific proteins in the target cell. Through cascades of highly regulated protein phosphorylations, elaborate sets of interacting proteins relay most signals from the cell surface to the nucleus, thereby altering the cell's pattern of gene expression and, as a consequence, its behavior. Cross-talk between different signaling cascades enables a cell to integrate information from the multiple signals that it receives.