Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Biochemical switches in the cell cycle wikipedia , lookup
Tissue engineering wikipedia , lookup
Signal transduction wikipedia , lookup
Endomembrane system wikipedia , lookup
Cell encapsulation wikipedia , lookup
Extracellular matrix wikipedia , lookup
Programmed cell death wikipedia , lookup
Cell culture wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell growth wikipedia , lookup
Cytokinesis wikipedia , lookup
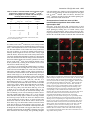
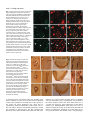
4435 Development 124, 4435-4446 (1997) Printed in Great Britain © The Company of Biologists Limited 1997 DEV8456 Only a subset of the binary cell fate decisions mediated by Numb/Notch signaling in Drosophila sensory organ lineage requires Suppressor of Hairless Shuwen Wang, Susan Younger-Shepherd, Lily Y. Jan and Yuh Nung Jan* Howard Hughes Medical Institute, and Departments of Physiology and Biochemistry, University of California, San Francisco, San Francisco, California 94143-0724, USA *Author for correspondence (e-mail: [email protected]) SUMMARY In Drosophila, an adult external sensory organ (bristle) consists of four distinct cells which arise from a sensory organ precursor cell via two rounds of asymmetric divisions. The sensory organ precursor cell first divides to generate two secondary precursor cells, IIa and IIb. The IIa cell then divides to produce the hair cell and the socket cell. Shortly after, the IIb cell divides to generate the neuron and the sheath cell. The membrane-associated protein Numb has been shown to be required for the first two asymmetric divisions. We now report that a new hypomorphic numb mutant not only displays a double-socket phenotype, due to a hair cell to socket cell transformation, but also a double-sheath phenotype, due to a neuron to sheath cell transformation. This provides direct evidence that numb functions in the neuron/sheath cell lineage as well. Those results, together with our observation from immunofluorescence analysis that Numb forms a crescent in the dividing IIa and IIb cells suggest that asymmetric localization of Numb is important for the cell fate determination in all three asymmetric cell divisions in the sensory organ lineage. Interestingly, we found that in the hair/socket cell lineage but not the neuron/sheath cell lineage, a Suppressor of Hairless mutation acts as a dominant suppressor of numb mutations whereas Hairless mutations act as enhancers of numb. Moreover, epistasis analysis indicates that Suppressor of Hairless acts downstream of numb, and results from in vitro binding analysis suggest that the genetic interaction between numb and Hairless may occur through direct protein-protein interaction. These studies reveal that Suppressor of Hairless is required for only a subset of the asymmetric divisions that depend on the function of numb and Notch. INTRODUCTION cell then divides to produce two outer support cells, the hair cell and the socket cell. Shortly after the IIa cell division, the IIb cell divides to produce two inner cells, the neuron and the sheath cell (Fig. 5A). Previous studies of the Drosophila embryonic and adult PNS suggest that the gene numb plays a key role in generating asymmetry in the SOP lineage (Uemura et al., 1989; Rhyu et al., 1994). Immunocytochemical studies suggest that Numb is a membrane-associated protein and is asymmetrically localized in the dividing SOP cell. Upon SOP division, Numb is preferentially segregated into the IIb cell (Rhyu et al., 1994). In numb loss-of-function mutants, the SOP cell divides symmetrically to produce two IIa cells, which produce four outer support cells (Uemura et al., 1989). Ectopic expression of Numb causes the opposite cell fate transformation in embryos (Rhyu et al., 1994). The generation of the four different cells of a sensory bristle in adult flies also requires the function of numb. Mosaic analysis reveals that the most extreme numb loss-of-function phenotype is the formation of four socket cells. This foursocket phenotype arises from the abnormal symmetric division of SOP to produce two IIa cells and the abnormal symmetric Asymmetric cell division is a fundamental process for the generation of daughter cells with different fates. Two types of mechanisms may be used in generating asymmetric cell divisions, an extrinsic mechanism and an intrinsic mechanism. The intrinsic mechanism involves the segregation of an inherited determinant preferentially into one of the daughter cells. The extrinsic mechanism may be executed through cellcell communication (reviewed by Horvitz and Herskowitz, 1992). In Drosophila, both mechanisms are used to generate the distinct fates of the cells within an adult sensory organ which, like the external sensory organ in the embryonic peripheral nervous system (PNS), is derived from a single sensory organ precursor (SOP) cell (Hartenstein and Posakony, 1989). A simple sensory organ comprises four cells with distinct fates: a neuron, a sheath cell, a socket cell and a hair cell (Fig. 1A). To generate these different cells of a sensory organ, the SOP cell follows a stereotyped pattern of asymmetric cell divisions (Hartenstein and Posakony, 1989). The SOP cell first divides to generate two secondary precursor cells, IIa and IIb. The IIa Key words: numb, Suppressor of Hairless, cell fate, neuron, sheath, sensory organ, Drosophila 4436 S. Wang and others Fig. 1. The numbSW allele exhibits double-socket phenotype in the adult sensory organs. (A) Schematic view of a sensory bristle structure. (B) Macrochaete and microchaete bristles from a wild-type fly notum. The anterior (aDC) and posterior (pDC) dorsal central macrochaete bristles are as indicated. (C) Macrochaete and microchaete bristles from a numbSW/numb2 fly notum. Note the double-sockets of one aDC and two pDCs as indicated by the black arrowheads. (D,E) Wing margin bristles from a wild-type and a numbSW/numb2 mutant fly, respectively. (F) Higher magnification of the boxed region in (C), showing a aDC double-socket (black arrowhead) and a microchaete double-socket (white arrowhead). division of each IIa cell to produce two socket cells (Rhyu et al., 1994). The transformation of IIb to IIa has therefore precluded the examination of the role of numb in the asymmetric division of the IIb cell in numb null mutants. Besides the cell-intrinsic mechanism mediated by numb, a cell-extrinsic mechanism mediated by Notch and Delta is also used for asymmetric divisions in the SOP lineage (Hartenstein and Posakony, 1990; Parks and Muskavitch, 1993). Analyses of temperature-sensitive mutants of Notch and Delta reveal that reduction of Notch or Delta function during the first SOP division causes both daughter cells to assume the IIb cell fate, which then divide symmetrically to produce four neurons (Hartenstein and Posakony, 1990; Parks and Muskavitch, 1993). In addition, Notch is also involved in both the IIa cell division and the IIb cell division, affecting the cell fates of both the socket/hair lineage and the neuron/sheath lineage (Guo et al., 1996). Notch and Delta probably mediate cell-cell communication through a receptor-ligand interaction in which Notch acts as the receptor and Delta as a ligand (Heitzler and Simpson, 1991; Rebay et al., 1991), and Numb acts by suppressing Notch activity during embryonic sensory organ development, probably via direct protein-protein interactions between Numb and Notch (Guo et al., 1996). In addition to their function in the lateral inhibition step during the SOP formation, the genes Suppressor of Hairless (Su(H)) and Hairless (H) help control the fates of adult sensory organ cells (Lees and Waddington, 1942; Bang et al., 1991; Bang and Posakony, 1992; Furukawa et al., 1992; Schweisguth and Posakony, 1992; Schweisguth, 1995). Su(H) is a potent dominant suppressor of H. Loss of function or overexpression of Su(H) during SOP divisions causes the IIa cell to divide symmetrically into two hair cells or two socket cells, respectively (Schweisguth and Posakony, 1992). Conversely, loss of function or overexpression of H exhibits a two-socket or twohair phenotype, respectively (Bang et al., 1991; Bang and Posakony, 1992). Shortly after the division of the IIa cell, the Su(H) protein specifically accumulates in the nucleus of the socket cell (Gho et al., 1996). These results suggest that Su(H) is required for socket cell formation and H is required for hair cell formation. There is growing evidence suggesting that Su(H) functions in the Notch signaling pathway. Both Su(H) and H show allelespecific genetic interactions with Notch and Delta during SOP formation (Vaessin et al., 1985; de la Concha et al., 1988; Fortini and Artavanis-Tsakonas, 1994). The Su(H) gene encodes a protein that is highly homologous to the mammalian RBP-J κ DNA-binding protein (Furukawa et al., 1992; Schweisguth and Posakony, 1992), and by binding specifically to sites in the promoter of the genes of the Enhancer of Split (E(spl)) complex, acts as a transcriptional activator of these genes (Bailey and Posakony, 1995; Lecourtis and Schweisguth, 1995). Activated Notch causes transcriptional activation of these E(spl) genes in transgenic flies, and this activation requires Su(H), providing further evidence for the participation of Su(H) in Notch signaling (Bailey and Posakony, 1995; Lecourtis and Schweisguth, 1995). A functional link between Su(H) and Notch has also been established in cultured Drosophila cells. When expressed in the S2 cells, the Su(H) protein is localized to the nucleus. Coexpression of Su(H) and Notch causes Su(H) protein to be retained in the cytoplasm, probably due to direct binding of Su(H) to Notch. Upon activation of Notch by its ligand Delta, the Su(H) protein translocates to the nucleus (Fortini and Artavanis-Tsakonas, 1994). This ability of activated Notch to cause translocation of Su(H) protein is inhibited by Numb (Frise et al., 1996). It has been proposed that Su(H) is involved in all three binary decisions in the sense organ lineage (Posakony, 1994; Schweisguth, 1995), implying that Notch signaling is transduced by Su(H) in all three binary decisions. However, Schweisguth and Posakony (1994) did not observe a sheath-toneuron transformation in flies carrying extra copies of Su(H). This result by itself does not prove but does raise the possibility that Su(H) is not required for the neuron/sheath cell fate decisions. It is therefore important to resolve whether Su(H) is indeed involved in all binary cell fate decisions mediated by Notch and numb. In this report, we first present the characterization of a hypomorphic mutant of numb, which exhibits both a hair-to-socket transformation and a neuron-to-sheath cell transformation. We then show that Su(H) is epistatic to numb and that a Su(H) Interaction of Su(H) with numb 4437 mutation acts as a dominant suppressor of the numb hypomorph while a H mutation acts as an enhancer of the numb hypomorph. Moreover, the interactions of Su(H) and H with numb only occur in the hair/socket cell lineage, indicating that Su(H) and H are not required in determining the neuron/sheath cell lineage. Further, in vitro binding analysis suggests that the genetic interaction between numb and H probably occurs through physical association of their protein products. MATERIALS AND METHODS Drosophila strains and genetics Drosophila strains were raised on standard cornmeal-yeast agar medium at 25°C or at room temperature. The three mutant alleles of numb (numb1, numb2, and numb3) and the deficiency that uncovers the numb locus (Df(2L)A-C) are described by Uemura et al. (1989). The null mutation of Su(H) (Su(H)SF8) was as described by Schweisguth and Posakony (1992). The four haploinsufficient alleles of H (HB55, HDEB1, HB521 and HBD9) were isolated and characterized by E. Grell (E. Grell, personal communication). The numbSW allele was isolated through an F1 revertant screen of the P-element-induced numb1 allele. Dysgenic males of the genotype numb1 /CyO; Dr ∆ 2-3/+ were crossed to numb2 pr cn Elp/CyO females. The F1 Cy+ progeny were collected and examined for abnormal sensory bristle phenotypes on the fly nota. Each candidate with abnormal bristles was retested for reproducible bristle defects by crossing to the other numb alleles. One mutation (numbSW allele) was isolated from screening 3600 F1 Cy+ progeny. This numbSW allele displayed a double-socket bristle phenotype when trans-heterozygous to other numb mutations. All four alleles of numb were balanced over the second chromosome balancer CyO. To examine the bristle phenotypes, numbSW /CyO flies were mated to numbX /CyO flies and the Cy+ flies were scored for sensory bristle defects (X represents 1, 2, 3, or the Df(2L)A-C). To study the inner cell fates of the numbSW mutant nota, we crossed homozygous numbSW flies to numb2 pr cn Bc/CyO flies. Third instar larvae that displayed the marker Black cell (Bc) were selected and maintained at room temperature or 25°C till the formation of white prepupae. After staging, the prepupae were dissected and stained with antibodies that recognize the inner cells of each sensory organ. For studying the bristle phenotypes caused by overexpression of numb, the homozygous UAS-numb#34 flies were crossed to the GAL4 enhancer-trap line Gal4109-68/CyO, and Cy+ flies were analyzed. Prepupae were collected from the same cross for pupal notum antibody staining of the inner cells of each sensory organ. The genetic interaction analysis between numbSW and Su(H)SF8 was carried out as follows. The numb2 allele and Su(H)SF8 allele were first recombined onto the same chromosome, generating flies of the genotype yw; numb2Su(H)SF8 /CyO p[y+, ry+]. Homozygous yw; numbSW flies were crossed to yw; numb2Su(H)SF8 /CyO p[y+, ry+] flies and Cy+ flies were examined for phenotypes. For inner sense organ cell analysis, third instar larvae with yellow mouth hooks (y−) were selected and staged to prepupae for pupal notum staining. Each of the four alleles of H was double balanced to numbSW using CyO and TM6 p[y+, ry+], generating flies of the genotype yw; numbSW /CyO; H/TM6 p[y+, ry+]. Females homozygous for yw; numbSW were then crossed to yw; numbSW /CyO; H/TM6 p[y+, ry+] males. Bristle phenotypes were scored in Cy+ and y flies (yw; numbSW ; H/+) as compared to Cy+ and y+ flies (yw; numbSW ; +/TM6 p[y+, ry+]). Double mutant mosaic analysis For the loss-of-function mitotic recombinant clones in adults and pupae, two different GAL4 enhancer trap lines, Gal4109-68 (Frise et al., 1996; Guo et al., 1996) and sca-Gal4 (Nakao and Campos-Ortega, 1996) were used to drive UAS-FLP recombinase expression in the sensory organ lineage. For adult notum yellow (y) and crinkled (ck) marked clones, we mated yw UAS-FLP; p[y+, ry+]VC1 ck FRT40A Gal4109-68/CyO females to yw; numb2 Su(H) SF8 FRT40A /CyO and yw; A1-2-29 Su(H) SF8 FRT40A /CyO males. Marked numb null mutant clones were generated by using two independent UAS-FLP transgenes: (1) p[UAS-FLP, w+] (a gift from J. Duffy, N. Perrimon and D. Harrison; Zeng et al. personal communication) was used by mating yw UAS-FLP; p[π M, w+] FRT40A to yw; p[y+, ry+] numb2 ck FRT40A /CyO; 2) p[UAS-FLP, ry+] (a gift from K. Bunier and K. Golic; Frise et al., 1996) was used by mating yw; p[π M, w+] FRT40A; UASFLP/TM6, p[y+, ry+] to yw; p[y+, ry+]VC1 ck numb2 FRT40A Gal4109-68/CyO. For clones in the pupae, we mated yw UAS-FLP; FRT40A sca-Gal4 (or Gal4109-68) females to the following males: yw; numb2 Su(H) SF8 FRT40A/ CyO, p[y+, ry+], yw ; numb2 ck FRT40A/CyO, p[y+, ry+] and yw; A1-2-29 Su(H)SF8 FRT40A /CyO, p[y+, ry+]. The fly stock, A1-2-29 Su(H) SF8 FRT40A (Gho et al., 1996) was used along with p[y+, ry+]VC1 numb2 ck FRT40A (Frise et al., 1996) to make the recombinant double and single mutant chromosomes: numb2 Su(H)SF8 FRT40A and numb2 ck FRT40A. To identify y− larvae that were FRT40A sca-Gal4 (or Gal4109-68) /numb2 Su(H)SF8 FRT40A, the p[y+, ry+]VC1 transgene (a gift from V. Corces) was jumped onto the CyO balancer (CyO, p[y+, ry+]) to serve as a marker for larval mouth hooks. Dissection of adult and pupal nota Adult nota were dissected in PBS, soaked in 80% isopropanol, and mounted in Hoeyer’s medium (Ashburner, 1989). White prepupae (time zero) were collected and maintained at 25°C for 24-30 hours (i.e. 24-30 hours after puparium formation (APF)) for cell fate transformation analysis. To examine the Numb protein localization, prepupae were maintained at 25°C for 14-16 hours APF, 16-18 hours APF, and 18-20 hours APF. Staged pupae were then dissected in PBS and fixed for 10 minutes in 5% formaldehyde in PEM solution (100 mM PIPES pH6.9, 1 mM MgCl2, 1 mM EGTA). Antibody staining and confocal microscopy The fixed pupal nota were washed in PBT (PBS containing 0.3% Triton X-100). The nota were incubated with primary antibodies for 2 hours at room temperature. After washing several times with PBT, the nota were incubated with the secondary antibodies conjugated with either fluorescein or rhodamine. The nota were washed first with PBT and then with PBS and subsequently mounted in 90% glycerol with 2% n-propyl-gallate. The antibodies used in this study have been described previously: the rabbit anti-Prospero antibody (Vaessin et al., 1991), the monoclonal anti-Elav antibody (mAb44C11) (Bier et al., 1988), the rabbit anti-Numb antibody (Rhyu et al., 1994), the rat antiCut antibody (Blochlinger et al., 1990), and the rat anti-Su(H) antibody (Gho et al., 1996), which were used at 1:1000, 1:5, 1:1000, 1:5000, and 1:1000 dilutions, respectively. The anti-Su(H) antibody was kindly provided by F. Schweisguth. A Zeiss microscope and a Bio-Rad MRC-600 confocal microscope were used to analyze the images. Generation of UAS-numb transformants The full length numb cDNA was derived from the KpnI fragment of hs-numb#2 (Rhyu et al., 1994) and subcloned into the KpnI site of the pUAST vector. The pUAST-numb DNA was introduced into w− embryos to generate transgenic flies by standard injection methods (Rubin and Spradling, 1982). The transgenic flies were crossed to scaGal4 (Hinz et al., 1994) or Gal4109-68 flies (Frise et al., 1996) for phenotypic characterization. Fifteen out of 18 independent transformants displayed similar phenotypes, i.e. balding and twinned hairs, although the strength of the phenotypes varied from one tranformant to another. #34 of pUAST-numb (on the third chromosome) was used for the studies presented in this report. In vitro binding assays The Notch intracellular domain and various Numb fragments were 4438 S. Wang and others cloned in-frame into the pGEX vectors (Pharmacia) for expression in DH5α as GST-fusion proteins. The expression of fusion proteins was induced for 3 hours with 1 mM isopropyl-D-thiogalactopyranoside added to log-phase bacterial cultures. Following sonication of the bacteria in PBS, Triton X-100 was added to a final concentration of 1%. The bacterial lysate was then centrifuged at 7,000 rpm (9000 g) for 10 minutes in an HS-4 rotor in a Sorvall centrifuge. The supernatant was then mixed with glutathione Sepharose-4B beads (Pharmacia) at 4°C for 30 minutes. The beads were then washed three times in PBS/1% Triton X-100 and three times in PBS/0.1% NP-40. The washed beads were kept at 4°C as a 50% suspension until use. The plasmid pNB40-H, which contains the full length H cDNA, was kindly provided by J. Posakony. [35S]Met-labeled H protein was expressed from pNB40-H using the TNT-coupled in vitro transcription/translation rabbit reticulocyte system (Promega). For in vitro binding analyses, 20 µl of the 50% suspension of beads (with bound GST fusion protein) were mixed with 5 µl lysate of the [35S]Metlabeled in vitro translated protein. The protein mixture was incubated in 150 µl PBS/0.1% NP-40 at 4°C for 30 minutes and the beads were then washed 4-5 times with PBS/0.1% NP40. After washing, SDS-PAGE loading buffer was added, the samples were boiled, and the supernatant was analyzed by 10% SDS-PAGE (Sambrook et al., 1989). The gels were dried and subjected to autoradiography. RESULTS Reduction of numb function causes both hair to socket and neuron to sheath transformations The three previously isolated numb alleles (numb1, numb2 and numb3) cause recessive embryonic lethality, rendering these mutants unsuitable for the study of interactions of numb with other genes during adult sensory organ formation. We have therefore carried out a P-element revertant screen in order to isolate more numb mutants that are viable and show visible phenotypes in adult sensory bristles. We started with the allele numb1, which was generated by insertion of the transposon pUChsneo (see Materials and Methods for details), and isolated a viable hypomorphic numb allele (numbSW) which exhibits a double-socket phenotype (Fig. 1). The sensory organs on the fly notum (dorsal thorax) include both macrochaetae and microchaetae, large and small bristles, positioned in a fixed pattern. Each bristle has an external sensory structure that is composed of a hair and a socket (Fig. 1A). In the numbSW mutant flies, both macrochaetae and microchaetae exhibit abnormal external sensory structures (Fig. 1C,F). Double-socket bristles are produced at the expense of a hair in these partial loss of numb function mutants. The severity of the double-socket bristle phenotype varies with the genotype (Table 1). The homozygous numbSW flies show a small number of double-socket bristles (3 out of 26 macrochaetae on the notum), while transheterozygous numbSW/numb2 flies display a very strong double-socket phenotype (12 out of 26 macrochaetae). These abnormal bristles with double sockets are observed on almost every part of the fly, including notum, head, wing (Fig. 1E), abdomen, legs and sex combs. This double-socket phenotype can be rescued by overexpression of numb driven by the heat-shock hsp70 promoter (data not shown), suggesting that this new mutation is indeed a numb allele. The double-socket phenotype generated in numbSW flies is similar to the double-socket phenotype generated in some of the clones that have lost numb Table 1. Numbers of double-socket macrochaetae in flies that are trans-heterozygous for numbSW and various numb alleles, as compared to those of the wild-type (+/+) flies Average number of double sockets Genotypes numbSW/numb1 numbSW/numb2 numbSW/numb3 numbSW/numbSW numbSW/Df(2L)A-C Notum Head 11.2 12.4 11.8 3.0 15.4 7.9 10.2 9.2 1.9 11.3 Wild-type macrochaetae number: notum, 26; head, 14. function (Rhyu et al., 1994) and confirms the function of numb in the hair/socket lineage. Besides a hair and a socket, a normal sensory bristle also contains two inner cells, a neuron and a sheath cell (Fig. 1A). To examine the inner cells of sensory bristles in the numbSW mutant we have used a neuronal nuclear marker, the monoclonal antibody mAb44C11 (an anti-Elav antibody), and a sheath cell nuclear marker, the anti-Prospero antibody, and doubly stained numbSW mutant nota at 24-30 hours APF when all four cells of a sensory organ are fully differentiated. In wildtype nota, each sensory bristle has one mAb44C11-positive neuron and one Prospero-expressing sheath cell (Fig. 2A-C). Fig. 2. Partial loss of function of numb transforms the neuron into a sheath in an adult sensory organ. Pupal nota at 24-30 hour APF were dissected and doubly stained with the neuronal marker mAb44C11 (red) and the sheath cell marker anti-Prospero antibody (green). (A-C) Wild-type pupal notum. Each sensory organ contains one sheath cell (A, green) and one neuron (B, red) that are positioned next to each other (C). (D-F) A numbSW/numb2 mutant pupal notum. Note the double-sheath cells (marked by *), the two green cells that are positioned next to each other (D), are sensory organs in which the neuron is transformed into a sheath cell. Interaction of Su(H) with numb 4439 Table 2. Numbers of double-sheath cells in numbSW/numb2 mutant nota compared to those in numbSW + /numb2 Su(H)SF8, from pupal nota stained with anti-Prospero antibody and mAb44C11 antibody Pupa no. 1 Pros+ cell A 1 2 3 4 numbSW/numb2 B 1 2 3 numbSW+/numb2Su(H)SF8 9 8 10 9 8 4 7 2 Pros+ cells Total macrochaetae counted 9 10 9 11 18 18 13 18 10 13 10 19 21 20 cells with distinct fates. Similar crescent localization of Numb was also observed during the SOP development on the pupal nota of numbSW mutants (data not shown). Therefore, the numbSW mutation most likely affects the Numb signaling efficiency rather than Numb localization. Overexpression of numb can cause cell fate transformations opposite to those of the numb hypomorphic allele Previously, it has been shown that overexpression of numb using the heat-shock hsp70 promoter (hs-numb) results in two major phenotypes, balding (loss of external sensory organ structures) and twinned hairs with no socket. These two phenotypes presumably are due to the transformation of the IIa cell into the IIb cell and the socket cell into the hair cell in the IIa Average neuron to sheath transformation: numbSW/numb2, 58%; numbSW+/numb2Su(H)SF8, 55%. In contrast, in the numbSW mutant nota, some sensory bristles contain two Prospero-positive sheath cells but no neurons (Fig. 2D-F), indicating that the neuron is transformed into a sheath cell. The total number of cells within each sensory bristle does not seem to change, as revealed by staining with an anti-Cut antibody which recognizes all four cells of the sensory organ (data not shown). About 50%-60% of the sensory bristles have a neuron to sheath cell transformation (Table 2A), showing that numb also functions in the IIb cell lineage for the formation of the neuron and the sheath cell. Therefore, partial loss of numb activity in the numbSW allele causes both IIa and IIb cells to divide symmetrically, resulting in the formation of two socket cells and two sheath cells (Fig. 5B). Previously it was shown that the Numb protein is asymmetrically localized in the dividing SOP cell, forming a crescent in the cell cortex of SOP (Rhyu et al., 1994). We were therefore interested in knowing whether Numb protein localization is affected in the numbSW mutants. Since the Numb protein localization in later stages (i.e. the IIa and IIB divisions) during the SOP development has not been studied, we have first examined the Numb protein localization on the pupal nota of wild type during each of the divisions (SOP, IIa, and IIb) of a microchaete bristle (Fig. 3). In this experiment, we used the anti-Cut antibody which stains the nuclei of all the cells within each sensory organs, and the anti-Numb antibody, and doubly stained pupal nota at 14-16 hours APF (Fig. 3A-C), 16-18 hours APF (Fig. 3D-F), and 18-20 hours APF (Fig. 3G-I). On a notum at 14-16 hours APF when dividing SOPs are frequently present as revealed by the non-nuclear staining of the anti-Cut antibody, a clear crescent staining of Numb is associated with each dividing SOP cell (Fig. 3A-C). At 16-18 hours APF, each adult sensory organ is at the two-cell stage, with the IIa cell being ready to divide. We observed a Numb crescent in each dividing IIa cell (Fig. 3D-F). A couple of hours after the division of the IIa cell, the IIb cell starts to divide (18-20 hours APF). Fig. 3G-I shows that a Numb crescent also forms in the IIb cell before it divides during a three-cell stage. The observation of asymmetric localization of Numb protein in the dividing SOP, IIa cell and IIb cell suggests that Numb is preferentially segregated into one daughter cell during each of the three divisions in the SOP lineage, thus generating daughter Fig. 3. Numb is asymmetrically localized in all three divisions during SOP development. Pupal nota at 14-16 hours APF, 16-18 hours APF, and 18-20 hours AFP were dissected and doubly stained with the anti-Cut (red) and the anti-Numb (green) antibodies. Similar results were obtained with both wild-type and numbSW mutant pupal nota. (A-C) A region of pupal notum at 14-16 hours APF. Note that a Numb crescent (arrow) is formed during mitosis of a SOP cell (as revealed by the cytoplasmic staining of anti-Cut antibody). The arrowhead shows the two Cut-positive cells with nuclear staining, corresponding to a IIa cell and a IIb cell resulted from a SOP division. (D-F) A region of pupal notum at 16-18 hours APF. Arrow indicates the Numb crescent. Note that the Numb crescent is associated with the dividing cell, the IIa cell, whereas the neighboring cell with nuclear staining of Cut is presumably the IIb cell. The arrowhead shows a two-cell cluster positive for nuclear staining of Cut, corresponding to a stage before the IIa cell enters mitosis. (G-I) A region of pupal notum at 18-20 hours APF. Arrow indicates the Numb crescent, which is associated with the IIb cell during mitosis. The arrowhead shows the two cells next to the IIb cell which are positive for nuclear staining of Cut. These two neighboring cells are presumably the hair cell and the socket cell. Brackets show two clusters of three cells with nuclear staining of Cut, suggesting that the IIb cells in these three-cell clusters are at a stage before entering mitosis. 4440 S. Wang and others ments confirm the finding that numb functions in all three divisions of the SOP lineage. Fig. 5 summarizes the cell fate transformations in the IIa and IIb lineage within an adult sensory organ in numbSW and UAS-numb, as compared to the wild type. Fig. 4. Overexpression of numb can generate phenotypes that are opposite to those from reduction of numb function. The homozygous UAS-numb (#34A) flies were crossed to Gal4109-68 and the Gal4109-68/+; UAS-numb/+ flies were examined. (A,C) Twinned hairs caused by the socket to hair transformation. (B,D) Balding phenotype due to the transformation from IIa into IIb. (E-G) A pupal notum of Gal4109-68/+; UAS-numb/+ mutant at 24-30 hour APF doubly labeled with the anti-Prospero antibody (E, green) and the mAb44C11 (F, red). The three neurons and one sheath cell phenotype is marked by arrowhead, two neurons and two sheath cells by *s, and two neuron with no sheath cells by arrows. A Su(H) mutation acts as a dominant suppressor of the numb hypomorph in determining the hair/socket lineage but not the neuron/sheath lineage Partial reduction of Su(H) activity produces a twinned hair phenotype, resulting from transformation of the socket cell into a hair cell (Schweisguth and Posakony, 1994), opposite to the phenotype caused by decreased numb function. Conversely, overexpression of Su(H) causes a double-socket phenotype, presumably resulting from transformation of the hair cell into a socket cell (Schweisguth and Posakony, 1994). To test whether these genes act in the same pathway, we assessed the influence of the Su(H) null allele, Su(H)SF8, on the hypomorphic allele numbSW. As described above, numbSW /numb2 flies showed the double-socket phenotype in about 50% of the adult sensory bristles (Table 1) and a neuron to sheath transformation in 50-60% of pupal sensory bristles (Table 2A). Removing one copy of the Su(H) gene in numbSW /numb2 flies (i.e. numbSW + /numb2 Su(H)SF8) strongly suppressed the doublesocket phenotype (Fig. 6A-C). In contrast, reduction of Su(H) function had no effect on the numbSW double-sheath phenotype. As shown in Fig. 6D-F and summarized in Table 2B, double-labeling of the sensory bristle cells with mAb44C11 and anti-Prospero clearly showed the doublesheath cell phenotype characteristic of numbSW /numb2 mutant. These results establish that Su(H)SF8 behaves as a dominant suppressor of the hair cell to socket cell transformation but not of the neuron to sheath cell transformation in the numbSW/numb2 mutant. neuron neuron hair hair sheath sheath socket socket sheath neuron socket hair Numb determines the hair cell identity by negatively regulating Su(H) protein expression cell lineage, respectively (Rhyu et al., 1994). We examined the effects of numb overexpression on the cell fate of the inner The strong genetic interaction betwen numbSW and Su(H)SF8 cells, the neuron and sheath cell (daughters of the IIb cell). made us wonder how numb and Su(H) act antagonistically. The UAS-GAL4 system (Brand and Perrimon, 1993) was Shortly after division of the IIa cell, the Su(H) protein is used to study the effect of numb overexpression. Transgenic flies carrying one copy of UAS-numb A B C (#34A) were crossed to flies of a GAL4 enhancer-trap line, Gal4109-68, which drives the transgene expression SW Wild type numb UAS-numb in the SOP cell and its progeny. Two phenotypes resulted from such overexpression of numb, twinned hairs (Fig. 4A,C) and balding (Fig. 4B,D), the same SOP SOP SOP as the phenotypes caused by hs-numb (Rhyu et al., 1994). Sensory bristles in the dissected pupal nota of flies carrying both UAS-numb and Gal4109-68 were IIa IIb IIa IIb IIa IIb doubly stained using the neuronal marker mAb44C11 and the sheath cell marker anti-Prospero antibody. Four types of phenotypes were observed for the inner cells: four neurons, three neurons and one sheath cell, two neurons and two sheath cells, and two neurons with no sheath cells (Fig. 4E-G). Most likely, transformation of both the IIa cell to a IIb cell and the sheath to a neuron caused these phenotypes. Therefore, overexpression of numb can cause phenoFig. 5. Schematic diagrams of the SOP cell lineage for (A) the wild type, types opposite to the loss or reduction of numb (B) the hypomorph numbSW, and (C) the overexpression of numb (UASnumb). function phenotypes. These overexpression experi- Interaction of Su(H) with numb 4441 activity. Numb may normally be present primarily in the hair cell but not in the socket cell, thus resulting in low Su(H) protein level in the hair cell and high Su(H) protein level in the socket cell. When numb function is reduced, the level of Su(H) protein is increased in the hair cell, leading to the transformation of the hair cell into a socket cell. Conversely, when numb function is increased, the level of Su(H) protein is suppressed in the socket cell, resulting in the transformation of the socket cell into a hair cell. Fig. 6. Reducing one copy of Su(H) suppresses the double-socket phenotype but not the double-sheath phenotype of numbSW/numb2 mutants. (A) An adult notum from a numbSW + /numb2 Su(H)SF8 fly showing the normal looking macrochaete and microchaete bristles. This indicates that the double-socket phenotype generated in the numb hypomorph is suppressed by removing one copy of the wildtype Su(H) gene. (B) The dorsal central region of the notum in (A) at higher magnification. (C) The wing margin bristles from a numbSW + /numb2Su(H)SF8 fly showing a similar suppression of the double socket bristle phenotype. (D-F) Staining of a pupal notum from numbSW + /numb2Su(H)SF8 mutants with the anti-Prospero antibody (D, green) and mAb44C11 antibody (E, red). Note that the doublesheath cells (marked by *) are still present, indicating that the suppression of the numbSW/numb2 phenotype by Su(H)SF8 is restricted to the socket/hair lineage. present in the nucleus of the socket cell but not the hair cell (Gho et al., 1996), and it is required for socket cell formation (Schweisguth and Posakony, 1994). We have examined Su(H) protein expression in flies with partial reduction of numb activity (numbSW mutant flies) and in flies with increased numb activity (UAS-numb/Gal4109-68). Within each sensory bristle of the wild-type pupal nota, a single sheath cell is labeled with the anti-Prospero antibody and a single socket cell is labeled with the anti-Su(H) antibody (Fig. 7A-C). Similar immunofluorescence staining in the nota of the numbSW/numb2 mutants showed a number of sensory organs with two cells expressing Su(H) (Fig. 7D-F), consistent with the double-socket phenotype (Fig. 1C,F). Moreover, in pupae where Numb is overexpressed (UAS-numb/Gal4109-68), some sensory organ cells have greatly reduced Su(H) expression (Fig. 7G-I), consistent with the twinned hair phenotype (Fig. 4A,C). These results suggest that numb may determine the fate of the hair cell and the socket cell by controlling the level of Su(H) Epistatic relationship between Su(H) and numb The finding that loss-of-function mutations of numb and Su(H) have opposite effects on the hair/socket cell fate specification and the strong genetic interaction between these two genes raise the question of their epistatic relationship. To investigate this issue, we used UAS-FLP and specific GAL4 enhancer trap lines to target FRT mediated clones of both numb and Su(H) to the SOP lineage (Frise et al., 1996) (Zeng et al., personal communication). We took advantage of two GAL4 enhancertrap lines, Gal4109-68 and sca-Gal4, which express GAL4 specifically in the SOP and its daughter cells. Both Gal4109-68 and sca-Gal4 show similar expression of a reporter gene during SOP divisions, although the expression in sca-Gal4 appears to be stronger. The reporter gene expression in sca-Gal4 also starts earlier in proneural clusters before the SOP is singled out. We expected that the FLP recombinase would be expressed in a similar pattern from the UAS-FLP transgene. The SOP lineage specific FLP expression induced mitotic recombination between a FRT chromosome carrying mutations of numb, Su(H), or numb Su(H) and a ‘wild-type’ FRT chromosome, producing one daughter cell homozygous for the null mutations and the other daughter cell homozygous for the wildtype genes. Loss of numb function in the IIa cell due to UAS-FLP and Gal4109-68 caused a double-socket phenotype (Fig. 8A-C), which is indistinguishable from the double sockets observed in the numbSW allele (Fig. 1C,F) or in heat-shock FLP-numb FRT mutant clones generated during third instar larval and pupal stage (Rhyu et al., 1994). Interestingly, sensory organs with three sockets and one hair (Fig. 8D) or four sockets (data not shown) were observed with a moderate frequency when clones for the null mutation of numb were generated using the GAL4 line sca-Gal4. These phenotypes are most likely due to an early mitotic recombination event induced by the early expression of sca-Gal4 in the proneural cluster before the division that gives rise to the SOP cells. In this case, the SOP itself would be homozygous for the numb null mutation, causing transformation of the IIb cell into the IIa cell. Subsequently, loss of numb function in the two IIa cells may cause one IIa cell to divide symmetrically to form two socket cells, whereas the other IIa cell may either give rise to a hair cell and a socket cell due to an incomplete transformation or two socket cells due to a complete transformation. Twinned hairs as well as several other bristle phenotypes were observed when either Gal4109-68 or sca-Gal4 was used to remove Su(H) function in the SOP cell and its daughter cells (Fig. 8E-I). The twinned hair phenotype suggests that Su(H) functions in the IIa division to determine the socket cell fate (Schweisguth and Posakony, 1994). Loss of Su(H) function, like that of numb, also yields a broad spectrum of transformations. The phenotypes are always in the direction 4442 S. Wang and others Fig. 7. Su(H) protein expression in various numb mutant background. Pupal nota at 24-30 hour APF were dissected and doubly stained with the anti-Su(H) antibody (B,E,H, red) and the antiProspero antibody (A,D,G, green). (A-C) A notum from a wild-type pupa. Note that a Su(H)-expressing cell (the socket cell, red) is associated with a Prospero-expressing cell (the sheath cell, green). (D-F) A notum from a numbSW/numb2 mutant. Note that three phenotypes are generated: two Prosperoexpressing cells and two Su(H)-expressing cells (i.e. 2 sheath cells and 2 socket cells, *); two Prospero-expressing cells and one Su(H)expressing cell (i.e. 2 sheath cells and 1 socket cell, arrows); and one Prospero-expressing cell and two Su(H)-expressing cells (i.e. 1 sheath cell and 2 socket cells, not shown in the figure). (G-I) A notum from a Gal4109-68/+; UASnumb/+ pupa. Note the disappearance of the Su(H)-expressing cell in some sensory organs. Two phenotypes are associated with this absence of Su(H) protein expression: the presence (small white arrowheads) or the absence (large white arrowheads) of a Prospero positive cell. These two phenotypes are presumably due to the transformation from socket to hair and from sheath to neuron, respectively. Fig. 8. The double mutants of Su(H) and numb showed a Su(H) mutant phenotype, indicating that Su(H) acts downstream of numb. (A-C) Adult nota (A,B) with a scutellum (C) from the numb2 null mutant clones generated by Gal4109-68. (D) A notum from the numb2 null mutant clones generated by sca-Gal4. Note that a three socket and one hair phenotype (arrowhead) is also generated in addition to the double-socket phenotype. (E-I) Su(H)SF8 mutant clones showing the twinned hairs in nota (H,I) and scutellum (G) as well as the intermediate phenotypes caused by partial transformation of socket into hair (E,F). (J-N) The Su(H) and numb double mutant clones showing twinned hairs in both nota (M,N) and scutellum (L) as well as the intermediate phenotypes caused by partial transformation of socket into hair (J,K). Note that the various phenotypes generated in the double mutants are similar to those generated in the Su(H) null mutants. of forming two hair cells and no socket cell, and they range from a remnant of a hair cell associated with an abnormal socket to the formation of twinned hairs at the expense of the socket cell. These phenotypes (Fig. 8E,F) may be explained by partial transformation of the socket cell to a hair cell and are therefore intermediate phenotypes toward the formation of twinned hairs. We then examined the phenotype in the double mutants where both numb and Su(H) function were removed from the SOP and/or its daughter cells. We found that a range of phenotypes was produced in the numb2 Su(H)SF8 mosaic flies when either Gal4109-68 or sca-Gal4 was used (Fig. 8J-N). Phenotypes of the double mutants were indistinguishable from those due to loss of function of Su(H) alone (Fig. 8E-I). Heat-shock FLP induced mitotic recombination, generated in second instar larvae, also produced double null mutant clones with phenotypes Interaction of Su(H) with numb 4443 Fig. 9. The epistatic relationship between numb and Su(H) does not apply to the neuron/sheath cell lineage. Mutants of numb or Su(H) and double mutants of numb and Su(H) were generated by scaGal4. Pupal notum from numb mutant (A-C) or Su(H) mutant (D-F) or double mutant of numb and Su(H) (G-I) were doubly stained with antiProspero antibody (left panels, green) and mAb44C11 (middle panels, red). Note the presence of the double-sheath phenotypes (*s) in the numb mutant and double mutants but not in the Su(H) mutant. indistinguishable from those of Su(H) null mutant clones (data not shown). Therefore, the Su(H) gene appears to act downstream of numb in the same genetic pathway in determining the fates of the IIa daughter cells, the hair cell and the socket cell. We have further examined the inner cells on the pupal nota of these mutants at 24-30 hours APF by double staining with the anti-Prospero antibody and the monoclonal antibody mAb44C11 which specifically stain the sheath cells and the neurons, respectively (Fig. 9). As expected, the loss-offunction mutation of numb caused a double-sheath phenotype (Fig. 9A-C). In loss-of-function clones of Su(H), the sheath/neuron cell lineage is completely normal (Fig. 9D-F). In the double mutants where null mutations of both numb and Su(H) were generated in the SOP lineage, the double-sheath phenotype due to loss of numb function did not seem to be affected by the removal of the Su(H) function (Fig. 9G-I). Similar phenotypes of the inner cells in these double mutants were generated when either Gal4109-68 or sca-Gal4 was used. These results provide further evidence for Su(H) acting as a downstream target of numb only in determining the cell fates in the IIa lineage for the formation of the hair cell and the socket cell. Consistent with this scenario, overexpression of Su(H) via the heat-shock promoter (hs-Su(H)) did not affect the neuron/sheath cell lineage, as revealed by double staining with the anti-Prospero antibody and the mAb44C11 antibody (data not shown). Our results strongly indicate that Su(H) and numb function in the same genetic pathway, with Su(H) acting downstream of numb and negatively regulated by numb. However, this interaction is restricted to the hair/socket lineage and there is no such epistatic relationship in the Table 3. Numbers of the double-socket cells counted in numbSW nota compared to those in numbSW; H/+ nota Notum macrochaetae (number of double sockets) Genotype HB55 HDEB1 HB21 HD9 numbSW/numbSW; 12.1 13.0 11.7 7.1 H/+ numbSW/numbSW; +/+ numbSW/+; H/+ 2.0 2.8 2.5 2.7 4.2 2.1 2.7 0.7 The wild-type notum contains 26 macrochaetae. neuron/sheath cell lineage where apparently Su(H) is not required. H acts as a dominant enhancer of numb in determining the hair/socket lineage Previously, it has been shown that Su(H) and H act antagonistically to control the cell fates in the adult sensory organs (Schweisguth and Posakony, 1994). The Su(H) mutation acts as a dominant suppressor of the various haplo-insufficient alleles of H to suppress the double-socket phenotype of H mutants. The double-socket phenotype exhibited by H lossof-function mutants is similar to that of the numb loss-offunction phenotype. Given that numb and Su(H) act in the same genetic pathway, we have further examined the genetic interaction of H with numb. Homozygous numbSW flies display a very weak double-socket phenotype (middle column in Table 3; Fig. 10A,D). Removing one copy of H greatly enhances the double-socket phenotype of the numbSW flies (Fig. 10C,F). Table 3 summarizes the effect of four different H alleles on the numbSW flies. We found that the 4444 S. Wang and others Fig. 10. Partial loss of function of H enhances the double-socket phenotype in homozygous numbSW flies. (A,D) A notum and a wing margin from numbSW flies, showing the weak (low frequency) doublesocket phenotype. (B,E) A notum and a wing margin from a HB21/+ fly showing the fairly normal bristles on both the notum and the wing margin. (C,F) A notum and a wing margin from a numbSW; HB21/+ fly, showing the greatly enhanced double-socket phenotype. number of double-sockets in numbSW mutants carrying one copy of the H mutation is two- to three-fold greater than the sum of those in H and numbSW single mutants (Table 3). Thus the hair/socket cell lineage is very sensitive to the dosage of both H and numb. H mutations function as dominant enhancers of numbSW in the specification of cell fates of the hair/socket cell lineage of adult sensory bristles. In a genetic screen for modifiers of numb using the numbSW allele, we isolated a dominant enhancer of numbSW (data not shown). This enhancer mutant was subsequently found to be allelic to H, further substantiating the genetic interaction we observed between numb and H (Fig. 10 and Table 3). H and Numb physically interact with each other Given that numb, Notch, Su(H), and H all participate in the same genetic pathway in determining the IIa cell lineage and that Su(H) binds to H (Bailey and Posakony, 1995), Notch binds to Su(H) (Fortini and Artavanis-Tsakonas, 1994), and Numb binds to Notch (Guo et al., 1996), we wondered whether the interactions of numb with Su(H) and H are due to physical associations of their gene products. We performed an in vitro binding assay to determine whether Numb binds to Su(H) or H. As shown in Fig. 11, a bacterial fusion protein, GST-Numb, and in vitro translated H protein were coprecipitated by glutathione agarose beads (Fig. 11, lane 2). Similar coprecipitation of GST-Notch and the in vitro translated H protein was also observed (Fig. 11, lane 1). The in vitro translated H protein binds the Numb-N fragment which encompasses the PTB domain of Numb (Fig. 11, lane 3), whereas the GST-Numb-C (Fig. 11, lane 4) or GST alone (Fig. 11, lane 5) failed to bind to H. Similar binding analysis with the in vitro translated Su(H) protein showed no detectable binding of Su(H) to Numb (data not shown). Fig. 11. The in vitro binding of Numb and Notch to H. In vitro translated 35S-labeled full length H binds to GST-Numb (lane 2) and GST-Numb-N (lanes 3), but not GST-Numb C (lane 4) or GST alone (lane 5). In addition, the in vitro translated 35S-labeled H also binds to GST-Notch intra (lane 1). DISCUSSION By isolating and characterizing a hypomorphic numb mutant, numbSW, we have confirmed and extended previous studies to show that numb functions in all three asymmetric divisions that give rise to a simple external sensory organ. Moreover, we found that the IIa lineage which gives rise to the hair and the socket, but not the IIb lineage which gives rise to the neuron and the sheath cell, requires the functions of Su(H) and presumably H. These findings, and the genetic interactions among these genes, are discussed below. numb functions in all three asymmetric cell divisions during the formation of adult sensory organs Studies of numb in embryos indicate that numb functions during the SOP division to produce the two distinct secondary precursors, the IIa cell and the IIb cell (Uemura et al., 1989; Rhyu et al., 1994). Moreover, results from overexpression and mosaic analysis in adult flies suggest that numb functions during both SOP division and the IIa division in the development of adult sensory organs (Rhyu et al., 1994). Therefore, numb plays an important role in specifying the IIb cell fate and the hair cell fate. Although it was proposed previously that numb also plays a role in specifying the progeny cell fates of IIb, it has not been demonstrated directly owing to the transformation of the IIb cell into the IIa cell in numb null mutants. The characterization of the newly isolated hypomorphic allele of numb (numbSW) clearly indicates that numb is also involved in the sheath/neuron lineage (the IIb cell lineage), as revealed by the double-sheath cell phenotype on fly pupal nota as revealed by staining with anti-Prospero antibody (Fig. 2). Similar double-sheath phenotypes were also observed in the external sensory organs in fly embryos (data not shown). This involvement of numb in the choice between the neuron and the sheath cell fates was also confirmed by mosaic analysis using the UAS-FLP/FRT with GAL4 and a numb null allele (numb2) (Fig. 9). In addition, we have performed immunofluorescence analysis and found that the Numb protein is asymmetrically localized in the dividing SOP, the dividing IIa cell, and the dividing IIb cell (Fig. 3). Thus, Numb functions to promote the IIb cell fate in the SOP division, the hair cell fate in the IIa lineage, as well as the neuron fate in the IIb lineage presumably by being preferentially segregated into the IIb cell, the hair cell, and the neuron, respectively. The normal presence of asymmetric Numb localization in numbSW mutants suggests that the hypomorphic mutation of numb may affect the Numb signaling efficiency. Interaction of Su(H) with numb 4445 Su(H) and H are required for determining the hair/socket cell lineage but not the neuron/sheath cell lineage It has previously been shown that Su(H) and H are required for the determination of the socket cell and the hair cell within an adult sensory organ, respectively (Schweisguth and Posakony, 1994). Loss of function alleles of Su(H) are potent dominant suppressors of the double-socket phenotype in H mutants (Schweisguth and Posakony, 1992), suggesting that reduction of Su(H) activity favors the hair cell fate. Consistent with this hypothesis, a twinned hair phenotype without a socket was produced by either partial loss of function or the null mutant of Su(H) in mosaic analysis (Schweisguth and Posakony, 1994). Overexpression of Su(H) produces the opposite transformation. When Su(H) activity is increased during the IIa cell division, the hair cell is transformed into a socket cell, resulting in a double-socket phenotype (Schweisguth and Posakony, 1994). In contrast, reduction of function or overexpression of H during the later stage of the SOP lineage yields a doublesocket or twinned hair phenotype, respectively (Bang et al., 1991; Bang and Posakony, 1992). Therefore, unlike Su(H), which is required for socket cell formation, H activity seems to be required for hair cell formation. Although it was proposed that the Su(H) and H activities are required for all three binary cell fate decisions in sense organ development (Posakony, 1994), Schweisguth and Posakony (1994) observed that flies which carry extra copies of Su(H) contain a single neuron within each sensory organ, as revealed by staining with the anti-HRP antibody. This observation raised the possibility but did not prove that Su(H) is not involved in the neuron/sheath cell fate decision because it was not known whether the extra copies of Su(H) caused overexpression of Su(H) at the right place and right time to produce sheath/neuron transformation. Importantly, whether neuron/sheath decision is affected by Su(H) loss-of-function mutant was not tested. Here, we have used anti-Prospero antibody, which labels the nuclei of sheath cells, and the monoclonal antibody mAb44C11, which labels the nuclei of neurons, to analyze sensory bristle cells of pupal nota of both the null mutant (Fig. 9A-C) and mis-expression (hs-Su(H), data not shown) of Su(H). These experiments show that the neuron/sheath cell lineage is not affected by the loss or increase of Su(H) activity. numb interacts with Su(H) and H in determining hair/socket lineage As described above, numb is involved in all three asymmetric divisions during the formation of a sensory organ in both embryos and adults. The twinned hair phenotype due to loss of function of Su(H) is opposite to the double-socket phenotype due to loss of function of H or numb during the IIa cell division. It therefore raises the possibility that numb, Su(H), and H may determine the IIa cell lineage by acting in the same genetic pathway. Consistent with this, we found that loss of activity of one copy of Su(H) almost completely prevented the doublesocket formation in numb mutants, indicating that Su(H)SF8 behaves as a dominant suppressor of numb mutants. Conversely, we showed that various haplo-insufficient alleles of H act as dominant enhancers of the numbSW allele, since partial reduction of H function greatly increased the double-socket phenotype in flies homozygous for numbSW. In contrast to the suppression of the double-socket phenotype of numbSW/numb2 mutants (Fig. 6; Table 2), we failed to detect any effect on the double-sheath phenotype of the numbSW/numb2 mutants when one copy of the wild-type Su(H) gene was removed, suggesting that this genetic interaction between numb and Su(H) is restricted to the IIa lineage. Su(H) acts downstream of numb in the hair/socket lineage Having found that partially reduced Su(H) activity prevents the double-socket phenotype in the partial loss of numb function mutant, we asked whether Su(H) acts downstream of numb by examining mitotic recombination induced clones that lack the functions of both Su(H) and numb in the SOP lineage. Adult flies carrying clones of null mutations of both numb and Su(H) showed a twinned hair phenotype, resembling the loss of Su(H) function phenotype but opposite to the loss of numb function phenotype (Fig. 8). This result indicates that Su(H) is epistatic to numb in the same genetic pathway that leads to the determination of daughter cell fates of IIa. Given that complete removal of Su(H) functions had no effect on the neuron/sheath cell lineage nor did it alter the double-sheath cell phenotype generated by null mutations of numb (Fig. 9), it appears that Su(H) has no essential role in the neuron/sheath cell specification. How might numb, Su(H), and H regulate one another to specify the hair or the socket cell fate? Our immunostaining results indicate that the Numb protein is asymmetrically localized in the dividing IIa cell (Fig. 3D-F). Therefore, this intrinsic factor Numb is probably preferentially segregated into the future hair cell upon the asymmetric division of the IIa cell. High level of Numb leads to down regulation of Su(H) and the hair fate. Conversely, low level of Numb leads to high level of Su(H) and the socket fate. How does Numb (and possibly H) level affect Su(H) level is unknown. An intriguing possibility is that the regulation is at the level of protein stability as has been shown in the case of Tramtrack in photoreceptor specification (Li et al., 1997; Tang et al., 1997). Numb/Notch signaling mediated by Su(H)-dependent versus Su(H)-independent pathways Notch and numb are involved in an antagonistic manner in all three asymmetric divisions of the SOP lineage during embryogenesis and adult sensory organ formation (Hartenstein and Posakony, 1990; Rhyu et al., 1994; Guo et al., 1996). It has been proposed that Notch is a downstream target gene that is negatively regulated by numb in the determination of daughter cell fate during asymmetric division of the SOP and its daughter cells in the PNS (Guo et al., 1996) as well as the asymmetric division of MP2 in the CNS (Spana et al., 1995; Spana and Doe, 1996) during embryogenesis. Most likely, this epistatic relationship of Notch and numb also applies to adult sensory organ development. Su(H) is known to function in the Notch signaling pathway. Drosophila sense organ development provides a striking example for Su(H) action in only a subset of the binary cell fate decisions controlled by Numb/Notch signaling. Existence of Su(H)-independent Notch signaling is not limited to Drosophila sense organs. Lecourtois and Schweisguth (1995) found that Su(H)-independent Notch signaling is involved in mesectodermal cell fate specification. In a vertebrate muscle 4446 S. Wang and others cell culture system, Shawber et al. (1996) have demonstrated that Notch can inhibit muscle cell differentiation by a mechanism that is independent of CBF1 (a human homolog of Su(H)). An important task for the future is to identify the molecular components used in Su(H)-independent Notch signaling pathways. The Drosophila sense organ should prove to be a useful system for this purpose. We wish to thank C-P. Shen for various GST-fusion proteins of the Numb fragments, M. Guo for the GST-Notch construct, F. Schweisguth for the Su(H) antibody and the fly stock pLacWA1-2-29 Su(H) FRT40A, J. Posakony for the H cDNA. We thank S. Ralls for help with the revertant screen for numbSW isolation. We thank Ming Chan, Ming Guo, Zack Ma, Chaoyang Zeng, Salim Abdelilah, and Manuel Utset for critically reading the manuscript. S. Wang is a postdoctoral research associate, S. Younger-Shepherd is a research associate, and L. Y. Jan and Y. N. Jan are investigators of the Howard Hughes Medical Institute. REFERENCES Ashburner, M. (1989). Drosophila: a laboratory handbook. Cold Spring Harbor: Cold Spring Harbor Laboratory Press. Bailey, A. and Posakony, J. (1995). Suppressor of Hairless directly activates transcription of Enahncer of split complex genes in response to Notch receptor activity. Genes Dev. 9, 2609-2622. Bang, A. G., Hartenstein, V. and Posakony, J. W. (1991). Hairless is required for the development of adult sensory organ precursor cells in Drosophila. Development 111, 89-104. Bang, A. G. and Posakony, J. W. (1992). The Drosophila gene Hairless encodes a novel basic protein that controls alternative cell fates in adult sensory organ developement. Genes Dev. 6, 1752-1769. Bier, E., Ackerman, L., Barbel, S., Jan, L. Y. and Jan, Y. N. (1988). Identification and characterization of a neuron-specific nuclear antigen in Drosophila. 240, 913-916. Blochlinger, K., Bodmer, R., Jan, L. Y. and Jan, Y. N. (1990). Patterns of expression of cut, a protein required for external sensory organ development in wild-type and cut mutant Drosophila embryos. Genes Dev. 4, 1322-1331. Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118, 401-415. de la Concha, A., Dietrich, U., Weigel, D. and Campos-ortega, J. A. (1988). Functional interactions of neurogenic genes of Drosophila melanogaster. Genetics 118, 499-508. Fortini, M. E. and Artavanis-Tsakonas, S. (1994). The Suppressor of Hairless protein participates in Notch receptor signaling. Cell 79, 273-282. Frise, E., Knoblich, J. A., Younger-Shepherd, S., Jan, L. Y. and Jan, Y. N. (1996). The Drosophila Numb protein inhibits signaling of the Notch receptor during cell-cell interaction in sensory organ lineage. Proc. Natl. Acad. Sci. USA 93, 11925-11932. Furukawa, T., Maruyama, S., Kawaichi, M. and Honjo, T. (1992). The Drosophila homolog of the immunoglobulin recombination signal-binding protein regulates peripheral nervous system development. Cell 69, 11911197. Gho, M., Lecourtois, M., Geraud, G., Posakony, J. W. and Schweisguth, F. (1996). Subcellular localization of Suppressor of Hairless in Drosophila sense organ cells during Notch signaling. Development 122, 1673-1682. Guo, M., Jan, L. Y. and Jan, Y. N. (1996). Control of daughter cell fates during asymmetric division: interaction of Numb and Notch. Neuron 17, 27-41. Hartenstein, V. and Posakony, J. W. (1989). Development of adult sensilla on the wing and notum of Drosophila melanogaster. Development 107, 389405. Hartenstein, V. and Posakony, J. W. (1990). A dual function of the Notch gene in Drosophila sensillum development. Dev. Biol. 142, 13-30. Heitzler, P. and Simpson, P. (1991). The choice of cell fate in the epidermis of Drosophila. Cell 64, 1083-1092. Hinz, U., Giebel, B. and Campos-Ortega, J. A. (1994). The basic-helix-loophelix domain of Drosophila lethal of scute protein is sufficient for proneural function and activates neurogenic genes. Cell 76, 77-88. Horvitz, H. R. and Herskowitz, I. (1992). Mechanisms of asymmetric cell division: two Bs or not two Bs, that is the question. Cell 68, 237-255. Lecourtis, M. and Schweisguth, F. (1995). The neurogenic Suppressor of Hairless DNA-binding protein mediates the transcriptional activation of the Enhancer of split complex genes triggered by Notch signaling. Genes Dev. 9, 2598-2608. Lees, A. D. and Waddington, C. H. (1942). The development of the bristles in normal and some mutant types of Drosophila melanogaster. Proc. Roy. Soc. Ser. B 131, 87-110. Li, S., Li, Y., Carthew, R. W. and Lai, Z. C. (1997). Photoreceptor cell differentiation requires regulated proteolysis of the transcriptional repressor Tramtrack. Cell 90, 469-478. Nakao, K. and Campos-Ortega, J. A. (1996). Persistent expression of genes of the Enhancer of split complex suppresses neural development in Drosophila. Neuron 16, 275-286. Parks, A. L. and Muskavitch, M. A. T. (1993). Delta function is required for bristle organ determination and morphogenesis in Drosophila. Dev. Biol. 157, 484-496. Posakony, J. W. (1994). Nature versus nurture: asymmetric cell divisions in Drosophila bristle development. Cell 76, 415-418. Rebay, I., Fleming, R. J., Fehon, R. G., Cherbas, L., Cherbas, P. and Artavanis-Tsakonas, S. (1991). Specific EGF repeats of Notch mediate interactions with Delta and Serrate: implications for Notch as a multifunctional receptor. Cell 67, 687-699. Rhyu, M. S., Jan, L. Y. and Jan, Y. N. (1994). Asymmetric division of Numb protein during division of the sensory organ precursor cell confers distinct fates to daughter cells. Cell 76, 477-491. Rubin, G. M. and Spradling, A. C. (1982). Genetic transformation of Drosophila with transposable element vectors. Science 218, 348-353. Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. Schweisguth, F. (1995). Suppressor of Hairless is required for signal reception during lateral inhibition in the Drosophila pupal notum. Development 121, 1875-1884. Schweisguth, F. and Posakony, J. W. (1992). Suppressor of Hairless, the Drosophila homolog of the mouse recombination signal-binding protein gene, controls sensory organ cell fates. Cell 69, 1199-1212. Schweisguth, F. and Posakony, J. W. (1994). Antagonistic activities of Suppressor of Hairless and Hairless control alternative cell fates in the Drosophila adult epidermis. Development 120, 1433-1441. Shawber, C., Nofziger, D., Hsieh, J. J.-D., Lindsell, C., Bogler., Hayward, D. and Weinmaster, G. (1996). Notch signaling inhibits muscle cell differentiation through a CBF1-independent pathway. Development 122, 3765-3773. Spana, E., Kopczynski, C., Goodman, C. and Doe, C. (1995). Asymmetric localization of Numb autonomously determines sibling neuron identity in the Drosophila CNS. Development 121, 3187-3195. Spana, E. P. and Doe, C. Q. (1996). Numb antagonizes Notch signaling to specify sibling neuron cell fates. Neuron 17, 21-26. Tang, A. H., Neufeld, T. P., Kwan, E. and Rubin, G. M. (1997). PHYL acts to down-regulate TTK88, a transcriptional repressor of neuronal cell fates, by a SINA-dependent mechanism. Cell 90, 459-467. Uemura, R., Shepherd, S., Ackerman, L., Jan, L. Y. and Jan, Y. N. (1989). numb, a gene required in determination of cell fate during sensory organ formation in Drosophila embryos. Cell 58, 349-360. Vaessin, H., Grell, E., Wolff, E., Bier, E., Jan, L. Y. and Jan, Y. N. (1991). prospero is expressed in neuronal precursors and encodes a nuclear protein that is involved in the control of axonal outgrowth in Drosophila. Cell 67, 941-953. Vaessin, H., Vielmetter, J. and Campos-Ortega, J. A. (1985). Genetic interactions in early neurogenesis of Drosophila melanogaster. J. Neurogenet. 2, 291-308. (Accepted 12 September 1997)















![[SENSORY LANGUAGE WRITING TOOL]](http://s1.studyres.com/store/data/014348242_1-6458abd974b03da267bcaa1c7b2177cc-150x150.png)



