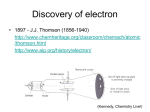
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Gene expression wikipedia , lookup
Point mutation wikipedia , lookup
Ancestral sequence reconstruction wikipedia , lookup
Biochemistry wikipedia , lookup
Paracrine signalling wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Expression vector wikipedia , lookup
Bimolecular fluorescence complementation wikipedia , lookup
Magnesium in biology wikipedia , lookup
Signal transduction wikipedia , lookup
Interactome wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
Protein structure prediction wikipedia , lookup
Protein purification wikipedia , lookup
Western blot wikipedia , lookup
Protein–protein interaction wikipedia , lookup
Magnesium transporter wikipedia , lookup
Proteolysis wikipedia , lookup
Two-hybrid screening wikipedia , lookup
Metalloprotein wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
RIKEN Review No. 35 (May, 2001): Focused on New Trends in Bio-Trace Elements Research Intracellular metal transport proteins Makoto Hiromura and Hiromu Sakurai Department of Analytical and Bioinorganic Chemistry, Kyoto Pharmaceutical University Metal ions are essential cofactors for functional expressions of many proteins in living organisms. In cells, several trace elements are needed to activate and stabilize enzymes, such as superoxide dismutase, metalloproteases, protein kinases, and transcriptional factors containing zinc finger proteins. Recently, intracellular metal trafficking proteins, that carry metal ions to specific target proteins, have been identified in various organisms including mammalians. Such proteins are thus called metallochaperones. This review focuses on the structural and functional characteristics of several metal transport systems, such as those of copper, iron, zinc, manganese, magnesium, and nickel. Abbreviations: Ctr or COPT, copper transporter; Atx1p, antioxidant protein 1; Hah1p, human ATX1 homolog; CopZ or CCH, copper chaperone; CCS, copper chaperone for SOD; Cox17p, cytochrome oxidase 17 protein; CopA, CopB, Ccc2, ATP7A or ATP7B, copper-cation P-type ATPase; RAN1, responsive-to-antagonist 1; PAA1, putative metal transporting P-type ATPase; DMT1, divalent metal transporter 1; Nramp, natural-resistance-associated macrophage protein; Fre, ferrireductase; Fet, multicopper oxidase (Fe transporter); Ftr1p, high-affinity ferric ion transport protein; IRT, iron-regulated transporter; ZRT, zinc-regulated transporter; ZIP, ZRT-, and IRT-like protein; Smf, manganese transporter; PMR, P-type ATPase manganese transporter; Alr or Mgt, magnesium transporter; Nik or Nic, nickel transporter; GST, glutathione S-transferase; M, methionine; C, cysteine; T, threonine; K, lysine; G, glycine; and X, any amino acids. Table 1. Homologous proteins that affect metal ion metabolism in bacteria, yeast, plant, and human. Copper transport proteins The systems consisting of intracellular copper chaperones were identified in bacteria, yeast, plants, and mammals (Table 1).1) As shown in Fig. 1, the copper chaperones consisting of three independent pathways in the cells deliver copper ions to specific target proteins. The first protein is a Atx1p family protein, which targets to copper-cation P-type ATPase in trans-Golgi. This protein has a copper-binding motif, MTCXXC, in the N terminus and a lysine-rich motif, KTGK, in the C terminus. The second protein is a CCS (copper chaperone for SOD) family protein, which has three domains, namely, domain I, domain II, and domain III. Domain I and domain III have metal-binding motifs, namely, MXCXXC and CXC, respectively. Among these three domains, domain III is essential to the transport of copper ions to Cu/Zn SOD. The third protein is a Cox17 family protein, which delivers copper ions to cytochrome c oxidase in mitochondria. Recently, cDNAs of Atx1p and CCS homolog proteins were cloned from rat. 2,3) The rat ATX1 homolog protein named Rah1p was found to have both copper-binding and lysinerich motifs. This protein shows 35% and 89% identities in deduced amino acid sequence with Atx1p and Hah1p (human ATX1 homolog), respectively. Thus, Rah1p was investigated whether it is complementary to a null atx1 mutant strain. The null atx1 mutant strain was defective at the high-affinity Fig. 1. Human metal transport machinery. site for uptake of ferrous ions by Fet3p and thus required an iron-supplemented medium for growth. Moreover, Rah1pexpressing-strains grow in an iron-depleted medium. 2) From these observations, Rah1p was concluded to be an intracel- 23 lular copper transport protein, which delivers copper ions to the copper-cation P-type ATPases. The rat copper chaperone for SOD (rCCS) has a copperbinding motif (MXCXXC) in domain I, a homolog of Cu/Zn SOD in domain II, and an active site (CXC) in domain III. The amino acid sequence of rCCS shows 27, 85, and 94% identities with those of three CCSs, namely, yCCS (yeast CCS), hCCS (human CCS), and mCCS (mouse CCS), respectively. Interestingly, domain I of rCCS shows 33% identity in amino acid sequence with Rah1p, and domain I of yCCS was found to have a very similar structure to that of Atx1p. The amino acid sequence of domain II of rCCS shows 50% identity with that of Cu/Zn SOD. This domain was confirmed to bind with Cu/Zn SOD, using a GST column binding assay. From these experiments, domain II of rCCS was determined to be necessary for the binding of rCCS with Cu/Zn SOD, but both MXCXXC and CXC motifs are not essential for this binding.3) Iron transport proteins Iron transport proteins were identified in various organisms 1 ) (Table 1). DMT1 (divalent metal transporter 1) cloned from rat, is a member of Nramp2 (natural-resistance-associated macrophage protein) family and its cDNA encodes 562 amino acids with 12 putative membrane domains. DMT1, which is a proton-coupled iron transporter protein, mediates the active transport of iron, but incorporates other divalent metal ions, such as Zn2+ , Mn2+ , Co2+ , Cd2+ , Cu2+ , Ni2+ , and Pb2+ , in the proximal duodenum. Ferroportin1 was cloned from zebrafish, mouse, and human. This protein has 570 amino acids with 10 putative membrane domains and its mRNA was found to be expressed at the basolateral surface of the duodenal enterocyte. Thus, ferroportin1 was proposed to be involved in the transport of iron from the basolateral surface to the circulatory system. Saccharomyces cerevisae has a multiple-iron transport protein. Fre1p (Ferrireductase1) identified as a membrane protein, reduced ferric ions to ferrous ions, which were in turn transferred from Fre1p to Fet3p on the cell surface. Fet3p is a multicopper oxidase that displays high functional homology to ceruloplasmin. This protein reoxidizes ferrous ions to ferric ions and transfers them to Ftr1p (high-affinity ferric iron transport protein). Ftr1p is also a membrane protein, and is involved in the uptake of ferric ions in the cell. The iron transport system of the Fet3p and Ftr1p is specific for ferric ions and functions when environmental iron is deficient. IRT (iron-regulated transporter) was identified in Arabidopsis thaliana as a ferrous transport protein. This protein is a cation transporter, which incorporates several metal ions in the cell. Zinc transport proteins The zinc transport protein is a ZIP (ZRT- and IRT-like) family protein (Table 1). ZRT (zinc-regulated transporter) is a zinc transporter in yeast. 4) ZIP family has amino acid sequence homology in yeast, plants, and mammals. The hu- 24 man zinc transport proteins are known as hZIP, hZIP2, and hZIP3. hZIP2 may transport not only zinc ions but also other metal ions in the cell. However, the function of hZIP2, which specifically incorporates zinc ions in the cell, was inhibited by the presence of other metal ions such as Fe2+ , Co2+ , Cd2+ , Cu2+ , and Mn2+ . Moreover, the zinc-uptake activity of hZIP2 is energy-independent. S. cerevisiae has three zinc transporters, namely ZRT1, ZRT2, and ZRT3. When both ZRT1 and ZRT2 act synergetically, it was suggested that zinc uptake by the proteins was regulated by different systems. These three transporters have their own unique characteristics. ZRT1 is a high functional zinc transporter (Km = 10 nM Zn2+ ) and is active in zincdeficient cells. On the other hand, ZRT2 is a low functional zinc transporter (Km = 100 nM Zn2+ ) and is detectable in zinc-replete cells. Finally, ZRT3, which is localized in the vacuolar membrane, was suggested to control a system of zinc influx and efflux transporters in the vacuole. ZIP1, ZIP2, and ZIP3 in plants have different time-, temperature-, and concentration-dependent zinc-uptake activities. On the other hand, ZIP4 was also identified as a ZIP family protein, however, when ZIP4 was expressed, there was no uptake of zinc ions in the cells. ZIP4 has a chloroplasttargeting sequence and therefore, this protein may be expressed in the chloroplast of plant cells. Other metal transport proteins (Table 1) Manganese Smf1 and Smf2, identified as manganese transport proteins in S. cerevisiae, were proposed to function as high- and low-affinity transporters for manganese uptake, respectively. PMR1, which is a member of the P-type ATPase family and is localized in the Golgi apparatus, accumulates elevated levels of intracellular manganese ions. This protein is involved in the homeostasis of manganese ions in yeast cells. Nramp1 and Nramp2 in mammals have high homologies in amino acid sequence to Smf1 and Smf2, respectively. Nramp2 was found to complement the function of the Smf yeast mutant, in contrast, Nramp1 could not complement this function. Thus, Nramp2 was suggested to be involved in the uptake of manganese ions in the cell. Magnesium Magnesium transport proteins were characterized in Gramnegative bacteria.5) Two magnesium transporters, MgtA and MgtB, are members of the P-type ATPases that are localized in the periplasmic membrane and mediate magnesium ion influx. These transporters were found to be related to the roles of calcium ATPases in eukaryotes. PhoP and PhoQ are also magnesium-regulated proteins in bacteria. PhoQ, which functions as a membrane sensor kinase of magnesium ions, is present as an inactive form when bound with magnesium ions. However, when magnesium ions dissociate from PhoQ, this protein is activated, which in turn phosphorylates PhoP. Therefore, PhoQ is called a magnesium-dependent transcriptional factor. Nickel Nickel transport proteins activate some nickel-dependent en- zymes such as urease, hydrogenase, and carbon monoxide dehydrogenase.6) Highly specific nickel transporters were identified as five genes in the Nik ABCDE operon in Escherichia coli under anaerobic conditions. Nik A is a nickel-binding protein, which is localized in the periplasmic membrane. Nik B and NikC are integral membrane subunits, and NikD and NikE are nucleotide-binding proteins. NixA was cloned as a nickel permease in Helicobacter pylori. The expression of this protein in E. coli allowed nickel transport and increased urease activity. UreE, the urease accessory protein in Klebsiella aerogenes, specifically delivers nickel ions to the urease apoprotein, then the urease is activated. Therefore, UreE is a nickel chaperone protein, which targets urease. Nic1p was identified as the firstly reported eukaryotic nickel permease as well as a homolog protein of the bacterial nickel transporter in Schizosaccharomyces pombe. This protein has four characteristic amino acids that are conserved in the bacteria nickel transporter. Two motifs are located within the transmembrane segment and other motifs are shown to be essential for transport activity. Nic1p is a high-affinity nickel transporter and its function is inhibited by cobalt ions. Conclusions Several metal ions are essential for all living organisms, and some metal ions activate many enzymes and maintain the homeostasis of the cells. In this review, the recently reported metal transport proteins are summarized. Metal transport proteins exist widely in both prokaryotes and eukaryotes and many transport proteins are identified. Metal transport proteins recognize specific metal ions. Membrane-type transporters carry metal ions from outside of the cell to inside, then the intracellular metal chaperones transport the metal ions to the specific targeted proteins. Recently, it has been reported that some diseases are caused by abnormal metabolisms of certain metal ions in cells and tissues. For example, Menkes and Wilson’s diseases, that are hereditary disorders of copper metabolism, were found to be caused by mutated ATP7A and ATP7B genes that encode copper-cation P-type ATPases. Excess iron in cells is related to the pathologies of hereditary hemochromatosis, Parkinson’s disease, and the neurological disease Friedreich ataxia. When manganese ions are present at abnormal levels in cells, pediatric neurological disorders develop. On the basis of the numerous observations reported, metal transport proteins are important in protecting living organisms from metabolic disorders that in turn develop into diseases and in maintaining the metal ion homeostasis in them. References 1) D. Radisky and J. Kaplan: J. Biol. Chem. 274, 4481 (1999). 2) M. Hiromura and H. Sakurai: Biochem. Biophys. Res. Commun. 265, 509 (1999). 3) M. Hiromura et al.: Biochem. Biophys. Res. Commun. 275, 394 (2000). 4) M.-L. Gueriot: Biochim. Biophys. Acta 1465, 190 (2000). 5) M. Beth et al.: J. Biol. Inorg. Chem. 4, 523 (1999). 6) R.-P. Hausinger: J. Biol. Inorg. Chem. 2, 279 (1997). 25