Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Tissue engineering wikipedia , lookup
Hedgehog signaling pathway wikipedia , lookup
Purinergic signalling wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cell culture wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
List of types of proteins wikipedia , lookup
Cellular differentiation wikipedia , lookup
Signal transduction wikipedia , lookup
Biochemical cascade wikipedia , lookup
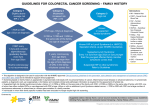
Molecular mechanisms involved in colorectal cancer initiation and progression Colorectal cancer (CRC) is the third most common type of cancer and the second cause of death by cancer in the Western world. Elena Sancho CRC results in around 650,000 deaths worldwide per year. Most sporadic CRCs arise from adenomas that initially are benign and occur frequently: approximately 50% of the Western population develops an adenoma by the age of 70. However, the development of a full-blown malignant colorectal tumour is a progressive process that often takes several years. During this period, the progression of the disease appears to follow a precise series of molecular events, requiring the accumulation of mutations in proto-oncogenes and tumour suppressor genes in these initially benign lesions. Access to specimens of CRC at different stages of the malignancy has allowed the analysis of the molecular alterations most frequently associated with each step of the disease (reviewed in Sancho et al, 2004). The aim of the research performed in our laboratory is to decipher the molecular instructions that underlie the signalling pathways that are altered in CRC and that are responsible for the initiation and progression of the disease. WNT signalling and the initiation of CRC Around 70% of sporadic colorectal tumours show biallelic inactivation of the APC gene (Adenomatous polyposis Coli). A high percentage of remaining tumours show activating mutations in beta-catenin or axin. These molecules are components of the Wnt signalling pathway. Activating mutations of the Wntsignalling pathway are the only known genetic alterations present in early premalignant lesions in the intestine, such as aberrant crypt foci and small adenomas. In various animal models, activating mutations in this pathway effectively initiate tumorogenesis in the intestine in a process characterised by the formation of dysplastic crypts and adenomas similar to those found in humans. Therefore, it is widely accepted that constitutive activation of Wnt signalling caused by mutations in components of the pathway Figure 1. Genetic alterations frequently associated with CRC progression. 118 2007 S cie ntif ic Re po r t O nco lo gy Pro gramme is responsible for the initiation of CRC (reviewed in Sancho et al, 2004; Figure 1). Mutations in Wnt signalling components that lead to CRC result in the stabilization and accumulation of beta-catenin in the nucleus, and, as a result, in increased transcriptional activation mediated by the beta-catenin/TCF complex. Therefore, the transactivation of beta-catenin/TCF target genes represents a primary transforming event in CRC. A few years ago we identified the genetic programme driven by betacatenin and TCF in CRC cells. Our studies indicated that beta-catenin/TCF target genes are expressed not only in tumours but also in normal non-transformed intestinal progenitors cells at the bottom of crypts (van de Wetering et al, 2002; see Figures 2 and 3). Our results, together with those obtained from several animal models in which Wnt signalling had been genetically manipulated (Pinto et al, 2003; Korineck et al, 1998), implied that the stem cell and progenitor compartments were controlled by Wnt signalling. These findings led us to propose that in CRC the first step towards malignancy consists of the acquisition of a crypt progenitor-like phenotype (van de Wetering et al, 2002). Our hypothesis has marked a milestone in the field and has totally changed views on the initiation of this disease. We are currently developing animal models that will formally confirm this notion and help to shed light on why Wnt signalling mutations are an important pre-requisite for the development of CRC. Our studies are oriented towards the identification of the nature of the founding CRC cell and the mechanisms by which it escapes cell renewal. During 2007, we have generated several DNA constructs to enable the development of animal models that will be used for this purpose. These include the conditional expression in the intestine of oncogenes involved in CRC combined with colour markers that will help to identify mutant cells. These studies may shed additional light on specific pathways that could be targeted to block the progression of this disease. Likewise, in collaboration with Eduard Batlle’s laboratory (IRB Barcelona), and by systematic analysis of colorectal samples at different stages of the disease, we aim to identify the core set of instructions imposed by Wnt signalling mutations that remains unaltered throughout the carcinogenetic process. This research will provide crucial information on the molecular targets for CRC at all stages of the malignancy. Having identified that the initial event triggering transformation is the blockage of founder tumour cells into a progenitor phenotype, our laboratory now seeks to identify differences between the true physiological progenitors and initial founder mutant cells. To this end, during 2007, we have developed a protocol which allows us to isolate epithelial cells from the bottom of colonic crypts (ie, stem cells and early progenitors) from fresh tissue. This protocol can also be used to isolate tumour cells from early adenomas or dysplastic crypts. We are currently comparing the genetic profile of physiological progenitors with that of tumour cells from adenomas. We aim to identify tumour-specific molecular targets susceptible for targeting by the pharmaceutical industry. These targets will be particularly useful for patients suffering Familial Adenomatous Polyposis (FAP). These patients inherit a mutation in the APC gene, and due to loss of heterozygosity (LOH) they develop hundreds of polyps in the intestinal tract and are therefore predisposed to the development of malignant CRC. TGF-beta signalling during CRC progression In recent years, some of the leading scientific teams, including ours, have reported that the onset and progression of CRC can be understood using several concepts taken from the Darwinian evolution model. Under this view, the sequence of mutations acquired during CRC progression (Figure 1) can be explained by colorectal tumours evolving through a series of bottlenecks or restriction points at which only those cell cells acquiring the correct mutational event expand and progress to the next stage of malignancy. Our research interest focuses on determining how the Research Group Members I Principal Investigator: Elena Sancho I Postdoctoral Fellows: Alexandre Calon, Annie Rodolosse I PhD Students: Elisa Espinet I Research Assistant: Sergio Palomo I Lab Technician: Mireia Humà acquisition of mutations in other signalling pathways may modulate the initial progenitor phenotype imposed by Wnt signalling to overcome the bottlenecks associated with CRC progression. One of the most prevalent types of mutation during CRC progression are those that inactivate the TGF-beta signalling pathway (reviewed in Grady and Markowitz, 2003; Figure 1). This pathway is involved in numerous processes in the development and homeostasis of adult tissues. TGF-beta ligands activate the signalling pathway by binding to TGF-beta receptor type II homodimers. Ligand-bound receptor II recruits TGF-beta receptor I homodimers, which are Onc ology Programme 2 0 0 7 S c i e n t i f i c Report 11 9 subsequently transphoshorylated and thus activated by receptor type II. Phosphorylation of the intracellular mediators smads by activated receptor I allows dimer formation with smad-4 and translocation to the nucleus, where the specific outcome of the signalling will depend on the cell type and the context of the cell itself (reviewed in Shi and Massagué, 2003). Mutations found in CRC affect mainly the TGF-beta receptor type II and the intracellular smads, smad-2 and smad-4, by abolishing the transcriptional effects mediated by TGF-beta. Figure 2. Expression pattern of EphB2 (a,b) and Carbonic Anhydrase II (c,d) in early tumour lesions. EphB2, an example of a target gene under the control of beta-catenin/TCF, is expressed by tumour cells from dysplastic crypts (a, black arrows), but also by normal progenitor cells at the bottom of intestinal crypts (b, black arrows). In a complementary fashion, the differentiated marker carbonic anhydrase II is expressed only in the top half of the crypts and surface epithelium (c, d black arrows), and is absent from tumour lesions (c, white arrows). This expression pattern is observed for the beta catenin/ Tcf target genetic programme identified so far. Figure 3. Depiction of a colon crypt and proposed model for CRC initiation. The intestinal epithelium is organised in invaginations called crypts of Lieberkühn. Epithelial cells within these crypts are in constant renewal thanks to a small group of stem cells that reside at the bottom of the crypts. Stem cells divide asymmetrically to give rise to early progenitors, which rapidly divide whilst migrating towards the intestinal lumen. As they do so, they become predetermined towards differentiation into one of the functional cell types present in the intestine (adsorptive, mucosecreting or enteroendocrine cells). The proliferative compartment of intestinal crypts is maintained by the target gene programme directed by beta-catenin/Tcf in response to wnt signals. When these cells physiologically down-modulate beta-catenin/Tcf activity, they cease proliferation and differentiate. Cells mutant in components of wnt signalling become independent of these signals, exhibit constitutive activation by betacatenin/Tcf, and are blocked in a progenitor proliferative phenotype. 120 2007 S cie ntif ic Re po r t O nco lo gy Pro gramme Our lab currently focuses on the role of TGF-beta signalling in CRC progression. We are studying the transcriptional events controlled by TGF-beta in CRC cells. We have already identified changes in approximately 500 genes in response to TGF-beta in these cells. Unsupervised analysis of a collection of tumours of known transcriptomes on the basis of the TGF-beta signature obtained in our laboratory perfectly discriminates adenomas from carcinomas, thereby implying that these genes contain the information that drives the adenoma/carcinoma transition. We are now dissecting this information in order to identify TGF-beta genes that play an executive role in the adenoma/carcinoma transition. Our studies in 2007 pinpoint a relevant role for TGF-beta signalling in the modification of several cellular responses required for tumour progression. Our results reveal a gain-of-function in TGF-beta signalling during CRC progression. This increase appears to translate into the expression of a series of TGF-beta target genes in several cell types within the tumour, including mesenchymal cells (see Figure 5). We have developed orthotropic models of colorectal tumours in nude mice to test the role of the TGF-beta-controlled gene signature. We are currently performing systematic shRNA-mediated down-regulation of genes contained in this signature in order to screen for TGF-beta-regulated genes that are crucial for CRC. In addition, we are studying whether the TGF-beta signature in CRC has a predictive value for clinical outcome. Publications Cortina C, Palomo S, Iglesias M, Fernandez-Masip JL, Vivancos A, Whissell G, Huma M, Peiro N, Gallego L, Jonkheer S, Davy A, Lloreta J, Sancho E and Batlle E. EphB ephrin-B interactions suppress colorectal cancer progression by compartmentalising tumour cells. Nat Genet, 39, 1376-83 (2007) Other References Batlle E, Henderson JT, Beghtel H, van den Born MM, Sancho E, Huls G, Meeldijk J, Robertson J, van de MW, Pawson T and Clevers H. Beta-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell, 111, 251-63 (2002) Batlle E, Bacani J, Begthel H, Jonkeer S, Gregorieff A, van de Born M, Malats N, Sancho E, Boon E, Pawson T, Gallinger S, Pals S and Clevers H. EphB receptor activity suppresses colorectal cancer progression. Nature, 435, 1126-30 (2005) Grady WM and Markowitz SD. Genetic and epigenetic alterations in colon cancer. Annu Rev Genomics Hum Genet, 3, 101-28 (2002) Korinek V, Barker N, Moerer P, van Donselaar E, Huls G, Peters PJ and Clevers H. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nature Genet, 19, 379-83 (1998) Pinto D, Gregorieff A, Begthel H and Clevers H. Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes Dev, 17, 1709-13 (2003) Sancho E, Batlle E and Clevers H. Signalling pathways in intestinal development and cancer. Annual Review of Cell and Dev Biol, 20, 695-723 (2004) Sancho E, Batlle E and Clevers H. Live and Let die in the intestinal epithelium. Curr Opin Cell Biol, 15, 763-70 (2003) Shi Y and Massagué J. Mechanisms of TGF-beta signalling from cell membrane to the nucleus. Cell, 113, 685-700 (2003) Figure 4. The TGF-beta signature discriminates between adenomas and carcinomas. Unsupervised clustering analysis of a collection of tumours of known transcriptomes on the basis of target genes controlled by TGF-beta signalling clearly classifies adenomas and carcinomas in two separate branches. van de Wetering M, Sancho E, Verweij C, de Lau W, Oving I, Hurlstone A, van der HK, Batlle E, Coudreuse D, Haramis AP, Tjon-Pon-Fong M, Moerer P, van den BM, Soete G, Pals S, Eilers M, Medema R and Clevers H. The beta-catenin/TCF-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell, 111, 241-50 (2002) Research Networks and Grants a b Molecular mechanisms involved in colorectal cancer initiation Spanish Ministry for Education and Science, SAF2005-002170: 2006-2008 Research Director: Elena Sancho Start-up grant for emergent research groups Generalitat de Catalunya, Agència de Gestió d’Ajuts Universitaris i de Recerca, 2005SGR 00775: 2006-2009 Research Directors: Eduard Batlle and Elena Sancho Variations in the genetic program under the control of betacatenin/Tcf during colorectal cancer progression Fundació La Caixa, BM06-241-0: 2007-2009 Research Directors: Eduard Batlle and Elena Sancho Collaborations TGF-beta target genes in CRC Giancarlo Marra, Institute of Molecular Cancer Research (Zurich, Switzerland) Wnt signalling in CRC Hans Clevers, Hubrecht Laboratory (Utrecht, The Netherlands) Figure 5. Representative example of the expression pattern of TGF-beta target genes in colorectal adenomas (a) and carcinomas (b). Onc ology Programme 2 0 0 7 S c i e n t i f i c Report 12 1