Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix wikipedia , lookup
Cytoplasmic streaming wikipedia , lookup
Endomembrane system wikipedia , lookup
Chromatophore wikipedia , lookup
Tissue engineering wikipedia , lookup
Cell growth wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell culture wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cytokinesis wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
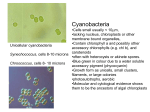
1 Cyanobacteria (Oxygenic Phototrophs) The cyanobacteria (the earlier blue-green algae), or the blue-green bacteria, represent a group of photosynthetic, mostly photolysis-mediated oxygen-evolving monerans (prokaryotes). These are the only organisms able to perform oxygenic photosynthesis that can also fix nitrogen. These organisms are amongst the oldest organisms known dating back to the early Precambrian period 3.6 x 109 years ago and probably played a crucial role in the evolution of higher plants Cyanobacterial thallus ranges from unicellular, colonial to filamentous; multiseriate branched filamentous condition is the highest level of organization attained by them. Because of their close architectural, physiological and biochemical similarities with bacteria, especially the gram-negative ones, the cyanobacteria have been placed under a separate division, namely, 'The Cyanobacteria' in Bergey's Manual of Determinative Bacteriology (8th edition, 1974), which has been accepted and used widely as the standard reference for bacterial toxonomy. This manual, however, has recognized the Kingdom-Monera of Whittaker, but has called it Kingdom-Prokaryotae because of the prokaryotic nature of all the monerans. The cyanobacteria possess various distinguishing characters, which can be summarized as under. (a) Cyanobacteria can grow in diverse habitats, but one striking feature in their occurrence and predominance in habitats alternating between photoaerobic and photoanaerobic conditions can be correlated with their preference for low oxygen tension and low redox-potential. These properties stem from their recently discovered dual-capacity of oxygenic photosynthesis and facultative anoxygenic photosynthesis. (b) The cyanobacteria possess various morphologically distinctive structures, e.g., akinetes and heterocysts. (c) The main cell wall constituent of cyanobacteria is peptidoglycan. (d) The cyanobacterial cytoplasm is traversed extensively by flattened vesicular structures called thylakoids or lamellae, the photosynthetic sites. (e) The principal photosynthetic pigment of all cyanobacteria is chlorophyll a. Besides, they possess bcarotene and other accessory pigments, namely, phycobiliproteins. The phycobiliproteins are phycocyanin (PC), allophycocyanin (AP), allophycocyanin B (APB), and phycoerythrin. (f) Most filamentous cyanobacteria show a gliding motility at some stage of development; they lack flagella. Classification Rippka et al. (1979) have proposed a modern scheme of cyanobacteria classification taking mainly their physiology, cell constituents and DNA characteristics into consideration. They have created five sub-groups called 'sections'. 2 System of classification of cyanobacteria Section Cell type I Unicells or aggregates II Unicells or aggregates III IV V Filaments; unbranched trichomes with only vegetative cells Filaments can form heterocysts; no true branching Filaments can form heterocysts and true branches Mode of reproduction Range Recognized Genera of G + C% Binary fission or 35-71 Synechocoous, budding Synechocyatis, Gloeobacter, Gloeotheca, Gloeocapsa, Chamesiphon Multiple fission to 38-47 Dermocarpa, form baeocytes Xenococcus, Myxosarchina, Binary fission in a 40-67 single plane Hormogonia form; 38-47 binary fission in a single plane Hormogonia, Akinetes, 42-46 Hormocysts; binary fission in more than one plane. 42-46 Chroococcidiopsis Spirulina, Lyngbya, Oscillatoria, Pseudoanabaena, Phormidium, Plectonema Anabaena, Nodularia, Nostoic, Cylindrospermum Scytonema, Calothrix, Tolypothrix, Rivularia Chogloeopsis, Fischerella, mastigocladus Other properties Almost always nonmotile Usually some baeocytes are motile Usually motile Often motile; may form akinetes Greatest morphological complexity and differentiation in cyanobacteria Ultrastructure of Cyanobacterial Cell (i) Mucilage Sheath The cells and filaments of most cyanobacteria are generally surrounded by a mucilaginous sheath whose thickness, pigmentation, consistency, and nature is greatly influenced by environmental factors. It is considered that these microorganisms secrete the mucilage through pores present in their cell walls (iii) Plasma membrane A typical cyanobacterial plasma membrane is 70A thick. It is selectively permeable, lacks sterole such as cholesterol, and consists of a high proportion of protein to phospholipid (typically 2: 1) like other monerans. It generally fuses with the photosynthetic lamellae and gives rise to inward foldings in the cytoplasm; the foldings are called lamellosomes or mesosomes (Fig. 4.10). The latter are mostly similar in functions to mesosomes occurring in other monerans. 3 Anaeba Possessing Heterocyst and Akinete 7.Cell wall 1. Photosynthetic Lamella 8. Cell Membrane 2. Ribosome 9. Cyanophycin Granule 3. Beta-Granule 10. DNA 4. Alpha-Granule 11. Protein Body 5. Polyphosphate Granule 12. Reserve Food 6. Sheath 13. Lamellosome The Cytoplasm The cytoplasm of cyanobacterial cell, like that of bacteria, is incredibly boring. It lacks eukaryotic organelles such as chloroplasts, mitochondria, endoplasmic reticulum, Golgi bodies. But it possesses photosynthetic apparatus, ribosomes, and a large number of subcellular inclusions such as glycogen or a-granules, polyphosphate bodies, polyhedral bodies, cyanophycin granules, and the genetic material Photosynthetic apparatus. In place of the chloroplasts of photosynthetic eukaryotes, cyanobacteria have flattened vesicular structures called thyllakoids or lamellae, which resembles the individual thyllakoids of the true chloroplasts of photosynthetic eukaryotes. The lamellae or thyllakoids are both physiologically or structurally complex and possess photosynthetic pigments. As described earlier, the principal pigment of all cyanobacteria is chlorophyll a. In addition, there are b-carotene and other accessory pigments, namely, phycobiliproteins. The phycobiliproteins are phycocyanin (PC), allophycocyanin (AP), allophycocyanin-B (APB), and phycoerythrin. By possessing phycocyanin and phycoerythrin accessory pigments, the cyanobacteria resemble with red-algae. However, the necessary pigments of these organisms are generally organized into organelles called phycobilisomes and trap light energy of lower wavelengths, which cannot be absorbed by chlorophyll a, and pass it on to the chlorophyll in the diagram. This is the reason why cyanobacteria, like green algae, can exploit deeper waters where the quality and quantity of illumination is inappropriate for the photosynthetic plants. 4 Energy Shunting of Phycoerythrin and Phycocyanin (b) Ribosomes. These are the sites of protein synthesis. Cyanobacterian ribosomes occur freely In the cytoplasm and are identical to those of bacteria in being 70S ribosomes. (c) Glycogen or alpha-granules. Glycogen or alpha-granules are the sites for storage of excess photosynthetic products. The latter is used as energy source in darkness or when CO2 supply is limiting. (d) Polyphosphate bodies. These are the spherical structures formed as a result of the aggregation of high molecular weight linear polyphosphates. These subcellular inclusions are also called metachromatin granules or volutin granules and serve as phosphate stores and are consumed during periods of phosphate starvation. These structures develop mostly in those cyanobacteria, which grow in a phosphate-rich environment. (e) Polyhedral bodies. All cyanobacteria store their ribulose I, 5-bisphosphate carboxylase (RUBP carboxylase) enzyme in structures referred to as polyhedral bodies. (f) Cyanophycin granules. Cyanobacteria growing in nitrogen-rich environment produce structures called cyanophycin granules, which are made up of arginine and aspartic acid. (g) Genetic material. The genetic material of cyanobacteria is made up of naked DNA fibrils found dispersed in the central region of the cytoplasms. Like other monerans, they lack membrane-¬bound organized nucleus. The exact number of genomes per cell is not yet known; it has recently been reported that Agmenellum contains 2, 3 or more copies of its genetic material. The molecular weight of the cyanobacterial genome is considered to range from 2.7 to 7.5 x 109 daltons. (h) Plasmids. All the naturally occurring plasmids in cyanobacteria are phenotypically cryptic. They are covalently closed circular DNAs and their genetic compositions and complete function is not yet known. However, plasmid-mediated transfer of genetic material has been reported in certain cyanobacteria Reproduction Like bacteria, the cyanobacteria also reproduce asexually and the commonest mode of reproduction in them in transverse binary fission (see bacteria). In addition, there are certain specialized structures such as akinetes, hormogonia, hormocysts and spores, which are partly involved in the process of reproduction. So far as the sexual reproduction in its true sense is concerned, it is absent in them and the requirements of sexuality are considered to be met by some alternative pathways referred to as parasexual-pathways 5 (i) Akinetes Most filamentous cyanobacteria develop perennating (dormant structures) in adverse condition. These structures are larger than the vegetative cells, are equipped with thick walls, and are called akinetes (Fig. 4.12). When favourable conditions return, they germinate and produce new filaments. (ii) Hormogonia All filamentous Cyanobacterium reproduce by fragmentation of their filaments (trichomes) at more or less regular intervals to form short pieces each consisting of 5-15 cells. These short pieces of filaments are called hormogonia. The latter show gliding motility and develop into new full-fledged filaments. (iii) Hormocysts Some cyanobacterial produce hormocysts, which are multicellular structures having a thick and massive sheath. They may be intercalary or terminal in position and may germinate from either end or both the ends to give rise to the new filaments. (iv) Spores Nonfilamentous cyanobacteria generally produce spores such as endospores, exospores and nanocysts which contribute by germinating and giving rise to new vegetative cells when the unfavourable condition is over. Endospores are produced endogenously like those in bacteria; exospores are the result to exogenous budding of cells, and the nanocysts are produced endogenously like endospores. The difference between an endospore and a nanocysts is that in endospore formation the parent cell concomitantly enlarges in size, whereas in nanocysts formation there is no such enlargement of the cell. Parasexuality in cyanobacteria The knowledge of cyanobacterial genetics is relatively new and was pioneered by Kumar (1962) who obtained penicillin and streptomycin resistant strains of Anacystis nidulans, crossed them, and successfully demonstrated the appearance of a third type of recombinant strain resistant to both the antibiotics. However, the mechanisms of genetic recombination in cyanobacteria are thought to be the same as those of bacteria The existence of the process of transformation in cyanobacteria was established experimentally in 1979 by Doolottle. Stevens and Porter in 1980 has successfully demonstrated this process in Agmenellum quadruplicatum. The transforming principle was shown to be DNA. Some cyanobacteriologists, however, have found that the transformation is mediated in some cases by complexes of DNA and RNA. The first report on conjugation in a Cyanobacterium, namely, Anacysts nidulans was by Kumar and Ueda (1984). The frequency of conjugation is very low (about 1 in 106 cells) and cells conjugate by means of a conjungation tube. The knowledge of transduction in these microorganisms at present is restricted to some preliminary reports. However, the occurrence of different cyanophages, e.g., LIP 1-7, SM-1, N-1 and infection of several cyanobacteria by them prompts one to imagine that the virus-mediated method of genetic transfer (transduction) in cyanobacteria will be conclusively established in the near future Heterocyst in Cyanobacteria As stated earlier, the cyanobacteria are the only organisms able to perform oxygenic photosynthesis 6 that can also fix nitrogen; many, but not all, are vigorous nitrogen fixers. The coexistence of the processes of oxygenic photosynthesis and intrinsically anaerobic nitrogen fixation process in a single organism presents an obvious paradox because nitrogenase, the key enzymes, is rapidly and irreversibly inactivated by an exposure even to low partial pressure of oxygen. However, the nitrogen fixing cyanobacteria produce a specialized type of cell, the heterocyst, within which nitrogen is fixed Filamentous forms of cyanobacteria such as Anabaena form large, distinctive cyst-like cells, the heterocysts at intervals along the trichome (filament), . The latter develop from normal vegetative cells particularly in conditions deficient in NH3+ or NO3-, and are considered to be the site of nitrogen fixation. The conversion of atmospheric nitrogen to ammonia takes place under highly anaerobic conditions, that only the heterocysts are able to provide. For instance, the oxygen-evolving part of the photosynthetic mechanism (photosystem II) is blocked in heterocysts, and the remaining photosynthetic machinery becomes geared to provide energy for the reduction of nitrogen to NH3+. Trichome of Anabaena possessing Heterocyst and Akinetic 1. Vegetative Cells 2. Heterocyst 3. Akinete 4. Thick wall 5. Cytoplasm Devoid of Photosystem II 6. Cyanophycin Plug 7. Thicker Mucilaginous Coat Heterocyst: 1. Heterocysts are distinctive and specialized cells which occur in certain filamentous genera of blue-green algae. 2. These are round , large in size and possess a thick wall and one or two polar nodules on the side of attachment with the negative cell or cells. 3. Mucilage envelops except at the region of polarity. 7 4. 5. 6. 7. The cell contents are homogenous and yellowish green in appearance. They occur singly in terminal or intercalary position in a filament. Newly formed vegetative cell gets transformed into a heterocyst. Vegetative enlarges and cell wall become many layered . Except carotenoides other pigments are lost. 8. granular bodies disappear , the photosynthetic lamella become and form a complex reticulation . 9. Under certain environmental conditions , the protoplast of the heterocyst may rejuvenate and germinate. 10. Heterocyst function differently under different environmental conditions. They reported to play role in sporulation. 11. Heterocyst can transform in to akinetes . 12. They may provide a suitable place for fragmentation of the filaments. 13. It is also reported to be a store house of enzymes and they play role in nitrogen fixation. Akinetes ( Resting Spores ): 1.These Are oblong or spherical and large in size than vegetative cells. 2. They do not contain photosynthetic pigment. 3. Contain abundant food granules. 4.They have thick wall ; the outer wall shows ornamentation and yellow or brown in colour. 5. Thickness of wall is due to deposition of material during enlargement . 6.Akinetes may occur singly or in long chains. 7. They can with stand desiccation and extremes of temperatures. 8.They germinate after a period of rest. Nutrition: Blue-green algae are obligatory autotrophic . A few members of Nostoc can grow heterotrophically. Similarities between blue-green algae and Bacteria. 1. The nucleus in both lack double membrane. DNA fibrils are dispersed in the central region of the cell . Histones are absent . 2. Blue-green algae and photosynthetic bacteria possess thylakoids which are not membrane bound. 3. True sexuality is absent .No meiosis 4. Cytoplasmic streaming is absent .Cell organelles such as endoplasmic reticulum , mitochondria, golgi bodies and membrane bound vacuoles are absent. 5. photosynthetic lamellae or thylakoides perform photosynthesis , respiration , oxidative phosphorylation and nitrogen fixation. 6. Cell wall possess a characteristic muco eptide , a l- aminopimelic acid and muramic acid component. 7. Both have a highly refined and complex biochemical property of fixation of elementary nitrogen and oxidation of H2S . 8. 9. Morphological resemblance are noticed between Beggiatoa and Oscillatoria in structure, method of movement and deposition of sulphur droplets in the cells. 10. Hormogonia like structure occur in thiothrix , Sheath and false branches are known to be present in the filaments of Cladothrix. 11. Both are susceptible to phages ;cyanophages and bacteriophages respectively. 8 12. Fossile evidence show that the unicellular forms of the two groups were first cellular organism to have evolved on the surface of earth some 3 billion years before. Cyanobacteria Promient Characters 1.Possess two photosystems; PS-I and PS-II 2.Photosynthetic pigments are Chlorophyll-a Phycobiliproteins and carotenoides. 3.Electron source is water . Reduced compounds for few species. 4.Respiration is aerobic when growing photosynthetically. 5. Some strains can fix Nitrogen in the dark. Differences between Blue-green algae and Bacteria Blue-green Algae 1) Majority of them are photosynthetic. Bacteria Majority of them are saprophytic , parasites or chemo synthetic. 2) Flagella are absent. Flagella are present. 3) Endogenous spore formation is absent. Present 4) Chlorophyll-a is main photosynthetic Bacteriochlorophyll is main photosynthetic pigment. pigment. 5) Hydroxyl ions are used as hydrogen donor Hydroxyl ions are not used and oxygen is not and oxygen is evolved. evolved. 6) Contain certain unsaturated fatty acids. Do not contain. 7) Heterocyst present. Heterocyst absent. Classification: The most commonly followed classification is the one proposed by Fristch ( 1945). a) Without hormogonia Order-I Chrococcales. 1) Unicellular or colonial 2) Multiplication by cell division and by endospores Ex: Chrococcus, Gleocapsa Order-II Chamaesiphonales 1) Unicellular or colonial epiphytes or lithophytes exhibiting marked polarity. 2) Multiplication by endospores or exospores. Ex: Dermocapsa , Chamaesiphon. Order-III Pleurocapsales 1)Heterotrichous filamentous types 2) Heterocyst absent 3) Multiplication by endospores , hormogonia absent. Ex: Pleurocapsa, Hyella. With hormogonia. Order-IV Nostacales 9 1) Non –heterotrichous filamentous type showing false branching. 2) Herocyst present 3) Multiplication by hormogonia, hormocysts and akinetes. Ex: Nostoc , Oscillatoria Order-V Stigonematales. 1) Heterotrichous filamentous type with true branching . 2) Herocyst present 3) Ex: Stigonema, Haplosiphon. Important Characteristics of Blue-Green Algae Order General Shape Reproduction & Growth Heterocyst Other properties Rep.Genera Chroococcales Unicellular rods or cocci:nonfilamentous aggregates Binary fission,budding Absent Almost always nonmotile Gleocapsa Chamaesiphon Pleurocapsales Unicelluar rods or cocci;may be held together in aggregates Multiple fission to form baeocytes Absent Only some baeocytes are motile Plerocapsa Dermocarpa oscillatoriales Filamentous, unbranched trichome with only vegetative cells Binary fission in a single plane,fragmentation Absent Usually motile Oscillatoria Spirulina Nostacales Filamentous , unbranched trichome may contain specialised cells Binary fission in a single plane , fragmentation to form hormogonia Present Often motile , may produce akinetes Filamentous trichomes either with branches or composed of more than one row of cells Binary fission in more than one plane , hormogonia formed Present Stigonematales May produce akinetes ,greatest morphological complexity in cyanobacteria Nostoc Anabaena Stigonema Fishcherella 10 CYANOPHYCEAE HORMOGONALES (NOSTOCALES) SCYTONEMATACEAE SCYTONEMA Occurrence: Generally found as intricately fused filamentous masses either in purely aquatic or semi terrestrial conditions. Species of Scytonema like S. simplex are completely submerged forming the undergrowths of ponds and. lakes. Some species are found attached to aquatic plants. Terrestrial species like S. oscillatum form mats on wet rocks, damps barks of trees etc. Some species of Scytonema are also known to occur as "Phycobionts" (algal partners! in Lichens. Structure: The plant body is a filament and is surrounded by mucilage sheath, like in other blue green algae. The. sheath is lamellated and of golden yellow or brown color. 'Ihe filaments possess either intercalary or terminal heterocysts. The genus is characterized by its unique type of branching. This is known as ,"false branching" or "Gemminate branching". The filament breaks up near a heterocyst and the broken end protrudes out of the sheath as a branch. If both the free ends protrude out as branches there will be two branches and it is known-as "gemminate branching". If only one develops into a branch, it is known as "false branching". Later the branches develop their own mucilage sheath. Cause of Branching: Branching is the result of the following causes: (1) By the degeneration of Intercalary cell: When one intercalary cell dies, the free ends of cells continue growth resulting in gemminate branching. (2) By the development of separation discs: An intercalary cell in the filament becomes dark in color and thin walled losing its contents. Gradually it becomes biconcave in shape and degenerates ultimately. Soon after, branching follows, producing a pair of branches . . (3) By the formation of Loop: It is also due to the formation of loops resulting in branching of Scytonema. The filament bulges out and forms a loop. One of the terminal cells in the loop degenerates and this results 'in two pseudobranches. (4) By the Heterocysts: The trichome breaks up at the point of' heterocyst and branching follows the usual pattern. Cell structure: Cells are cylindrical or rectangular in shape. Heterocysts are found in between the vegetative cells. The terminal cell is usually hemisperical There is no sexual reproduction: Asexual reproduction is brought about by the formation of hormogonia. A hormogonium is formed by the formation of two separation discs. Each one consists of several cells and germinates into a new filament. 11 Classification Kingdom: Bacteria Division: Cyanobacteria Class: see taxonomic note Order: Nostocales Family: Nostocaceae Genus: Nostoc Species: Nostoc calcicola, N. commune, N. cycadae, N. desertorum, N. edaphicum, N. ellipsosporum, N. entophytum, N. flagelliforme, N. indistinguenda, N. lichenoides, N. linckia, N. muscorum, N. paludosum, N. piscinale, N. punctiforme, N. sphaericum. Nostoc is a genus of fresh water cyanobacteria that forms spherical colonies composed of filaments of moniliform cells in a gelatinous sheath. When on the ground, a Nostoc colony is ordinarily not seen; but after a rain it swells up into a conspicuous jellylike mass, which was once thought to have fallen from the sky, whence the popular names, fallen star and star jelly. It is also called witches' butter (not to be confused with the fungus Tremella mesenterica). Michael Quinion of the World Wide Words newsletter says that it is known in Welsh as pwdre sêr, or rot of the stars.[1] Nostoc can be found on moist rocks, at the bottom of lakes and springs, and rarely in marine habitats. It may also grow symbiotically within the tissues of plants, such as the aquatic fern Azolla (mosquito fern) or hornworts, providing nitrogen to its host. These bacteria contain photosynthetic pigments in their cytoplasm to perform photosynthesis. Description and Significance Nostoc is a diverse genus of simple algae, belonging to the blue-green group. They are found in gelatinous colonies, composed of filaments called "trichomes" surrounded by a thin sheath. They are common in both aquatic and terrestrial habitats. These organisms are known for their unusual ability to lie dormant for long periods of time and abruptly recover metabolic activity when rehydrated with liquid water. The bacteria's ability to withstand freezing and thawing cycles make them well-adapted to living in extreme environments, such as the Arctic and Antarctica. They can fix atmospheric nitrogen, making them good candidates for environments with low nitrogen rates. Nostoc, first discovered in the 19th century, is one of the most widespread phototrophic bacteria in the world. As a nitrogen fixer, these bacteria may provide plants with important nutrients and therefore can be used agriculturally. In 1988 a terrestrial species, Nostoc commune, was found to harbor a previously unidentified UV-A/B absorbing pigment. This protective pigment has enabled them to survive not only while under hydration-related stress, but in areas of extreme UV radiation as well. Genome Structure Nostoc's genetics are worth studying because of the genus' unique adaptations which allow them to survive and even thrive in extreme environments. Also, a better understanding of soil-dwelling nitrogen fixers such as Nostoc may help advance fertilizer production and benefit agriculturalists. 12 Cell Structure and Metabolism Nostocs are photosynthesizers which use cytoplasmic photosynthetic pigments rather than chloroplasts in their metabolic process. They are single-celled, and lack a nucleus or other internal membrane systems; their cytoplasm is composed 70%-85% of water. The cells do not possess flagella, but are motile by a swaying motion. Division is by binary fission; some branching may occur. The cells form filamentous structures known as trichomes, which in turn make up colonies encased by a thin sheath; these colonies may be mat-like or spherical and are either micro- or macroscopic-spherical colonies may reach sizes of up to 2.6 kg wet weight. * * * * * * * *** Anabaena Class : Cyanophyceae Order: Nostacales Family: Nostacaceae Genus : Anabaena Occurrence: Commonly found in aqatic environments . They grow luxiantly in the rice fields .many species are plankonic in lakes and ponds and impart a blue-green coloration to water. Symbiotic members are found in association with Azolla. Thallus Structure: A thallus may consists of single filament with or without individual envelop . In the planktonic forms , it may consists of many filaments entangled and enclosed in a delicate mucilagenous matrix of indefinite shape . The sheath surronding trichomes are always hyaline and diffluent, narrow or broad. Mostly thalli are free floating in water or form a mucilagenous stream on the surface of moist soil. Filaments are usually straight . Planktonic species may be coiled or irregularly contorted .Filaments are uniserate , in which the cells are arranged end to end. Trichomes are uniformly broad throughout. The cells are cylindrical but spherical cells are also present in some species. Pseudovacuoles are present in planktonic forms. The protoplasm is either homogenous or granular and cell contents are blue-green , gray or variously colored. Heterocysts are also found.Heterocyst are usually of the same shape and slightly larger than vegetative cells.In many species a restricted number of akinetes occur adjacent to heterocysts. In other species , they develop in long chains and away fron heterocysts. Reproduction : 13 a) By Hormogonia:Hormogonia are frequently formed due to breaking up of trichomes in to smaller pieces at the region of heterocysts. On germination , they develop heterocyst and grow into long filaments. They are motile in many species. b) By akinetes : in many species the number and position of akinetes is a specific character i.e; they may be one or two or more in chains .Further , they may be on one or both the sides of a heterocyst or may be disposed away from it. Akinetes are generally cylindrical but may be spherical .On germination , akinetes form a few celled hormogonia. Nitrogen deficiency appears to be one factor leading to production of akinetes. c) By germination of heterocysts:In a few species , heterocyst germinate to form a few celled hormogonia . The pigments reappear and cell contents transform into a germling. Anabaena possess motile stage in the vegetative pase and the non motile phase is merely a response to adverse conditions. Fixation of nitrogen has been convingly demonstrated in species of A.cylindrica, A. ambigua, A. naviculodes Stigonema Order:Stigonamatales Family:Stigonemataceae Genus:Stigonema Occurence: Stigonema is widely distributed on sub aerial situations including moist rocks and tree trunks . Some species grow on moist soils and some other as free floating forms in fresh water reservoirs . The algae often grows mixed with mosses and smaller plants . Some species are known to dwell as the algal component in certain lichens. Thallus Structure: Thallus possess a filamentous and hetertrichous habit. The filaments are variuosly curved and irregularly branched . Many species have densely interwined filaments and form a thin cushion like growth. The branches may arise from all sides of the main axis and may branch and rebranch repeatedly . The branches are true branch and lateral which retain the continuity with the main filament. In inition of a true branch, the cell divides parallel to the axis of the filament . The upper cell as grows, is accommodated within simultaneously growing sheath and it does not come out . in the young filaments , the sheath remains close to the trichome while in mature filaments it becomes wide , firm and mucilagenous . The sheath may remain hyaline and homogenous or mor e commonly it may become lamellated and develop yellow brown or black colour. In early stage , the filaments may be uniserate but later stage they become multiserrate due to longitudinal divisions of cells. The cells are characteristically spherical but in young axis and branches they are flattened due to mutual pressure. In old filaments the lie at a distance fro one another and these cells may be surronded by their individual sheath. In mature filaments ,adjacent sister cells are connected with each other by pit connections. Heterocysts are present both in the main filament as well as in the branches . They may be terminal or intercalary but lateral in position 14 The filament exhibit apical growth .In many species , as the filament becomes older , the pit connections are withdrawn , cells divide to form independent pockets with lamellated envelop. Reproduction: In the multiserate species ,ther are two kinds of true lateral branches . Some branches are of limited growth , multiserate and are purely vegetative . Others are short with club-shaped apieces , uniserate and produce a few celled hormogonia . The hormogonia groe independently and form new filaments . Fixation of elementary nitrogen has been recorded in Stigonema informe ******** 15 16