Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Catalytic triad wikipedia , lookup
Lipid signaling wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Biochemistry wikipedia , lookup
Biochemical cascade wikipedia , lookup
Signal transduction wikipedia , lookup
Paracrine signalling wikipedia , lookup
Phosphorylation wikipedia , lookup
Drug discovery wikipedia , lookup
Ultrasensitivity wikipedia , lookup
MTOR inhibitors wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Citric acid cycle wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Enzyme inhibitor wikipedia , lookup
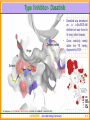
Structural Insights into Kinase Inhibition Ramesh Sistla and Subramanya H.S. Aurigene Discovery Technologies Ltd. #39-40, KIADB Industrial Area, Electronic City Phase II Bangalore 560 100 Kinases - Introduction • Kinases are enzymes catalyze phosphorylation • that ATP + protein = ADP + phosphoprotein • Key signaling enzyme • Human genome encodes > 500 kinases - Kinome • They have been implicated in different diseases including cancer, metabolic disorders and central nervous system indications. • Depending on the amino acid a kinase phosphorylates, they are known as Serine/Threonine or Tyorsine kinases. AURIGENE……Acccelerating Discovery www.cellsignal.com 2 Signaling Cascades • The figure shows the involvement of kinases in cell proliferation and survival. • In this cascade the phosphorylation of each kinase by its upstream kinase serves as a signal for downstream activity. • Inhibiting the pathway through inhibition of kinase involved in the pathway is an attractive proposition Current Medicinal Chemistry, 2008 Vol. 15, No. 29 3037 AURIGENE……Acccelerating Discovery 3 Promise of Kinase Inhibitors Some Advanced Kinase Inhibitors Druggable Genome Structure Identifier Target F F Kinome N N F O H N AMN-107 BCR/ABL N H N H 3C N N N N H N Imatinib CH 3 N O H 3C CH 3 N H STI-571 N BCR/ABL N Dasatinib CH 3 Cl O N H N S N N N H N N O O HC H N N BCR/ABL + OSI-774 Erbb OH CH 3 O BMS354825 CH 3 O CH 3 N • Kinases are an attractive target class – Druggability – Early successes (FDA approval of some of the kinase inhibitors) • Possibility of structure guided design – Large number of crystal structures in complex with inhibitors are available AURIGENE……Acccelerating Discovery 4 General Structure of Kinases N-terminal lobe • Bi-lobial structure • N-termial lobe – Mainly made of beta-sheets and connecting loops – One functionally important helix C-terminal lobe • Both lobes joined by a loop called hinge. • ATP binding pocket is in the interface between the lobes • C-terminal lobe – Mainly made of α-helices • Activation loop spans both Nand C-terminal lobes AURIGENE……Acccelerating Discovery 5 Important Structural Elements …GxGxxG… Helix-C Hinge • Plays an important role in catalysis Hinge – • Closes in on the ATP Helix C – • DFG……APE loop Glycine rich loop – • ATP Adenosine moiety of the ATP makes bidentate H-bond with this region Activation loop – Starts with conserved sequence DFG and ends with APE. AURIGENE……Acccelerating Discovery 6 Binding of ATP and Catalysis Orientation of the DFG motif critical for the phosphorylation H-bonds Hinge Metal Metal γ-phosphate coordinates with the metal S •Activation loop (DFG……APE) provides docking site for the substrate •Highly disordered and usually unresolved in the x-ray structures AURIGENE……Acccelerating Discovery T Substrate Y Phosphate 7 Important Residues Close up of the catalytic machinery N-terminal lobe Helix-C ATP Lys Salt bridge Asp Water Metal C-terminal lobe • • Glu In the active conformation of the kinases, a conserved Lys residue makes a salt bridge with a conserved Glu residue in the middle of the helix-C. This interaction ensures the positioning of the amino acid Asp (of the DFG motif) to coordinate with the γ-phosphate, the divalent metal ion and catalytic water molecule to facilitate catalysis AURIGENE……Acccelerating Discovery 8 Kinase Inhibitors • In most cases, inhibitors compete with ATP in order to inhibit the kinase – Such inhibitors are ATP mimetics in the sense that they make interactions similar to what ATP makes. G-loop Hinge ATP Inhibitor Ribose pocket Phosphate pocket ATP Inhibitor AURIGENE……Acccelerating Discovery 9 Various Subsites in Kinases Hinge Gatekeeper Deeper cavity Inhibitor Solvent ATP Schematic of the binding pockets PDB: 2C6E/1MQ4 An example of a kinase inhibitor bound in the ATP pocket is shown. Apart from hinge region interaction and solvent interaction, the inhibitor occupies a deeper hydrophobic cavity, also known as selectivity pocket Size of an amino acid preceding the hinge region controls the accessibility to the deeper pocket – Gatekeeper, (Typically Met/Leu/Thr/Ile/Tyr) AURIGENE……Acccelerating Discovery 10 Type I Inhibitor- Dasatinib • Deeper pocket Hinge • Dasatinib was developed as a c-Src/BCR-Abl inhibitor but was found to hit many other kinases. Cross reactivity mainly within the TK family; Approved by FDA Solvent 1nM 10nM 100nM 1μM 10μM Ref: Karaman et. al., NATURE BIOTECHNOLOGY VOLUME 26 NUMBER 1 JANUARY 2008 AURIGENE……Acccelerating Discovery 11 DFG-IN vs DFG-OUT Helix-C Gly rich loop DFG-Out DFG-In • The activation loop (DFG….APE) has to be IN when the kinase is active – DFG “in” conformation • The DFG loop has been shown to be in an “out” position when kinases are inactive. • This can be exploited in the design of inhibitors. AURIGENE……Acccelerating Discovery 12 DFG-IN vs DFG-OUT Helix-C ATP Gleevec DFG OUT DFG IN • Differences between DFG IN and DFG OUT structures are exemplified. • DFG loop in OUT position will clash with phosphate of ATP • When DFG moves to OUT helix-C also moves away creating the pocket shown by bold red arrow. • Gleevec binds to the DFG-OUT conformation of the C-Abl kinase. PDB:1T46 AURIGENE……Acccelerating Discovery 13 Example of Type-II Inhbition Hinge Phe-out conformation Schematic of the binding pockets PDB:1KV1 BIRB-796 binds to p-38 in the Phe-out conformation • The doublet of H-bonds with E-111 (helix-C) and D-207 (DFG loop) backbone is very important • Hence a urea or amide is the common feature in these inhibitors Ref: Karaman et. al., NATURE BIOTECHNOLOGY VOLUME 26 NUMBER 1 JANUARY 2008 AURIGENE……Acccelerating Discovery 14 R Some Known DFG OUT Inhibitors 2oo8 Tie – DFG out J.Med.Chem. 50: 611-626 2p4i Tie – DFG out Bioorg.Med.Chem.Lett. 17: 2886-2889 2osc Tie – DFG out R Bioorg.Med.Chem.Lett. 17: 2886-2889 N H 2og8 Lck – DFG out O 2ofv Lck – DFG out J.Med.Chem. 50: 611-626 2p2i KDR – DFG out Apart from a hinge binding group, the common feature in these molecules is existence of the bi-aryl amide/urea group which makes interaction with Glu (helix-C) and Asp (DFG loop) AURIGENE……Acccelerating Discovery 15 Allosteric Kinase Inhibition – Type III Helix-C ATP DFG loop • Certain kinases have an allosteric pocket in which an inhibitor can co-bind with ATP • The phosphorylation of the substrate is prevented by unavailability of the catalytic Asp • There are no hinge region interactions in these inhibitors. AURIGENE……Acccelerating Discovery 16 A Still Different Type of Inhibitor? • Recently Merck published the co-crystal structure of CHK1 kinase with an inhibitor that is bounds far away from the active site. • DFG loop is has IN conformation, but the inhibitor probably occupies substrate binding site. • Such inhibitors are not being designed yet. They could be results of HTS campaigns. PDB:3F9N AURIGENE……Acccelerating Discovery 17 SBDD at Aurigene All the structural biology efforts are to aid in more focused medicinal chemistry AURIGENE……Acccelerating Discovery 18