Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Gluten immunochemistry wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Immune system wikipedia , lookup
IgA nephropathy wikipedia , lookup
Innate immune system wikipedia , lookup
DNA vaccination wikipedia , lookup
Autoimmune encephalitis wikipedia , lookup
Duffy antigen system wikipedia , lookup
Complement system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Immunocontraception wikipedia , lookup
Adaptive immune system wikipedia , lookup
Molecular mimicry wikipedia , lookup
Anti-nuclear antibody wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
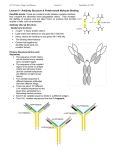
Antigens/Antibodies Chapters 4-6 Antigen vs. Immunogen • Antigen – Any substance that can bind to an antibody or T cell receptor • Immunogen – Any substance that can elicit an immune response – All immunogens are antigens – Not all antigens are immunogens, i.e. haptens Immunogenicity of Antigens • Determined by – – – – Foreignness Molecular Size Chemical Composition Degradability • Influenced by – Host genotype – Dose and route of administration – Presence of adjuvants Epitopes • Discrete sites on immunogens recognized by antibodies or T cell receptors • Macromolecules contain many epitopes with different immunogenicities • Immunogenicity can vary between individuals and populations. • Some epitopes are immunodominant • Identification of B-cell epitopes within the HPV16 E7 protein. • Overlapping peptides were generated and injected into mice. • The antibody response to each peptide was measured by ELISA. • Comerford, S.A. et al. J. Virol. 65:4681-4690, 1991 Epitopes • Overlapping • Non-overlapping • • • • Allosteric Linear Conformational Neoantigen B and T Cell Epitopes Can Overlap •Comerford, S.A. et al. J. Virol. 65:4681-4690, 1991 Antibodies Objectives • To discuss the general properties of all immunoglobulins • To describe the basic structure of immunoglobulins • To relate immunoglobulin structure with function • To define immunoglobulin hypervariable and framework regions • To define immunoglobulin classes and subclasses, types and subtypes • To describe the structures and properties of immunoglobulin classes Immunoglobulin (Ig) • Immunoglobulins are glycoprotein molecules that are produced by plasma cells in response to an immunogen and which function as antibodies. The immunoglobulins derive their name from the finding that they migrate with globular proteins when antibody-containing serum is placed in an electrical field. Electrophoretic separation of serum proteins General Functions • Antigen Binding • Effector Functions GENERAL FUNCTIONS OF IMMUNOGLOBULINS • Antigen binding – Immunoglobulins bind specifically to one or a few closely related antigens. – Binds to a specific antigenic determinant. – Antigen binding is the primary function of antibodies and can result in protection of the host. – The “valency” of antibody refers to the number of antigenic determinants that an individual antibody molecule can bind. – The valency of all antibodies is at least two and in some instances more. Effector Functions • The immunoglobulins mediate a variety of these effector functions: – Fixation of complement - This results in lysis of cells – Binding to various cell types: • Phagocytic cells, lymphocytes, platelets, mast cells, and basophils have receptors that bind immunoglobulins. This binding can activate the cells to perform some function. • Some immunoglobulins also bind to receptors on placental trophoblasts, which results in transfer of the immunoglobulin across the placenta. As a result, the transferred maternal antibodies provide immunity to the fetus and newborn BASIC STRUCTURE OF IMMUNOGLOBULINS • Heavy and light chains • Disulphide bonds • Variable (V) and constant (C) regions • Hinge region • Domains • Oligosaccharides • IgG structure – 2 HEAVY chains = red and yellow – 2 LIGHT chains = green and blue. • 2 antiparallel beta sheets – make up each of the immunoglobulin domains (2 Ig domains per L chain and 4 domains per H chain) • Papain cleavage site. – Papain cleavage gives ONE Fc fragment (2 immunoglobulin domains from each H chain) and – TWO Fab fragments (each with 2 immunoglobulin domains from an H chain and 2 immunoglobulin domains from an L chain) • each Fab fragment contains ONE antigen binding site plot of variability as a function of position along the sequence of the amino acids that make up the heavy chain of Ig molecules. STRUCTURE OF THE VARIABLE REGION • Hypervariable (HVR) or complementarity determining regions (CDR) – Comparisons of the amino acid sequences of the variable regions of immunoglobulins show that most of the variability resides in three regions called the HVR/CDR. – Antibodies with different specificities (i.e. different combining sites) have different complementarity determining regions while antibodies of the exact same specificity have identical complementarity determining regions (i.e. CDR is the antibody combining site). – Complementarity determining regions are found in both the H and the L chains. STRUCTURE OF THE VARIABLE REGION • Framework regions • The FR regions form a beta-sheet structure which serves as a scaffold to hold the HV regions in position to contact antigen. • Based on similarities and differences in the framework regions the immunoglobulin heavy and light chain variable regions can be divided into groups and subgroups. • Antibodies are divided into five major classes, IgM, IgG, Iga, IgD, and IgE, based on their framework/constant region structure. HUMAN IMMUNOGLOBULIN CLASSES, SUBCLASSES, TYPES AND SUBTYPES • Immunoglobulin classes The immunoglobulins can be divided into five different classes, based on differences in the amino acid sequences in the constant region of the heavy chains. • All immunoglobulins within a given class will have very similar heavy chain constant regions. These differences can be detected by sequence studies or more commonly by serological means (i.e. by the use of antibodies directed to these differences). – – – – – IgG - Gamma heavy chains IgM - Mu heavy chains IgA - Alpha heavy chains IgD - Delta heavy chains IgE - Epsilon heavy chains Immunoglobulin Subclasses • The classes of immunoglobulins can de divided into subclasses based on small differences in the amino acid sequences in the constant region of the heavy chains. • All immunoglobulins within a subclass will have very similar heavy chain constant region amino acid sequences. Again these differences are most commonly detected by serological means. • IgG Subclasses – – – – IgG1 - Gamma 1 IgG2 - Gamma 2 IgG3 - Gamma 3 IgG4 - Gamma 4 heavy chains heavy chains heavy chains heavy chains • IgA Subclasses – IgA1 - Alpha 1 heavy chains – IgA2 - Alpha 2 heavy chains IgG • Structure – All IgG's are monomers (7S immunoglobulin). The subclasses differ in the number of disulfide bonds and length of the hinge region. Properties • IgG is the most versatile immunoglobulin because it is capable of carrying out all of the functions of immunoglobulin molecules. • IgG is the major Ig in serum - 75% of serum Ig is IgG • IgG is the major Ig in extra vascular spaces • Placental transfer - IgG is the only class of Ig that crosses the placenta. Transfer is mediated by a receptor on placental cells for the Fc region of IgG. Not all subclasses cross equally well; IgG2 does not cross well. • Fixes complement - Not all subclasses fix equally well; IgG4 does not fix complement • Binding to cells - Macrophages, monocytes, granulocytes and some lymphocytes have Fc receptors for the Fc region of IgG. Not all subclasses bind equally well; IgG2 and IgG4 do not bind to Fc receptors. A consequence of binding to the Fc receptors on PMNs, monocytes and macrophages is that the cell can now internalize the antigen better. The antibody has prepared the antigen for eating by the phagocytic cells. • IgG is a good opsonin. IgG • • • • • • • • Increases in: Chronic granulomatous infections Infections of all types Hyperimmunization Liver disease Malnutrition (severe) Dysproteinemia Disease associated with hypersensitivity granulomas, dermatologic disorders, and IgG myeloma Rheumatoid arthritis • • • • • • Decreases in: Agammaglobulinemia Lymphoid aplasia Selective IgG, IgA deficiency IgA myeloma Bence Jones proteinemia Chronic lymphoblastic leukemia Antibodies Continued Reading • Chapter 4 Kuby et al, Immunology 6th Edition • Chapter 4 Abbas et al, Cellular and Molecular Immunology 6th Edition. IgM • Structure: – Is a pentamer (19S Ig) but it can also exist as a monomer. – In the pentameric form all heavy chains are identical and all light chains are identical. – IgM has an extra domain on the mu chain (CH4) and it has another protein covalently bound via a S-S bond called the J chain. This chain functions in polymerization of the molecule into a pentamer. Properties • Third most common serum Ig. • IgM is the first Ig to be made by the fetus and the first Ig to be made by a virgin B cells when it is stimulated by antigen. • Pentameric structure, IgM is a good complement fixing Ig - IgM antibodies are very efficient in leading to the lysis of microorganisms. • IgM is also a good agglutinating Ig - IgM antibodies are very good in clumping microorganisms for eventual elimination from the body. • IgM binds to some cells via Fc receptors. Surface IgM Monomer No J chain Extra 20 aa in tail Associated with two additional proteins involved in signal transduction IgA • Structure – Serum IgA is a monomer but IgA found in secretions is a dimer – When IgA exits as a dimer, a J chain is associated with it. – When IgA is found in secretions is also has another protein associated with it called the secretory piece or T piece – IgA is sometimes referred to as 11S immunoglobulin. • Unlike the remainder of the IgA which is made in the plasma cell, the secretory piece is made in epithelial cells and is added to the IgA as it passes into the secretions. The secretory piece helps IgA to be transported across mucosa and also protects it from degradation in the secretions. Properties • IgA is the 2nd most common serum Ig. • IgA is the major class of Ig in secretions - tears, saliva, colostrum, mucus. Since it is found in secretions secretory IgA is important in local (mucosal) immunity. • Normally IgA does not fix complement, unless aggregated. • IgA can binding to some cells - PMN's and some lymphocytes. IgD • Structure – Exists only as a monomer. • Properties – IgD is found in low levels in serum; its role in serum uncertain. – IgD is primarily found on B cell surfaces where it functions as a receptor for antigen. – IgD on the surface of B cells has extra amino acids at C-terminal end for anchoring to the membrane. – It also associates with the Igalpha and Ig-beta chains. – IgD does not bind complement. IgE • Structure – IgE exists as a monomer and has an extra domain in the constant region. • Properties – IgE is the least common serum Ig since it binds very tightly to Fc receptors on basophils and mast cells even before interacting with antigen. – Involved in allergic reactions - binding of the allergen to the IgE on the cells results in the release of various pharmacological mediators that result in allergic symptoms. – IgE also plays a role in parasitic helminth diseases. Since serum IgE levels rise in parasitic diseases, measuring IgE levels is helpful in diagnosing parasitic infections. Eosinophils have Fc receptors for IgE and binding of eosinophils to IgE-coated helminths results in killing of the parasite. • d) IgE does not fix complement. Isotypes of Light Chain • Kappa (κ) and Lamda (λ), which are distinguished by their carboxyterminal constant (C) regions. • An antibody molecule has either two κ light chains or two λ light chains, but never one of each. • In humans, about 60% of antibody molecules have κ light chains, and about 40% have λ light chains. • Changes in this ratio can occur in patients with monoclonal B cell tumors because the neoplastic clone produces antibody molecules with the same light chain. • In fact, the ratio of κ-bearing cells to λ-bearing cells is often used clinically in the diagnosis of B cell lymphomas. • In mice, κ-containing antibodies are about 10 times more abundant than λ-containing antibodies. • Unlike in heavy chain isotypes, there are no known differences in function between κ-containing antibodies and λ-containing antibodies. Bringing together information Natural Distribution and Production of Antibodies • Found in biological fluids throughout the body • On surface of limited number of cells • B cells are the only cells that synthesize antibodies Interesting facts • A healthy 70-kg adult human produces about 2g3g of antibodies every day. • Almost two thirds of this is IgA, which is produced by activated B cells and plasma cells in the walls of the gastrointestinal and respiratory tracts and actively transported into the lumens. • The large amount of IgA produced reflects the large surface areas of these organs. • Antibodies that enter the circulation have limited half-lives. • After exposure to an antigen, much of the initial antibody response occurs in lymphoid tissues: – the spleen, – lymph nodes, – mucosal lymphoid tissues, • Secreted forms of antibodies accumulate in the plasma (the fluid portion of the blood), in mucosal secretions, and in the interstitial fluid of tissues. • Secreted antibodies often attach to the surface of other immune effector cells, such as mononuclear phagocytes, natural killer (NK) cells, and mast cells, which have specific receptors for binding antibody molecules. Looking at Structure of Ig Molecules • Breakthrough came when researchers discovered that: – multiple myeloma patients produced biochemically identical antibody molecules • Led to an extremely powerful technique for producing monoclonal antibodies, described by Georges Köhler and Cesar Milstein in 1975. – They developed a method for immortalizing individual antibody-secreting cells from an immunized animal by producing "hybridomas," each of which secreted individual monoclonal antibodies of predetermined specificity. Antigen-antibody complexes are held together by non-covalent forces (therefore, antigen binding by antibody is reversible) Affinity of Igs • The strength of the binding between a single combining site of an antibody and an epitope of an antigen is called the affinity of the antibody. • The affinity is commonly represented by a dissociation constant (Kd), which indicates how easy it is to separate an antigen-antibody complex into its constituents. • A smaller Kd indicates a stronger or higher-affinity interaction because a lower concentration of antigen and of antibody is required for complex formation. • The Kd of antibodies produced in typical humoral immune responses usually varies from about 10-7M to 10-11M. • Serum from an immunized individual will contain a mixture of antibodies with different affinities for the antigen, depending primarily on the amino acid sequences of the CDRs. Avidity of Igs • Polyvalent antigens will have more than one copy of a particular determinant. • Although the affinity of any one antigen-binding site will be the same for each epitope of a polyvalent antigen, the strength of attachment of the antibody to the antigen must take into account binding of all the sites to all the available epitopes. • This overall strength of attachment is called the avidity and is much greater than the affinity of any one antigen-binding site. • Thus, a low-affinity IgM molecule can still bind tightly to a polyvalent antigen because many low-affinity interactions (up to 10 per IgM molecule) can produce a single high-avidity interaction. Antigen-Ig Interactions • Specificity – antibodies generated in response to the antigens of one microbe usually do not react with structurally similar self molecules or with the antigens of other microbes – biochemical constituents of all living organisms are fundamentally similar – However, some antibodies produced against one antigen may bind to a different but structurally related antigen. – This is referred to as a cross-reaction. – Antibodies that are produced in response to a microbial antigen sometimes cross-react with self antigens, and this may be the basis of certain immunologic diseases Antigen-Ig Interactions • Diversity – The ability of antibodies in any individual to specifically bind a large number of different antigens is a reflection of antibody diversity – Total collection of antibodies with different specificities represents the antibody repertoire. – The genetic mechanisms that generate such a large antibody repertoire occur exclusively in lymphocytes. – They are based on the random recombination of a limited set of inherited germline DNA sequences into functional genes that encode the V regions of heavy and light chains as well as on the addition of nucleotide sequences during the recombination process Antigen-Ig Interactions • Affinity Maturation – The ability of antibodies to neutralize toxins and infectious microbes is dependent on tight binding of the antibodies. Tight binding is achieved by high-affinity and high-avidity interactions. – Generation of high-affinity antibodies involves subtle changes in the structure of the V regions of antibodies during T cell-dependent humoral immune responses to protein antigens. – These changes come about by a process of somatic mutation in antigenstimulated B lymphocytes that generates new V domain structures, some of which bind the antigen with greater affinity than did the original V domains – Those B cells producing higher-affinity antibodies preferentially bind to the antigen and, as a result of selection, become the dominant B cells with each subsequent exposure to the antigen. – Called affinity maturation, results in an increase in the average binding affinity of antibodies for an antigen as a humoral immune response evolves. – An antibody produced during a primary immune response often has a Kd in the range of 10-7 to 10-9M and in secondary responses, the affinity increases, with a Kd of 10-11M or less. The Immunoglobulin Superfamily • Many of the cell surface and soluble molecules that mediate recognition, adhesion, or binding functions in the vertebrate immune system share partial amino acid sequence homology and tertiary structural features that were originally identified in Ig heavy and light chains. • Features are found in many molecules outside the immune system that also perform similar functions. • These diverse proteins are members of the Ig superfamily (sometimes called the Ig supergene family). • A superfamily is broadly defined as a group of proteins that share a certain degree of sequence homology, usually at least 15%. • The conserved sequences shared by superfamily members often contribute to the formation of compact tertiary structures referred to as domains.