Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Pre-Bötzinger complex wikipedia , lookup
Patch clamp wikipedia , lookup
Threshold potential wikipedia , lookup
Membrane potential wikipedia , lookup
Alveolar macrophage wikipedia , lookup
Resting potential wikipedia , lookup
Electrophysiology wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Cardiac action potential wikipedia , lookup
End-plate potential wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
G protein-gated ion channel wikipedia , lookup
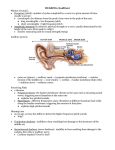
Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting Problem 1.12: Special Senses Topic: Biochemical Aspects of Tasting Prepared by: Joseph Mak 1) Overview - The elementary role of the gustatory system is to distinguish between food and potential toxins, two components are required to accomplish this task: (i) - - - A detection system of receptor cells capable of responding to the great diversity of substances in the environment that might be ingested (ii) Neuronal pathways that refer taste information to appropriate cortical structures Pleasing sensations associated with food are necessary to maintain the appetite and to initiate appropriate digestive and respiratory responses; Unpleasing sensations associated with potential toxins elicit protective reflexes, such as coughing, sneezing, gagging, or vomiting Some taste responses are inborn, eg, a preference for sweetness and an aversion to bitter tastes. However, the gustatory system is highly modifiable by experience. Illness that occurs soon after ingestion of a particular food can greatly diminish subsequent preferences for that food Taste can be acquired so that some bitter tastes, like quinine, are tolerated or even enjoyed. Taste receptors are classified as modified epithelial cells rather than true neurons. They occur in clustered structures called taste buds, each of which contains 50 to 100 receptor cells arranged like slices of an orange with a central opening or pore open to the surface of the tongue. The taste bud also contains basal cells that are the stem cells for the production of new taste receptors The typical life span of taste receptors is 1 to 2 weeks. Taste receptors synapse on dendrites of afferent gustatory fibres projecting into the taste bud. 2) The Five Basic Tastes - - - Humans perceive five basic tastes: salt, sour, sweet, bitter and umami. Further distinctions are made based on combinations of these basic sensations, and subtleties can be made by combinations of smell and taste, and to a lesser extent the temperature and texture of food The five basic tastes are derived from different transduction mechanisms located on taste receptor cells and their specific interactions with different types of molecules In general, (i) Acids elicit a sour taste (ii) Salt elicits a salty taste (iii) Sugars, some proteins, and amino acid artificial sweeteners such as saccharin and aspartame invoke sweetness Page 1 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting (iv) (v) - Ions such as potassium and magnesium and organic compounds such as quinine and coffee taste bitter Umami, recently recognized as an additional basic taste, is associated with some amino acids such as glutamate, the common culinary form of which is monosodium glutamate, commonly known as MSG. These compounds interact with taste receptors and cause an increased release of transmitter, which in turn stimulates primary afferent taste fibres. There are four mechanisms by which chemicals cause increased transmitter release from taste receptors (i) (ii) (iii) (iv) direct passage of ions through ion channels blockage of ionic channels opening of ionic channels activation of second-messenger systems through ligand interactions with membrane receptors 2.1 Salt - - - Sodium ions are largely responsible for establishing a salty taste Sodium ions diffuse through the pore of the taste bud and enter the taste receptor cell through sodium-selective channels present in the cell membrane. These channels are characterized by their sensitivity to the drug amiloride and their insensitivity to changes in voltage. Thus they are different from sodium channels involved in propagation of action potentials The entry of positively charged ions depolarizes the taste receptor cell, opens voltage-dependent calcium channels, and increases the influx of calcium, thereby allowing calcium to enter the cell and cause the release of neurotransmitter Hydrogen ions can also enter these channels, therefore acidic food has a salty taste 2.2 Sour - - A sour taste is associated with acidic substances that affect sour-sensitive taste receptors in two ways. (i) Acids in solution generate hydrogen ions that can permeate the amiloridesensitive sodium channel described and cause depolarization-stimulated release of neurotransmitter in the same manner as sodium ions. (ii) Hydrogen ions also block a potassium-selective channel within the membrane. This also causes depolarization because the normal movement of potassium out of the cell is blocked, and more positively charged potassium ions are trapped intracellulary (Since intracellular content of K+ is high and is continually lost from the cell, as these potassium channels are blocked, K+ then accumulates inside the cell, and hence causes depolarization) Foods that cause depolarization and increased transmitter release through both of these mechanisms are perceived as sour Page 2 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting - Those that cause depolarization only through diffusion of cations through the sodium channel are perceived as salty 2.3) Sweet - - There are specific membrane receptor proteins on some taste cells that bind sugars and other sweet-tasting substances. Binding to these receptor sites activates a second messenger system similar to the one associated with noradrenergic receptors. When the receptor protein is activated it activates a Gs-protein, which stimulates adenylyl cyclase to produce cAMP. This cAMP releases protein kinase A (PKA) from its regulator proteins, and PKA phosphorylates potassium channels, closing them and depolarizing the cell. (Details of which is dealt with in later sections) As a result, sweet-sensitive taste cells are depolarized 2.4) Bitter - - Some bitter compounds (eg, calcium ions and quinine) decrease conductance of potassium-selective channels similar to the mechanisms used for detection of sweetness Other bitter substances bind to specific membrane receptors that activate secondary-messenger systems and cause membrane depolarization One type of bitterness receptor triggers an increased production of the intracellular messenger, inositol triphosphate (IP3) In all other transduction mechanisms described earlier, depolarization of the receptor cell causes an increase in calcium influx through votage-sensitive calcium channels, and calcium ions act as the trigger for release of neurotransmitter. This is not so in the case of the IP3 transduction mechanism Here, the membrane potential is not altered; rather, IP3 causes the release of calcium from internal storage sites, which in turn directly stimulates neurotransmitter release. 2.4) Umami - This is a taste sensation associated with certain amino acids such as glutamate and perhaps arginine These amino acids bind to and activate a cation-permeable channel, causing depolarization in a similar manner to glutamate activation of cation channels in the brain Page 3 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting Fig 1.12-1 Transduction mechanisms for salt, sour, umami, sweet and bitter All taste receptors release neurotransmitters in response to an increase in free intracellular calcium, usually as a result of depolarization that opens reactive calcium channels. This cause of this depolarization varies with the specific receptor cell. (A) Salty taste is tranduced by amiloride-sensitive sodium channels that are always open. When the sodium concentration increases on the surface of the microvilli, it can enter the channels and depolarize the cell. Hydrogen ions can also enter these channels, and therefore acidic food has a salty taste. Page 4 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting (B) Sour taste is elicited when hydrogen ions act directly on open potassium channels, closing them and depolarizing the cell. (C ) Glutamate (eg, in MSG) binds to the receptor that opoens, allowing all small cations to pass, including sodium, potassium, and calcium. The net effect of these ionic movements is to depolarize the cell (D) Sweet tastes, such as sugars and certain proteins, produce depolarization by a chain of events that resemble many CNS neurotransduction systems. (dealt with in later sections) (E) There at least two types of bitter receptors. Quinine stimulates the bitter 1 receptor and closes potassium channels. (F) Other bitter substances bind with a bitter 2 receptor, which activates a Gprotein, which in turn causes the production of inositol triphosphate (IP3). IP3 triggers the release of calcium from internal stores, stimulating neurotransmitter release without an intervening depolarization. 3) Central Gustatory Pathways - - - - - Taste receptors synapse with dendritic elements of the primary afferent neurons of the gustratory pathway, causing the neurons to fire in response to appropriate stimuli above threshold levels These afferent fibres arise from cell bodies within three cranial nerve ganglia Afferent fibres within the facial nerve (cranial nerve VII) arise from the geniculate ganglia and innervate the anterior two-thirds of the tongue. Fibres from glossopharyngeal nerve (cranial nerve IX) arise from the inferior glossopharyngeal ganglia and innervate the posterior one third of the tongue Fibres from the vagus nerve (cranial nerve X) arise from the inferior vagal ganglia and innervate the smattering of taste buds found in throat regions, including the glottis, epiglottis and pharynx. Central projections from these three cranial nerve ganglia enter the brain stem along the lateral aspect of the medulla and converge to make synaptic contact with cells in the rostral or gustratory division of the solitary nucleus of the medulla. Ascending fibres from the solitary nucleus make a synaptic connection in the ventral posterior medial nucleus of the thalamus. Thalamic neurons then project to the primary gustratory cortex in the insular and orbitofrontal regions. The large number of distinguishable tastes is represented by population responses of various central taste neurons rather than by activation of individual tastespecific neurons ~ Coding for the four primary taste qualities is not based on complete selectivity of the chemoreceptors for the different qualities. ~ Instead, a given chemoreceptor responds to stimuli that evoke several different taste qualities, although perhaps most vigorously to one. Page 5 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting ~ Recognition of taste quality appears to depend on the patterned input from a population of chemoreceptors. The intensity of the stimulus is reflected in the total amount of evoked activity Fig 1.12-2 Central Gustratory Pathway Central projections of taste buds. Information from taste buds enters the ventral surface of the medulla via three pairs of cranial neurons and synapses in the rostral or gustatory division of the solitary nucleus. Secondary neurons carrying gustatory information project to the ventral posterior medial nucleus of the thalamus, where it is relayed to the cerebral cortex for conscious and appreciation. Here shows oral cavity and dorsal view of the brain stem showing the three cranial nerves that carry information is processed ipsilaterally. 4) Cellular Communication The above transduction mechanisms described are some means of cellular communication, this section deals with the basic mechanisms of them. This section is for better understanding of the biochemical sensation of taste. - - Cells respond to a variety of external stimuli and transduce these signals into electrical and biochemical changes. In many instances this involves the interaction of a signal with a receptor in the plasma membrane. This may lead directly to activation of ion channels, but more commonly it induces the production of intracellular messengers that then regulate cell function Important intracellular pathways include the cyclic adenosine monophosphate (cAMP) system, polyphosphoinositide system Page 6 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting - For chemical stimuli (eg, gustatory, olfactory, neurotransmitters, hormones, growth factors, etc) the first event leading to a cellular response is the interaction of the chemical signal (ligand) with its receptor. Receptors - Receptors are molecular entities, either proteins or glycoproteins, that bind ligands with high affinity Ligands - an organic molecule that donates the necessary eletrons to form coordinate covalent bonds with metallic ions, as oxygen is bound to the central atom of Hb. This term is also used to indicate any ion or molecule that reacts to form a complex with another molecule, frequently a macromolecule. Ion channels - Ion channels are membrane proteins that contain small, highly selective aqueous pores. They allow specific ions, eg, Na+, K+, Ca2+ or Cl- to move down their electrochemical gradients across the membrane, and are often named after the ions for which they show the greatest selectivity. - The major types of regulated ion channel include: (i) voltage-gated channels, in which the sensor responds to the voltage across the membrane, eg, the voltage-sensitive Na+ channel involved in action potential generation (ii) chemically gated channels, in which the sensor is sensitive to a chemical signal such as the binding of a ligand to a receptor that forms part of the channel, eg, the acetylcholine receptor channel involved in neuromuscular transmission; and (iii) mechanically gated channels, in which the sensor is sensitive to mechanical deformation of the membrane, eg, stretch-activated channels on sensory nerve terminals. 4.1 Intracellular messengers - The first such intracellular chemical to be indentified was cyclic adenosine monophosphate (cAMP), named a second messenger It is now known that many chemical stimuli result in an increase in concentration of cAMP in their target cells. Ca2+ is also an important intracellular messenger; stimuli may open channels allowing it to flow into the cell or be released from intracellular stores in endoplasmic reticulum. Other important intracellular messengers (e.g, cyclic GMP, IP3 and diacylglycerol) have since been identified. 4.1.1 G proteins - In both the cAMP and the polyphosphoinostide systems that are described in detail later in this section, the step that follows the binding of the ligand to its Page 7 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting - - receptor at the membrane surface, and that initiates the cell response, is the activation of a regulatory protein that binds GTP (Guanosine triphosphate) Hence this protein is called a G protein. About 100 different ligand-receptor complexes are now known to activate G proteins, which have come to be known as universal transducers G proteins consist of subunits. In the inactive state, the group of the G protein binds guanosine diphosphate (GDP), and the protein can be represented as “-GDP”. (See Fig 1.12-3) Following occupation of a surface receptor, a conformational change in the G protein results in the exchange of GDP for cytoplasmic GTP, giving -GTP. The -GTP subunit then dissociates from the -subunits and binds to an effector molecule (e.g, an enzyme such as adenylate-cyclase, or a channel protein) The -subunit has GTPase activity, it converts GTP back to GDP and is recombined with -subunits and the inactivated G protein is reconstituted. Fig 1.12-3 Role of the G protein in transduction. The ligand binds to the receptor in the plasma membrane and activates the G protein, which consists of - subunits. This allows the -subunit to exchange its bound GDP for GTP. The GTP then dissociates from the subunits and stimulates an effector (e.g, Adenylate cyclase, ion channel, etc) The GTPase on the -subunit breaks down GTP to GDP, causing the -GDP to dissociate from the effector and recombine with the -subunits. Page 8 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting 4.1.2 cAMP system - - - Adenylate cyclase (an enzyme) catalyses the synthesis of cAMP from adenosine triphosphate (ATP) Fig 1.12-4 summarizes the events that follow increased cAMP production. The raised level of cAMP within the cell stimulates protein kinase A, this may lead to specific changes in ion channel gating, enzyme activity, protein synthesis or gene activation, depending on the tissue involved. Eventually, the concentration of cAMP is restored to its basal level by degradation to AMP, this step being catalysed by the enzyme cyclic nucleotide phosphodiestease Importance of many hormone and neurotransmittors are mediated by cAMP: (i) Only certain tissues contain receptors which will react with a particular ligand to produce an increase in intracellular cAMP. (ii) The responses of various tissues to raised levels of cAMP are different. For example, in response to an increase in cAMP, liver cells break down glycogen but heart cells increase their rate and strength of contraction. Fig 1.12-4 Signals acting through cAMP. The ligand binds to its receptor (R) on the outer surface of the plasma membrane. At the inner surface are the GTPregulatory protein (G) and the enzyme adenylase cyclase (AC), which catalyses the conversion of ATP to cAMP. Binding of the ligand to the R-G complex allows the G unit to bind GTP and stimulate adenylate cyclase activity. cAMP activates protein kinase A, which phosphorylates cellular proteins to produce the characteristic cellular response. 4.1.3 Polyphosphoinositide system - Phosphoinositides are a family of plasma membrane phospholipids containing inositol. Binding of a hormone to its receptor activates a different G protein This in turn stimulates a phosphodiesterase (phospholipase C) in the plasma membrane, which cleaves PIP2 (phophatidylinositol 4,5-bisphophate) in the membrane to form IP3 (inositol trisphosphate) and DAG (diacylglycerol) Page 9 Problem 1.12: Special Senses Topic: Biochemical Aspect of Tasting - IP3 migrates to the endoplasmic reticulum where it stimulates the release of Ca2+ DAG remains in the membrane and, in the presence of Ca2+, activates protein kinase C. This enzyme phosphorylates many intracellular proteins, including some that are also phosphorylated by protein kinase A. Fig 1.12-5 Signals operating through the polyphosphoinositide system. The binding of the ligand to its receptor (R ) in the plasma membrane activates the enzyme phospholipase C (PLC) via the G protein. This enzyme cleaves PIP2 to IP3 and DAG. IP3 mobilizes Ca2+ from the endoplasmic reticulum while DAG, in the presence of Ca2+, activates the enzyme protein kinase C (PKC), and these actions lead to specific cellular responses. IP3 and DAG are rapidly metabolized and their end-products recycled to form PI, which is phosphylated to form PIP and finally PIP2. References: 1) Lecture Notes on Human Physiology 3rd Ed 612 L47 2) Esssential Medical Physiology 2nd Ed by Leonard R. Johnson D 612 E78 J Page 10