
Disorders of Acid
... The second line of defense against acid-base disturbances is the control of CO2 by the respiratory system. Excess CO2 or excess H+ ions in the blood mainly act directly on the respiratory center in the brain to control ventilation. Although H+ ions do not easily cross the blood–brain barrier, CO2 cr ...
... The second line of defense against acid-base disturbances is the control of CO2 by the respiratory system. Excess CO2 or excess H+ ions in the blood mainly act directly on the respiratory center in the brain to control ventilation. Although H+ ions do not easily cross the blood–brain barrier, CO2 cr ...

01 Mills
... Peripheral and central chemoreceptors provide inputs to the respiratory centres. The peripheral chemoreceptors lie in the carotid and aortic bodies. The carotid bodies are more important in stimulating ventilation, while the aortic bodies are also capable of responding to hypotension. The peripheral ...
... Peripheral and central chemoreceptors provide inputs to the respiratory centres. The peripheral chemoreceptors lie in the carotid and aortic bodies. The carotid bodies are more important in stimulating ventilation, while the aortic bodies are also capable of responding to hypotension. The peripheral ...

Respiratory Basics
... The lungs also requires a supply of oxygenated arterial blood to sustain the lung tissues. This is provided by bronchial arteries that branch off of either the aorta or the intercostal arteries. This is called the bronchial circulation. Normally, the bronchial circulation accounts for about 2% of ca ...
... The lungs also requires a supply of oxygenated arterial blood to sustain the lung tissues. This is provided by bronchial arteries that branch off of either the aorta or the intercostal arteries. This is called the bronchial circulation. Normally, the bronchial circulation accounts for about 2% of ca ...

- Orange Coast College
... Decreases ability of bronchioles to remain open during expiration. Cigarette smoking stimulates macrophages and leukocytes to secrete protein digesting enzymes that destroy tissue. ...
... Decreases ability of bronchioles to remain open during expiration. Cigarette smoking stimulates macrophages and leukocytes to secrete protein digesting enzymes that destroy tissue. ...

5-Endo thyroid
... • Describe the regulation of thyroid hormones secretion. • Know thyroid function tests. • Identify thyroid hormone receptors. • Describe thyroid hormone resistance. ...
... • Describe the regulation of thyroid hormones secretion. • Know thyroid function tests. • Identify thyroid hormone receptors. • Describe thyroid hormone resistance. ...

RAAS Basic physiology and Pathophysiolgy
... ACE/Ang II/AT1R and the new ACE2/Ang 1–7/Mas1R pathway Imbalance in classical ACE-Ang II and ACE2-Ang 1–7 is Critical in CVD development. ACE2 is regarded as the central regulator of the RAAS. As its changes can affect not only Ang II detrimental actions but also Ang 1–7 protective effects. ...
... ACE/Ang II/AT1R and the new ACE2/Ang 1–7/Mas1R pathway Imbalance in classical ACE-Ang II and ACE2-Ang 1–7 is Critical in CVD development. ACE2 is regarded as the central regulator of the RAAS. As its changes can affect not only Ang II detrimental actions but also Ang 1–7 protective effects. ...

Insufficiency of digestion
... also raise output of Са++ due to action on IP3 . Parietal cell has a receptor for prostaglandin E2 (PGE2)) which stimulation reduces a level cAMP and results in braking secretion of hydrochloric acid. • Secretion of a hydrochloric acid by parietal cell is carried out by a principle of the proton pum ...
... also raise output of Са++ due to action on IP3 . Parietal cell has a receptor for prostaglandin E2 (PGE2)) which stimulation reduces a level cAMP and results in braking secretion of hydrochloric acid. • Secretion of a hydrochloric acid by parietal cell is carried out by a principle of the proton pum ...

Acid – Base Physiology
... Renal Tubular Acidosis: due to a defect in H+ secretion or HCO3 reabsroption. Diarrhea: Excess bicarbonate loss into the feces without time to reabsorb (most common cause). Diabetes mellitus: In the absence of normal glucose metabolism the cells metabolize fats and form acetoacetic acid, reducing pH ...
... Renal Tubular Acidosis: due to a defect in H+ secretion or HCO3 reabsroption. Diarrhea: Excess bicarbonate loss into the feces without time to reabsorb (most common cause). Diabetes mellitus: In the absence of normal glucose metabolism the cells metabolize fats and form acetoacetic acid, reducing pH ...

Respiratory physiology - Assets - Cambridge
... for this response. Overall the peripheral chemoreceptors are only responsible for 20% of the body’s response to a raised PaCO2 , with the rest being due to the central chemoreceptors; however, it is the peripheral chemoreceptors that act fastest. Other receptors that feed information to the respirat ...
... for this response. Overall the peripheral chemoreceptors are only responsible for 20% of the body’s response to a raised PaCO2 , with the rest being due to the central chemoreceptors; however, it is the peripheral chemoreceptors that act fastest. Other receptors that feed information to the respirat ...
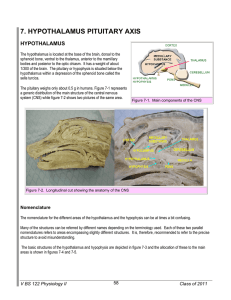
7. HYPOTHALAMUS PITUITARY AXIS
... hypothalamic centres produce a variety of inhibitory and stimulatory factors, which are released into a specific microcirculatory system called the portal system. This network of vessels starts in the base of the hypothalamus and extends into the pars distalis of the hypophysis where it irrigates al ...
... hypothalamic centres produce a variety of inhibitory and stimulatory factors, which are released into a specific microcirculatory system called the portal system. This network of vessels starts in the base of the hypothalamus and extends into the pars distalis of the hypophysis where it irrigates al ...

1 FORM W KEY deducted if you fail to do this!!!!!!
... 22. On planet Tatooine the Hutts evolved a urinary system very similar to humans. Fig 2 shows the collecting duct passing through the gradient set up by the Loop of Henle. What is the most concentrated urine the Hutts can produce? The most dilute urine they can produce? a) 1000; 100 b) 1400; 300 c) ...
... 22. On planet Tatooine the Hutts evolved a urinary system very similar to humans. Fig 2 shows the collecting duct passing through the gradient set up by the Loop of Henle. What is the most concentrated urine the Hutts can produce? The most dilute urine they can produce? a) 1000; 100 b) 1400; 300 c) ...

Acid – Base Physiology
... Renal Tubular Acidosis: due to a defect in H+ secretion or HCO3- reabsroption. Diarrhea: Excess HCO3- loss into the feces without time to reabsorb (most common cause). Diabetes mellitus: In the absence of normal glucose metabolism the cells metabolize fats and form acetoacetic acid, reducing pH, and ...
... Renal Tubular Acidosis: due to a defect in H+ secretion or HCO3- reabsroption. Diarrhea: Excess HCO3- loss into the feces without time to reabsorb (most common cause). Diabetes mellitus: In the absence of normal glucose metabolism the cells metabolize fats and form acetoacetic acid, reducing pH, and ...

BHS Anatomy and Physiology
... The lymph nodes at the junctions of the extremities and trunk destroy the pathogens before they reach the main part of the body and the vital organs. THE BLACK DEATH In 1347, several Italian merchant ships returned to Messina on the Mediterranean island of Sicily from a trip to the Black Sea. As the ...
... The lymph nodes at the junctions of the extremities and trunk destroy the pathogens before they reach the main part of the body and the vital organs. THE BLACK DEATH In 1347, several Italian merchant ships returned to Messina on the Mediterranean island of Sicily from a trip to the Black Sea. As the ...


Diffusion, Blood O2, CO2 Content and Transport
... the blood PCO2/pH. The Haldane effect on the other hand (or the other Hb hand) states that the CO2 content in blood is affected by the amount of oxygen dissolved in blood or the PO2. Carbon dioxide physiologically is more than merely a metabolic waste product. CO2 is a major determinant of blood pH, ...
... the blood PCO2/pH. The Haldane effect on the other hand (or the other Hb hand) states that the CO2 content in blood is affected by the amount of oxygen dissolved in blood or the PO2. Carbon dioxide physiologically is more than merely a metabolic waste product. CO2 is a major determinant of blood pH, ...
Neurotransmitter Flashcards
... They cause over-stimulation of nearby neurons, which then release too much calcium, causing cytotoxicity. It can kill the cell. 10% of the time it is genetic. 90% of the time it is caused by calcium dyshomeostasis (The calcium is not being monitored properly in the body). Glutamate Antagonist ...
... They cause over-stimulation of nearby neurons, which then release too much calcium, causing cytotoxicity. It can kill the cell. 10% of the time it is genetic. 90% of the time it is caused by calcium dyshomeostasis (The calcium is not being monitored properly in the body). Glutamate Antagonist ...

Respiratory and Renal Review
... overcome by surfactant - reduces the surface tension of the aqueous air-water interface proportional to the area it covers. ! surface tension of an alveolus decreases as the alveolus gets smaller and pressure does not increase as it shrinks (! alveolar capillary filtration forces $). • Interdependen ...
... overcome by surfactant - reduces the surface tension of the aqueous air-water interface proportional to the area it covers. ! surface tension of an alveolus decreases as the alveolus gets smaller and pressure does not increase as it shrinks (! alveolar capillary filtration forces $). • Interdependen ...

Relation of the Lungs to the Thoracic (Chest) Wall
... must focus, therefore, on how the changes in lung dimensions are brought about. The lungs are passive elastic structures—like balloons—and their volume, therefore, depends upon: (1) The difference in pressure between the inside the (Lung alveoli Palv) and the outside of the lungs (intra pleural Pip) ...
... must focus, therefore, on how the changes in lung dimensions are brought about. The lungs are passive elastic structures—like balloons—and their volume, therefore, depends upon: (1) The difference in pressure between the inside the (Lung alveoli Palv) and the outside of the lungs (intra pleural Pip) ...
Neurotransmitter Flashcards
... They cause over-stimulation of nearby neurons, which then release too much calcium, causing cytotoxicity. It can kill the cell. 10% of the time it is genetic. 90% of the time it is caused by calcium dyshomeostasis (The calcium is not being monitored properly in the body). Glutamate Antagonist ...
... They cause over-stimulation of nearby neurons, which then release too much calcium, causing cytotoxicity. It can kill the cell. 10% of the time it is genetic. 90% of the time it is caused by calcium dyshomeostasis (The calcium is not being monitored properly in the body). Glutamate Antagonist ...

Dear Notetaker:
... enough O2 to function and fight off infection 4) Exercise Effect o ↓ pH (due to lactic acid build up) o ↑ pCO2 (at tissue) o ↑ temperature (due to ATP used during muscle contraction) o Results in a right shift of the dissociate curve, resulting in an increase in O2 delivery ...
... enough O2 to function and fight off infection 4) Exercise Effect o ↓ pH (due to lactic acid build up) o ↑ pCO2 (at tissue) o ↑ temperature (due to ATP used during muscle contraction) o Results in a right shift of the dissociate curve, resulting in an increase in O2 delivery ...
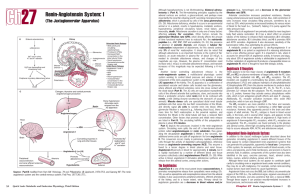
Renin-Angiotensin System: I
... Another factor controlling aldosterone release is the renin–angiotensin system, a multifactorial physiologic control system working to control blood pressure and volume. A major component of the renin–angiotensin system is the juxtaglomerular (JG) apparatus of the kidney. The JG apparatus is a combi ...
... Another factor controlling aldosterone release is the renin–angiotensin system, a multifactorial physiologic control system working to control blood pressure and volume. A major component of the renin–angiotensin system is the juxtaglomerular (JG) apparatus of the kidney. The JG apparatus is a combi ...

Cell Membranes - University of Nevada, Las Vegas
... Can include or exclude various proteins Are concentrating platforms for cell-signaling molecules ...
... Can include or exclude various proteins Are concentrating platforms for cell-signaling molecules ...

Respiratory - GEOCITIES.ws
... hemorrhage (under such high pressures capillaries will rupture – can control will drugs ie furosamide/lasix) Fluid tends to accumulate in lungs: due to mechanical forces, fluid moves from capillaries to interstitial space Uptake of interstitial fluid by lymphatics critical to keep alveoli free of ex ...
... hemorrhage (under such high pressures capillaries will rupture – can control will drugs ie furosamide/lasix) Fluid tends to accumulate in lungs: due to mechanical forces, fluid moves from capillaries to interstitial space Uptake of interstitial fluid by lymphatics critical to keep alveoli free of ex ...

Physiology Student Objectives Enduring understanding 2.C
... ● Plant defenses against pathogens include molecular recognition systems with systemic responses; infection triggers chemical responses that destroy infected and adjacent cells, thus localizing the effects. ● Vertebrate immune systems have nonspecific and nonheritable defense mechanisms against path ...
... ● Plant defenses against pathogens include molecular recognition systems with systemic responses; infection triggers chemical responses that destroy infected and adjacent cells, thus localizing the effects. ● Vertebrate immune systems have nonspecific and nonheritable defense mechanisms against path ...

hormon
... Regardless of their chemical nature hormones are present in the blood stream in very low concentration 10-7 – 10-12 M They bind to a very specific, high-affinity cellular receptors located at CELL MEMBRANES or in the NUCLEUS. ...
... Regardless of their chemical nature hormones are present in the blood stream in very low concentration 10-7 – 10-12 M They bind to a very specific, high-affinity cellular receptors located at CELL MEMBRANES or in the NUCLEUS. ...
Alveolar macrophage

An alveolar macrophage (or dust cell) is a type of macrophage found in the pulmonary alveolus, near the pneumocytes, but separated from the wall.Activity of the alveolar macrophage is relatively high, because they are located at one of the major boundaries between the body and the outside world.Dust cells are another name for monocyte derivatives in the lungs that reside on respiratory surfaces and clean off particles such as dust or microorganisms.Alveolar macrophages are frequently seen to contain granules of exogenous material such as particulate carbon that they have picked up from respiratory surfaces. Such black granules may be especially common in smoker's lungs or long-term city dwellers.Inhaled air may contain particles or organisms which would be pathogenic. The respiratory pathway is a prime site for exposure to pathogens and toxic substances. The respiratory tree, comprising the larynx, trachea, and bronchioles, is lined by ciliated epithelia cells that are continually exposed to harmful matter. When these offensive agents infiltrate the superficial barriers, the body's immune system responds in an orchestrated defense involving a litany of specialized cells which target the threat, neutralize it, and clean up the remnants of the battle. Deep within the lungs exists its constituent alveoli sacs, the sites responsible for the uptake of oxygen and excretion of carbon dioxide. There are three major alveolar cell types in the alveolar wall (pneumocytes): Type I pneumocyte (Squamous Alveolar) cells that form the structure of an alveolar wall. Type II pneumocyte (Great Alveolar) cells that secrete pulmonary surfactant to lower the surface tension of water and allows the membrane to separate, thereby increasing the capability to exchange gases. Surfactant is continuously released by exocytosis. It forms an underlying aqueous protein-containing hypophase and an overlying phospholipid film composed primarily of dipalmitoyl phosphatidylcholine. Macrophages that destroy foreign material, such as bacteria.Type 1 and type 2 pneumocytes. Type 1 pneumocytes (or membranous pneumocytes) form the structure of the alveolus and are responsible for the gas exchange in the alveolus. Type 1 pneumocytes are squamous epithelial cells which are characterized by a superficial layer consisting of large, thin, scale-like cells; they also cover 95% of the alveolar surface, although they are only half as numerous as Type 2 pneumocytes. Type 2 pneumocytes are important in that they can proliferate and differentiate into type 1 pneumocytes, which cannot replicate and are susceptible to a vast numbers of toxic insults. Type 2 pneumocytes are also important because they secrete pulmonary surfactant(PS), which consists 80-90% of phospholipids [(phosophatidylcholine(PC), phosphatidyglycerol(PG), phosphaditylinositol (PI)] and 5-10% of surfactant proteins (SP-A, SP-B, SP-C, AND SP-D). PS is synthesized as lamellar bodies, which are structures consisting of closely packed bilayers that are secreted and then undergo transformation into a morphological form called tubular myelin. PS plays an important role in maintaining normal respiratory mechanics by reducing alveolar surface tension. By lowering alveolar surface tension, PS reduces the energy required to inflate the lungs, and reduces the likelihood of alveolar collapse during expiration. Loosely attached to these alveoli sacs are the alveolar macrophages that protect the lungs from a broad array of microbes and aerosols by devouring and ingesting them through phagocytosis.Alveolar macrophages are phagocytes that play a critical role in homeostasis, host defense, the response to foreign substances, and tissue remodeling. Since alveolar macrophages are pivotal regulators of local immunological homeostasis, their population density is decisive for the many processes of immunity in the lungs. They are highly adaptive components of the innate immune system and can be specifically modified to whatever functions needed depending on their state of differentiation and micro-environmental factors encountered. Alveolar macrophages release numerous secretory products and interact with other cells and molecules through the expression of several surface receptors. Alveolar macrophages are also involved in the phagocytosis of apoptotic and necrotic cells that have undergone cell-death. They must be selective of the material that is phagocytized because normal cells and structures of the body must not be compromised. To combat infection, the phagocytes of the innate immune system facilitates many pattern recognition receptors (PRR) to help recognize pathogen-associated molecular patterns (PAMPs) on the surface of pathogenic microorganisms. PAMPs all have the common features of being unique to a group of pathogens but invariant in their basic structure; and are essential for pathogenicity(ability of an organism to produce an infectious disease in another organism). Proteins involved in microbial pattern recognition include mannose receptor, complement receptors, DC-SIGN,Toll-like receptors(TLRs), the scavenger receptor, CD14, and Mac-1. PRRs can be divided into three classes:signaling PRRs that activate gene transcriptional mechanisms that lead to cellular activation,endocytic PRRs that function in pathogen binding and phagocytosis, andsecreted PRRs that usually function as opsonins or activators of complement.The recognition and clearance of invading microorganisms occurs through both opsonin-dependent and opsonin–independent pathways. The molecular mechanisms facilitating opsonin-dependent phagocytosis are different for specific opsonin/receptor pairs. For example, phagocytosis of IgG-opsonized pathogens occurs through the Fcγ receptors (FcγR), and involves phagocyte extensions around the microbe, resulting in the production of pro-inflammatory mediators. Conversely, complement receptor-mediated pathogen ingestion occurs without observable membrane extensions (particles just sink into the cell) and does not generally results in an inflammatory mediator response. Following internalization, the microbe is enclosed in a vesicular phagosome which then undergoes fusion with primary or secondary lysosomes, forming a phagolysosome. There are various mechanisms that lead to intracellular killing; there are oxidative processes, and others independent of the oxidative metabolism. The former involves the activation of membrane enzyme systems that lead to a stimulation of oxygen uptake (known as the respiratory burst), and its reduction to reactive oxygen intermediates (ROIs), molecular species that are highly toxic for microorganisms. The enzyme responsible for the elicitation of the respiratory burst is known as nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, which is composed of five subunits. One component is a membrane cytochrome made up of two protein subunits, gp91phox and p22phox; the remaining three components are cytosolic-derived proteins: p40phox, p47phox, and p67phox. NADPH oxidase exists in the cytosol of the AM when in a quiescent state; but upon activation, two of its cytosolic components, p47phox and p67phox, have their tyrosine and serine residues phosphorylated, which are then able to mediate translocation of NADPHox to the cytochrome component, gp91phox/p22phox, on the plasma membrane via cytoskeletal elements.Compared to other phagocytes, the respiratory burst in AM is of a greater magnitude. Oxygen-independent microbicidal mechanisms are based on the production of acid, on the secretion of lysozymes, on iron-binding proteins, and on the synthesis of toxic cationic polypeptides. Macrophages possess a repertoire of antimicrobial molecules packaged within their granules and lysosomes. These organelles contain a plethora of degradative enzymes and antimicrobial peptides that are released into the phagolysosome, such as proteases, nucleases, phosphatases, esterases, lipases, and highly basic peptides. Moreover, macrophages possess a number of nutrient deprivation mechanisms that are used to starve phagocytosed pathogens of essential micronutrients. Certain microorganisms have evolved countermeasures which enable them to evade being destroyed by phagocytes. Although lysosomal-mediated degradation is an efficient means by which to neutralize an infection and prevent colonization, several pathogens parasitize macrophages, exploiting them as a host cell for growth, maintenance and replication. Parasites like Toxoplasma gondii and mycobacteria are able to prevent fusion of phagosomes with lysosomes, thus escaping the harmful action of lysosomal hydrolases. Others avoid lysosomes by leaving the phagocytic vacuole, to reach the cytosolic matrix where their development is unhindered. In these instances, macrophages may be triggered to actively destroy phagocytosed microorganisms by producing a number of highly toxic molecules and inducing deprivational mechanism to starve it. Finally, some microbes have enzymes to detoxify oxygen metabolites formed during the respiratory burst.When insufficient to ward off the threat, alveolar macrophages can release proinflammatory cytokines and chemokines to call forth a highly developed network of defensive phagocytic cells responsible for the adaptive immune response. The lungs are especially sensitive and prone to damage, thus to avoid collateral damage to type 1 and type II pneumocytes, alveolar macrophages are kept in a quiescent state, producing little inflammatory cytokines and displaying little phagocytic activity, as evidenced by downregulated expression of the phagocytic receptor Macrophage 1 antigen (Mac-1). AMs actively suppress the induction of two of the immunity systems of the body: the adaptive immunity and humoral immunity. The adaptive immunity is suppressed through AM’s effects on interstitial dendritic cells, B-cells and T-cells, as these cells are less selective of what they destroy, and often cause unnecessary damage to normal cells. To prevent uncontrolled inflammation in the lower respiratory tract, alveolar macrophages secrete nitric oxide, prostaglandins, interleukin-4 and -10(IL-4, IL-10), and transforming growth factor-β (TGF-β).