Survey
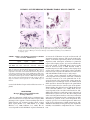
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
DEVELOPMENTAL GENETICS 25:224–236 (1999) Ectopic Expression of AINTEGUMENTA in Arabidopsis Plants Results in Increased Growth of Floral Organs BETH ALLYN KRIZEK* Department of Biological Sciences, University of South Carolina, Columbia, South Carolina ABSTRACT AINTEGUMENTA (ANT) was previously shown to be involved in floral organ initiation and growth in Arabidopsis. ant flowers have fewer and smaller floral organs and possess ovules that lack integuments and a functional embryo sac. The present work shows that young floral meristems of ant plants are smaller than those in wild type. Failure to initiate the full number of organ primordia in ant flowers may result from insufficient numbers of meristematic cells. The decreased size of ant floral organs appears to be a consequence of decreased cell division within organ primordia. Ectopic expression of ANT under the control of the constitutive 35S promoter results in the development of larger floral organs. The number and shape of these organs is not altered and the size of vegetative organs is normal. Microscopic and molecular analyses indicate that the increased size of 35S::ANT sepals is the result of increased cell division, whereas the increased sizes of 35S::ANT petals, stamens, and carpels are primarily attributable to increased cell expansion. In addition, 35S::ANT ovules often exhibit increased growth of the nucellus and the funiculus. These results suggest that ANT stimulates cell growth in floral organs. Dev. Genet. 25:224–236, 1999. r 1999 Wiley-Liss, Inc. Key words: Arabidopsis flower development; AINTEGUMENTA; cell division; cell expansion; AP2/EREBP family INTRODUCTION Flowers are derived from groups of undifferentiated cells called floral meristems. Floral organ primordia arise at defined positions from within these meristems, grow, and eventually differentiate into the four organs of a flower (sepals, petals, stamens, and carpels). As plant cells do not undergo migration, cell division and cell expansion are the predominant mechanisms by which the number and position of organ primordia is determined. In addition, the final size and shape of each organ is also controlled by these processes. The number, location, and plane of each cell division in the developing organ primordia, as well as the amount and direction of cell expansion, are critically important in deter- r 1999 WILEY-LISS, INC. mining the final form of each organ. Although certain aspects of flower development, such as the establishment of floral organ identity, are well characterized [reviewed in Coen and Meyerowitz, 1991; Ma, 1994; Sessions et al., 1998; Weigel and Meyerowitz, 1994], very little is known about how the patterns and numbers of cell divisions are controlled in flowers during organ initiation and organ growth [Meyerowitz, 1997]. Furthermore, little is known about the control of cell expansion during plant development. Cell expansion is known to be regulated by phytohormones such as auxin, gibberellin, and brassinosteroids [reviewed in Cleland, 1987; Hooley, 1996; Métraux, 1987]; mutants in the biosynthesis or signal transduction pathways of these hormones often exhibit defects in cell expansion. Recently, additional genes with roles in controlling cell expansion have been identified [Hanzawa et al., 1997; Kim et al., 1998; Sablowski and Meyerowitz, 1998; Wilson et al., 1996]. Whereas some genes seem to play general roles in cell expansion in all tissues [Takahashi et al., 1995], others appear to have specific functions in particular organs [Sablowski and Meyerowitz, 1998] or at particular times in development [Hanzawa et al., 1997], suggesting that cell expansion is controlled by different factors in different tissues. One gene that is involved in the control of organ growth during Arabidopsis flower development is AINTEGUMENTA (ANT). Mutations in ant result in a random reduction in floral organ number, the production of narrow floral organs, and defects in ovule development including the absence of integuments and a female gametophyte [Baker et al., 1997; Elliott et al., 1996; Klucher et al., 1996; Schneitz et al., 1997]. These defects in both organ initiation and organ growth suggest that ANT may be involved in regulating cell Contract grant sponsor: Department of Energy; Contract grant number: 98ER20312. *Correspondence to: Beth Allyn Krizek, Department of Biological Sciences, University of South Carolina, Columbia, SC 29208. E-mail: [email protected] Received 3 March 1999; Accepted 1 June 1999 ECTOPIC ANT EXPRESSION INCREASES FLORAL ORGAN GROWTH division in flowers. ANT is a member of the AP2/EREBP family of transcription factors, containing two AP2 domains of approximately 70 amino acids [Elliott et al., 1996; Klucher et al., 1996]. These domains have been shown to bind DNA in other members of the AP2/ EREBP family [Buttner and Singh, 1997; Kagaya et al., 1999; Liu et al., 1998; Ohme-Takagi and Shinshi, 1995; Stockinger et al., 1997; Zhou et al., 1997]. In addition, ANT has been shown to function as a transcription factor in yeast [Vergani et al., 1997]. ANT is expressed in cotyledon, leaf, floral organ, and ovule primordia [Elliott et al., 1996; Klucher et al., 1996]. In flowers, ANT RNA is initially detected throughout organ primordia but later becomes restricted to particular subdomains within developing organs [Elliott et al., 1996]. In particular, these domains of ANT expression seem to correlate with regions undergoing active growth [Elliott et al., 1996]. Ovule primordia consist of several morphological regions: the base or stalk of the primordia (funiculus) that connects the ovule to the maternal tissue, a central or chalazal region from which two integuments arise and eventually develop into the seed coat, and the apical tip of the primordia (the nucellus) in which the megaspore mother cell is produced [reviewed in Reiser and Fischer, 1993]. In ovules, ANT is initially expressed throughout the primordia and later becomes restricted primarily to the chalazal region of the ovule before integument initiation [Elliott et al., 1996]. Expression continues in the integuments during their early development, decreases as the outer integument grows to cover the nucellus, and eventually becomes limited to the interiormost cell layer of the inner integument [Elliott et al., 1996]. To investigate further the role of ANT in floral organ initiation and growth, ANT was ectopically expressed in wild-type Arabidopsis plants under the constitutive cauliflower mosaic virus 35S promoter (35S::ANT). The most dramatic phenotype exhibited by 35S::ANT plants is the production of larger floral organs. The increased size of these organs results from an increase in cell number in the case of sepals and appears to be largely due to an increase in cell size in petals, stamens, and carpels. The relative shape of these organs is maintained, suggesting that organ size is controlled independently from organ shape. In addition, 35S::ANT ovules often exhibit increased growth of the nucellus and funiculus but decreased growth of the outer integument. Characterization of ant flowers by two photon fluorescence microscopy shows that ant stage 3 floral meristems are smaller than wild-type meristems of similar age. Such data provide a possible explanation for the decreased numbers of organs initiated in ant mutants. The smaller size of ant floral organs results from decreased cell division within floral organ primordia. Both the ant mutant and 35S::ANT phenotypes can be explained by a model in which ANT stimulates cell growth. 225 MATERIALS AND METHODS Production of 35S::ANT Plants ANT cDNA was PCR amplified using Pfu polymerase (Stratagene) with primers containing BamHI and XbaI restriction sites at the 58 and 38 ends of the cDNA, respectively. The polymerase chain reaction (PCR) product was originally cloned into pLITMUS28 (New England Biolabs), and its sequence was verified by doublestranded sequencing of the recombinant plasmid. ANT was subsequently subcloned into pGEM3Z containing the 35S promoter in the KpnI/BamHI sites. 35S::ANT was cut out of this plasmid with KpnI/XbaI and subcloned into the plant transformation vector pCGN1547, which contained a 38 NOS sequence [Krizek and Meyerowitz, 1996]. 35S::ANT/pCGN-NOS was transformed into Agrobacterium ASE by electroporation and subsequently transformed into L-er, ant ⫺6/⫹, and ant ⫺8/⫹ plants using the in planta vacuum infiltration procedure [Bechtold et al., 1993]. Transformants were selected by germination of the seeds on MS media containing kanamycin. Putative 35S::ANT ant ⫺6 and 35S:: ANT ant ⫺8 plants were genotyped as described later in Materials and Methods. Organ Length, Cell Length, and Cell Area Measurements Floral organ lengths were measured using an ocular micrometer. Two adjacent sepals, two adjacent petals, and two lateral stamens were removed from stage 14 flowers (flowers staged as described in Smyth et al. [1990] and Müller [1961]). The length of the carpel was measured after the remaining floral organs were removed from each flower. The average size of petal blade cells from L-er and 35S::ANT flowers was determined using the software IPLab (Scanalytics, Fairfax, VA). Scanning electron micrograph images were segmented into individual cells for quantitation by either hand drawing around the cell or using intensity thresholding. The areas of segments corresponding to individual petal cells were then calculated by IPLab. The average length of anther epidermal cells also was determined using IPLab. Lines were drawn the middle of each of 13 cells from scanning electron micrographs of L-er and 35S::ANT anthers. The lengths of these lines were then calculated by IPLab. For the petal and stamen IPLab measurements, four to five SEM image files from four to five different petals or stamens, respectively, were used. The images used in these measurements corresponded to similar parts of the respective floral organs in the L-er and 35S::ANT flowers. Scanning Electron Microscopy and Two Photon Fluorescence Microscopy Samples for SEM were fixed and dried as described previously [Bowman et al., 1991]. For viewing ovules, carpels were sliced with a razor blade immediately before fixation. Flowers and floral organs were mounted 226 KRIZEK onto stubs, dissected with glass needles as necessary, and coated with gold in a Denton Desk II gold sputter/ etch unit. Images were collected on a Hitachi S-2500D scanning electron microscope and digitally saved using the software Iridium (IXRF Systems, Houston, TX). Inflorescence tissue from L-er, 35S::ANT, and ant ⫺5 plants was prepared for two photon microscopy as described previously for confocal microscopy [Running et al., 1995], except that in some cases, the tissue was treated with RNase at 50 mg/ml for 30 min at 37°C after fixation and before staining in propidium iodide. This step reduced staining in the cytoplasm. The data were collected on a Bio-Rad system equipped with a titaniumsapphire laser using a ⫻60 oil immersion lens. Propidium iodide was excited at 770 nm, and emitted light was collected after passage through filters absorbing wavelengths shorter than 575 nm. Sequencing and Genotyping of the ant ⴚ6 and ant ⴚ8 Alleles DNA from ant ⫺6 and ant ⫺8 plants [Baker et al., 1997] was isolated using standard methods. ant was PCR amplified from each of these DNA samples in five overlapping pieces using either Pfu (Stratagene) or vent (New England Biolabs) polymerase. These PCR products were then sequenced directly on a ABI 377 automated sequencer. The ant ⫺6 mutation C679=T results in a nonsense mutation (Gln227=stop codon). An exon 2 primer was engineered to create an MseI site specifically in the ant ⫺6 allele. PCR was performed on leaf tissue [Klimyuk et al., 1993] removed from putative 35S::ANT ant ⫺6 plants using this primer and an intron 2 primer. The ant ⫺8 mutation (G1267=A, which results in an Ala =Thr missense mutation) disrupts a PstI site. An intron 7-specific primer and exon 8 primer were used to PCR-amplify this region from candidate 35S::ANT ant ⫺8 plants. Restriction enzyme analyses distinguished plants that were homozygous wild-type, homozygous ant ⫺6 (or ant ⫺8), or heterozygous. In Situ Hybridization Flowers for radioactive ANT in situs and nonradioactive histone H4 in situs were fixed, embedded, sectioned, and hybridized as described previously [Sakai et al., 1995]. The radioactive ANT in situ slides were washed as described previously [Sakai et al., 1995]. To make the ANT antisense probe, a BamHI/ClaI fragment of ANT cDNA (corresponding to the 58 half of the gene, not including the AP2 repeats) was cloned into the HincII site of pGEM3Z (pANTsitu). pANTsitu was linearized by digestion with EcoRI and in vitro transcribed in the presence of [35S]UTP with SP6 RNA polymerase. Sections were exposed for 1–2 weeks. Flowers for the nonradioactive histone H4 in situs were washed as previously described [Coen et al., 1990]. Immunological detection of the hybridized probe was performed by blocking with 1% Boehringer blocking agent in phosphate-buffered saline (PBS) with 0.3% Triton X1000 for 1 h, blocking with 0.5% bovine serum albumin (BSA) in PBS with 0.3% Triton X1000 for 1 h, incubation in anti-DIG antibody diluted 1:1,000 in 0.5% BSA/PBS/0.3% Triton X1000 for 5 h, washing with PBS and subsequently 100 mM Tris pH9.5, 10 mM NaCl, 50 mM MgCl2, and color development with NBT/X-phos for approximately 12 h. Histone H4 antisense probe was made by linearization of pHS-H4 with SpeI and in vitro transcription with T7 RNA polymerase in the presence of digoxigenin-11-UTP (Boehringer-Mannheim). Histone H4 sense probe was made by linearization of pHS-H4 with NotI and in vitro transcription with SP6 RNA polymerase in the presence of digoxigenin-11-UTP. The number of cells expressing histone H4 in sepals and petals (of stage 7 and 10 flowers) was determined in the following manner. Adjacent tissue sections of appropriately staged flowers that contained a full-length longitudinal section of a sepal or petal were identified, the number of stained cells in each tissue section counted, and an average for that individual organ determined. An overall average was calculated from these individual organ averages. RESULTS Ectopic Expression of ANT Increases Floral Organ Size After initial expression throughout floral organ primordia, ANT RNA becomes limited to particular subdomains as the organs mature [Elliott et al., 1996] (Fig. 1A–D). To investigate the consequences of changing the levels and/or duration of ANT expression in developing organ primordia, ANT cDNA was fused to the constitutive 35S promoter from cauliflower mosaic virus and this construct was transformed into wild-type Arabidopsis plants. As shown in Figure 1E–H, 35S::ANT confers persistent ANT expression in developing floral organs. Approximately one-half of the transgenic plants (66 out of 127 plants) produced larger than normal flowers (Fig. 2A). Of the other transformants, most were wild-type in appearance (58), while three had phenotypes resembling ant mutants. The plants with ant phenotypes may arise from cosuppression [reviewed in Depicker and van Montagu, 1997]. Further characterization was performed on those plants which produced the larger flower phenotype. Because these plants are sterile (described later), all phenotypic characterizations were done on primary transformants. DNA gel blot analysis indicated that some of the 35S::ANT lines with the larger flower phenotype contained a single copy of the transgene, whereas others had multiple copies. The increased size of these 35S::ANT flowers is attributable to increases in the size of all four types of floral organs. Floral organ number is unchanged in 35S::ANT plants. The differences in organ size were quantitated by measuring the lengths of floral organs removed from L-er and 35S::ANT stage 14 flowers ECTOPIC ANT EXPRESSION INCREASES FLORAL ORGAN GROWTH 227 Fig. 1. Expression pattern of ANT in L-er and 35S::ANT plants. In situ hybridization of an ANT antisense RNA probe with longitudinal sections through wild-type (L-er) (A–D) and 35S::ANT tissue (E–H). Each section was photographed in brightfield (A,C,E,G) or darkfield (B,D,F,H). A,B: ANT RNA is detected throughout young floral primor- dia (arrow). Expression decreases in older flowers. C,D: ANT is no longer expressed in the sepals or anthers of this stage 10 flower. E,F: ANT RNA is detected throughout the inflorescence of 35S::ANT plants. G,H: ANT RNA is detected throughout this stage 9 35S::ANT flower. se, sepal; pe, petal; st, stamen; ca, carpel. (flowers staged as described in Smyth et al. [1990] and Müller [1961]), using a optical micrometer. These data were collected from the first 10 flowers on L-er and 35S::ANT plants that were grown side by side under equivalent conditions. Results from two different experiments are shown in Table 1. Organ size is increased from 11–34% in 35S::ANT flowers. The increased size of 35S::ANT floral organs is the opposite phenotype of that produced by mutations in ant which cause the development of smaller floral organs [Elliott et al., 1996; Klucher et al., 1996]. Although organ width was not measured, the overall shape of the larger 35S::ANT floral organs is normal (Fig. 2B). This is not true of mutations in ant, which tend to produce quite narrow floral organs [Baker et al., 1997; Elliott et al., 1996; Klucher et al., 1996] (Fig. 2B). The effects of ectopic ANT expression on organ growth appear to be restricted to floral organs as no differences were observed in the size of leaves on 35S::ANT plants compared with L-er plants or in the height of 35S::ANT plants compared with L-er plants (Fig. 2C). To confirm that ANT is expressed in these vegetative organs, in situ hybridization was performed on 35S::ANT leaf tissue sections. ANT RNA was detected throughout 35S::ANT leaf tissue (data not shown). layed in flowering compared with 35S::ANT plants with a wild-type appearance. Epicuticular wax that is normally present on ovary epidermal cells is not present on the surface of these cells in 35S::ANT flowers. 35S:: ANT plants are male sterile and show severe reductions in female fertility. The anthers do not dehisce; however, this is not the only cause of the male sterility phenotype. A defect in a late stage of pollen development appears to occur in 35S::ANT lines. Microspores with an exine wall are made and, in at least some cases, viable pollen grains are produced. Pollen viability was measured by staining with the dye fluorescein diacetate (FDA) [Regan and Moffatt, 1990], which assays for the integrity of the plasma membrane of the vegetative cell [Heslop-Harrison and Heslop-Harrison, 1970]. However, manual cutting of mature 35S::ANT anthers does not release individual dehydrated pollen grains as observed for wild-type anthers. Further work will be necessary to fully characterize pollen development in 35S::ANT flowers. When wild-type pollen is used to fertilize 35S::ANT carpels, a few seeds are occasionally produced. To investigate the basis for this reduced female fertility, the development of 35S::ANT ovules were characterized by scanning electron microscopy (SEM). The following discussion presents a brief description of wild-type ovule development [Robinson-Beers et al., 1992; Schneitz et al., 1995] using the stages assigned in Schneitz et al., 1995. Finger-like ovule primordia are initiated from the inner ovary walls during stage I of ovule development. During stage II, the inner integu- Other Floral Phenotypes Resulting From Ectopic Expression of ANT Several other effects on flower development were observed in 35S::ANT plants that exhibit the larger floral organ phenotype. These plants are slightly de- 228 KRIZEK Fig. 2. Phenotypes of 35S::ANT, 35S::ANT ant ⫺6, and 35S::ANT ant ⫺8 plants. A: Top down and side views of wild-type (L-er) and 35S::ANT flowers. B: Petals removed from L-er, 35S::ANT, and ant ⫺9 flowers. ant ⫺9 is a strong allele. C: L-er and 35S::ANT plants of equal age. The size of vegetative organs and the overall stature of these plants are the same. D: ant ⫺6 and 35S::ANT ant ⫺6 flowers showing the lack of complementation of the organ number and size defects of ant ⫺6. E: Scanning electron micrograph of a typical ant ⫺6 ovule. There is a slight expansion in the chalazal region of these ovules, where the inner and outer integuments initiate in wild-type ovules. F: Scanning electron micrograph of a 35S::ANT ant ⫺6 ovule showing increased growth of a single integument-like structure. G: Scanning electron micrograph of an ant ⫺8 ovule showing partial growth of a single integumentary-like structure around the nucellus. H: ant ⫺8 and 35S::ANT ant ⫺8 flowers showing complementation of the ant ⫺8 mutation by 35S::ANT. Scale bars ⫽ 10 µm (E–G). ment initiates as a symmetrical ring of cells in the middle or chalazal region of the ovule primordia. The outer integument initiates soon afterward as an asymmetrical extension of cells on the abaxial surface of the ovule just below the inner integument (Fig. 3A,B). During stages II and III, the inner integument expands symmetrically to cover the nucellus while the outer integument grows asymmetrically with more growth on the abaxial surface than the adaxial surface of the ovule. The outer integument eventually grows to cover the inner integument (Fig. 3C) and finally the nucellus (Fig. 3D). Differences in the development of 35S::ANT ovules are apparent shortly after the initiation of ovule primordia. Although two integuments are initiated in 35S:: ANT ovules, they are not as well defined as those in wild-type ovules (Fig. 3E,F). This is particularly true for the inner integument, which does not protrude significantly from the chalazal region of the ovule primordia (Fig. 3E). In 35S::ANT ovules, the outer integument appears to overtake the inner integument at a slightly earlier time than in wild-type ovules (cf. Fig. 3C,G). Unlike wild-type ovules, in which the outer integument completely surrounds the nucellus during ECTOPIC ANT EXPRESSION INCREASES FLORAL ORGAN GROWTH TABLE 1. Floral Organ Lengths (mm) in L-er and 35S⬋ANT Flowers* Sepals Petals Stamens Carpels L-er 1.86 ⫾ 0.08 3.15 ⫾ 0.21 2.51 ⫾ 0.16 2.49 ⫾ 0.21 35S⬋ANT 2.28 ⫾ 0.13 3.95 ⫾ 0.27 2.79 ⫾ 0.25 3.17 ⫾ 0.22 % increase 22.7% 25.2% 11.1% 27.6% L-er 1.88 ⫾ 0.14 3.11 ⫾ 0.38 2.65 ⫾ 0.21 2.54 ⫾ 0.31 35S⬋ANT 2.37 ⫾ 0.11 4.16 ⫾ 0.25 3.02 ⫾ 0.29 3.36 ⫾ 0.19 % increase 25.9% 34.0% 14.1% 32.0% *The data are indicated as averages ⫾ SD. The first data set is based on measurements on two sepals, two petals, two stamens, and one carpel from each of 28 L-er flowers on four plants and each of 45 35S⬋ANT flowers on 14 plants located at positions 1–10 on the inflorescence. The second data set is based on measurements on two sepals, two petals, two stamens, and one carpel from each of 29 L-er flowers on 4 plants and each of 47 35S⬋ANT flowers on eight plants located at positions 1–10 on the inflorescence. stage III, the outer integuments of 35S::ANT ovules rarely grow to surround the nucellus entirely (cf. Fig. 3D,H). A small number (approximately 10%) of mature 35S::ANT ovules are wild-type in appearance (cf. Fig. 3I and J), whereas the rest exhibit growth defects in the different ovule structures. In most 35S::ANT ovules, growth of the outer integument is prematurely terminated such that the nucellus is still visible (Fig. 3K). Some of these 35S::ANT ovules resemble ovules of the weak ant ⫺3 allele [Klucher et al., 1996]; however, in many cases, an unusually large nucellus protrudes from the reduced outer integument (Fig. 3L–N). Occasionally, ovules with a similar appearance were identified that had a micropyle-like hole visible in the tissue protruding from the outer integument (Fig. 3Q). This finding suggests that the protruding tissue is the inner integument; although analysis of the development of 35S::ANT ovules indicated that the inner integument stops growing before the outer integument. As this class of ovule was only observed rarely, it is possible that such a phenotype was missed during examination of early ovule development because of the smaller number of ovules examined. A few 35S::ANT ovules resemble sup ovules [Gaiser et al., 1995] in which there is increased growth of the outer integument on the adaxial surface of the ovule (Fig. 3H). In addition to the defects noted above, the funiculus of 35S::ANT ovules is often increased in length (Fig. 3N–P). This increased growth appears to primarily result from increased cell elongation (cf. Fig. 3I and P). 35S::ANT Sepals Contain More Cells, Whereas 35S::ANT Petals, Stamens, and Carpels Contain Larger Cells The increased size of 35S::ANT floral organs could result from an increase in cell number, an increase in cell size, or some combination of the two. SEM was used to characterize the size of sepal, petal, stamen, and 229 carpel epidermal cells in wild-type and 35S::ANT flowers. Comparison of cell size in sepals, petals, and carpels was performed on stage 14 flowers, at which time dehiscence of the anthers had already occurred. Stamen cell size was compared in stage 13 flowers in which the anthers had just begun to dehisce. Epidermal cells of petals, stamens, and carpels were found to be larger in 35S::ANT flowers compared with L-er flowers (cf. Fig. 4B–E and G–J). Thus, the increased size of petals, stamen, and carpels in 35S::ANT flowers is at least partially attributable to the presence of larger cells. No obvious difference in cell size was apparent in sepals (Fig. 4A,F), indicating that ectopic expression of ANT results in an increased number of cells in sepals. In general, cell shape and epidermal characteristics are conserved. To determine whether the increased size of cells in petals and stamens could account entirely for the increased size of these organs, scanning electron micrographs were analyzed with the graphics software IPLab. The cell area of both the adaxial and abaxial petal blade cells and the length of anther epidermal cells were determined (Table 2). The increase in the average cell area of adaxial and abaxial petal blade cells in 35S:: ANT flowers is approximately equal to the increase in petal size of 35S::ANT flowers, suggesting that increased cell expansion can account entirely for the increased size of these organs. An increase in the average length of 35S::ANT anther epidermal cells was similar to the overall increase in stamen length, suggesting that increased cell expansion may also account for the increased length of stamens in 35S::ANT flowers, although the relative size of cells in the stamen filament was not determined. Histone H4 Expression in L-er and 35S::ANT Flowers To investigate further whether the larger size of 35S::ANT sepals is due to increased cell division, the expression pattern of a cell division specific marker was examined in L-er and 35S::ANT sepals. Histone H4 has previously been shown to be expressed specifically during interphase in Antirrhinum floral meristem cells [Fobert et al., 1994] and predominantly during S phase of the cell cycle in many organisms [Marzluff and Pandey, 1988; Nakayama and Iwabuchi, 1993]. Overall, the pattern of histone H4 expression is similar in L-er and 35S::ANT flowers. In both L-er and 35S::ANT inflorescences and flowers, histone H4 is expressed in a spotty pattern, with each spot corresponding to an individual cell or a small group of cells (Fig. 5A–D). In sepals, petals, and stamens of older flowers (Fig. 5B,D), fewer cells are labeled, indicating decreased numbers of dividing cells or decreased rates of cell division, or both. These observations are similar to what has been reported previously for Antirrhinum [Fobert et al., 1994]. No signal was detected with a histone H4 sense probe. 230 KRIZEK Fig. 3. Scanning electron micrographs of ovule development in L-er and 35S::ANT flowers. Wild-type ovule development (A–D) and a mature wild-type ovule (I). 35S::ANT ovule development (E–H) and mature 35S::ANT ovules (J-P). Ovules have been staged according to the descriptions of Schneitz et al., 1995. A: Wild-type ovules at stage 2-IV. The inner integument and outer integument have initiated. B: Wild-type ovules at stage 2-V. Both integuments are growing around the nucellus. C: Stage 3-I wild-type ovules in which the outer integument is overtaking the inner integument. D: Stage 3-I wild-type ovules in which the outer integuments have enclosed the nucellus. E: 35S::ANT ovules in an early stage of development. Both the inner and outer integuments have initiated although they are not as well defined as in wild-type ovules. F: 35S::ANT ovules at stage 2-V, in which the outer integument is beginning to cover the inner integument. G: Stage 3-I 35S::ANT ovules in which the outer integument has completely enclosed the inner integument. H: Stage 3-I 35S::ANT ovules. In some cases, the nucellus continues to protrude from the outer integument. I: Mature wild-type ovule. J: 35S::ANT mature ovule that has a fairly normal morphology. K: 35S::ANT ovule in which the outer integument has not grown to fully enclose the nucellus. L: 35S::ANT ovule in which the outer integument does not fully enclose the nucellus and the nucellus has grown abnormally large. M: 35S::ANT ovules with a large nucellus protruding from the outer integument. N: 35S::ANT ovule with an enlarged nucellus and a long funiculus. O: 35S::ANT ovule in which the protruding tissue has an opening resembling a micropyle, suggesting that this is the inner integument. P: 35S::ANT ovule in which the outer integument shows increased growth on the adaxial side of the ovule, resembling sup ovules. ii, inner integument; oi, outer integument; n, nucellus; f, funiculus; m, micropyle. Scale bars ⫽ 10 µm. Fig. 4. Scanning electron micrographs of the epidermal cells on L-er and 35S::ANT floral organs. Epidermal cells from L-er organs are shown in A–E and corresponding epidermal cells (at the same magnification) from 35S::ANT organs are shown in F–J. A: L-er sepal epidermal cells. B: Epidermal cells on the adaxial surface of a L-er petal. C: Epidermal cells on the abaxial surface of a L-er petal. D: Anther epidermal cells from a L-er stamen. E: Ovary epidermal cells from a L-er carpel. F: 35S::ANT sepal epidermal cells. G: Epidermal cells on the adaxial surface of a 35S::ANT petal. H: Epidermal cells on the abaxial surface of a 35S::ANT petal. I: Anther epidermal cells from a 35S::ANT stamen. J: Ovary epidermal cells from a 35S::ANT carpel. Scale bars ⫽ 10 µm. 232 KRIZEK TABLE 2. Average Cell Size (Petals) and Average Cell Length (Stamens) in L-er and 35S⬋ANT Floral Organs* L-er 35S⬋ANT % increase Petals (adaxial) 1940 ⫾ 727 2458 ⫾ 746 26.7% Petals (abaxial) 1690 ⫾ 471 2245 ⫾ 496 32.8% Stamens 102 ⫾ 21 113 ⫾ 20 10.8% *The data are indicated as averages ⫾ SD. Measurement units are pixels. The adaxial petal data are based on measurements using 189 L-er and 259 35S⬋ANT cells from a total of four (L-er) or five petals (35S⬋ANT). The abaxial petal data are based on measurements using 132 L-er and 118 35S⬋ANT cells from a total of four petals each. The stamen data are based on measurements using 52 L-er and 52 35S⬋ANT cells from four stamens of each. The number of histone H4-expressing cells in sepals of L-er and 35S::ANT flowers of stages 6, 7, and 10 was counted (Table 3). Similar numbers of sepal cells expressing histone H4 were found in L-er and 35S::ANT stage 6 and 7 flowers. However, in sepals from stage 10 flowers, more than twice as many 35S::ANT cells expressed histone H4 as compared with L-er cells. These results are consistent with the SEM data providing further evidence that increased cell division accounts for the increased size of 35S::ANT sepals. These results do not indicate whether the rate of cell division in stage 10 35S::ANT sepals is faster than in wild-type sepals or whether the population of dividing cells is larger. For comparison, the number of L-er and 35S:: ANT petal cells in stage 10 flowers that expressed histone H4 was also determined. No significant differences were detected, consistent with earlier results indicating that increased cell expansion is primarily responsible for the increased size of 35S::ANT petals. Inflorescence and Floral Meristem Size in L-er, 35S::ANT, and ant ⴚ5 Plants To characterize further the effects of ectopic ANT expression on floral organ growth, two photon fluorescence microscopy was used to compare the size of young floral meristems and young floral organ primordia in L-er and 35S::ANT plants. Stage 3 floral meristems of L-er and 35S::ANT plants are similar in size (Fig. 6A,D), indicating that the increase in organ growth is not a consequence of a larger floral meristem. In addition, the sepal primordia are approximately the same size in both L-er and 35S::ANT flowers suggesting that 35S::ANT sepals do not initially consist of more cells. The height and width of stamen and carpel primordia in stage 6 (Fig. 6B,E) and stage 7 (Fig. 6C,F) L-er and 35S::ANT flowers is also quite similar. These data provide additional evidence that the predominant effects of ectopic ANT expression on stamens and carpels is increased cell expansion during later stages of floral organ development. The effects of ant mutations on floral meristem and organ growth were also investigated by two photon fluorescence microscopy. Stage 3 ant floral meristems are not as wide as wildtype stage 3 meristems (Fig. 6G). The average width of ant ⫺5 floral meristems was 40 ⫾ 4 mm, while that of wild type was 54 ⫾ 1 mm (average ⫾ standard deviation). The carpel primordia in a stage 6 ant ⫺5 flower (Fig. 6H) are approximately the same size as those in a L-er flower. By stage 7, both stamen and carpel primordia in ant ⫺5 flowers are thinner than those in wildtype flowers and appear to consist of fewer cell layers (Fig. 6I). These ant ⫺5 organ primordia have grown more in length than width, changing the shape of the primordia and making them somewhat difficult to stage. These results are consistent with mutations in ant affecting both the numbers and patterns of cell divisions in flowers. SEM analysis of ant ⫺5 mature floral organs indicates that in almost all cases, the size of epidermal cells are the same as in L-er flowers (data not shown). Ectopic Expression of ANT in ant ⴚ6 and ant ⴚ8 Because ectopic expression of ANT sometimes results in ovule defects similar to those observed in weak ant mutants, the ability of 35S::ANT to complement mutations in ant was investigated. Intermediate and weak ant alleles (ant ⫺6 and ant ⫺8, respectively) were used in these studies. ant ⫺6 contains the nucleotide substitution C679=T which converts Gln227 into a stop codon, while ant ⫺8 contains the nucleotide substitution G1267=A, which converts Ala422 into Thr. Interestingly, the ant ⫺6 allele displays an intermediate phenotype with slightly more expansion in the chalazal region of the ovule primordia than found in strong ant alleles (Fig. 2E) [Baker et al., 1997], yet contains a stop codon before the predicted AP2 repeat DNA binding domains. The Ala that is mutated in ant ⫺8 is conserved among different members of the AP2 protein family [Okamuro et al., 1997; Riechmann and Meyerowitz, 1998]. Since 35S::ANT plants are male sterile and mostly female sterile, the 35S::ANT construct was transformed into ant ⫺6/⫹ and ant ⫺8/⫹ plants. Kanamycin resistant progeny from these transformations were PCR genotyped [Jacobson and Moscovits, 1991]. Five 35S::ANT ant ⫺6 plants were obtained which all resembled ant ⫺6 plants with regard to floral organ number and floral organ size (Fig. 2D). However, a single integumentary-like structure grew around the nucellus in 35S::ANT ant ⫺6 ovules that is not present in ant ⫺6 ovules (cf. Fig. 2E,F). These 35S::ANT ant ⫺6 ovules closely resemble ovules found in the weak ant ⫺8 allele (Fig. 2G) [Baker et al., 1997]. Five 35S::ANT ant ⫺8 plants were obtained. Three of these plants resembled 35S::ANT plants with regard to floral organ number and size (Fig. 2H), while two resembled ant ⫺8 plants. The three lines with 35S::ANT-like larger flowers also mimicked the other 35S::ANT phenotypes (anther and ovule defects and the absence of epicuticular wax). Thus, ectopic expression of ANT is able to partially rescue the ovule defect of ant ⫺6 plants and ECTOPIC ANT EXPRESSION INCREASES FLORAL ORGAN GROWTH 233 Fig. 5. Expression of histone H4 in L-er and 35S::ANT flowers. L-er (A,B) and 35S::ANT (C,D). Small arrows, some of the histone H4 expressing cells in sepals. Wider arrows, petals of stage 10 flowers (B,D). A: Stage 6 L-er flower. B: Stage 10 L-er flower. C: Stage 6 35S::ANT flower. D: Stage 10 35S::ANT flower. se, sepal; pe, petal. TABLE 3. Number of Cells Expressing Histone H4 RNA in L-er and 35S⬋ANT Flowers* L-er 35S⬋ANT Stage 6 sepals 7.3 ⫾ 2.1 8.1 ⫾ 2.5 Stage 7 sepals 9.6 ⫾ 4.1 9.1 ⫾ 3.1 Stage 10 sepals 2.1 ⫾ 1.5 4.8 ⫾ 1.8 Stage 10 petals 21.1 ⫾ 6.9 22.6 ⫾ 3.6 *The data are indicated as averages ⫾ SD. The stage 6 sepal data are based on measurements from 12 sepals on 6 L-er flowers and 22 sepals on 11 35S⬋ANT flowers. The stage 7 sepal data are based on measurements from 12 sepals on 6 L-er flowers and 12 sepals on 6 35S⬋ANT flowers. The stage 10 sepal data are based on measurements from 17 sepals on 9 L-er flowers and 18 sepals on 9 35S⬋ANT flowers. The petal data are based on measurements from 11 petals on 11 L-er stage 10 flowers and 13 petals on 9 35S⬋ANT stage 10 flowers. rescues both the floral organ and ovule defects of ant ⫺8 plants. DISCUSSION Ectopic Expression of ANT Results in Larger Flowers Ectopic expression of ANT under a constitutive promoter results in increased growth of floral organs. This phenotype is the opposite of that resulting from loss of ANT function; ant mutants produce small, narrow floral organs [Baker et al., 1997; Elliott et al., 1996; Klucher et al., 1996; Schneitz et al., 1997]. The increased growth of 35S::ANT floral organs is manifested as increased cell division in sepals and increased cell expansion in petals, stamens, and carpels. Owing to the nature of the ANT expression pattern (broad initial expression with subsequent restriction to particular regions of developing organs), it is unclear whether the increased growth of 35S::ANT floral organs results from an increased level of ANT function and/or a lengthened period of ANT activity in developing floral organs. Some evidence for the latter of these possibilities is the lack of obvious size differences between L-er and 35S::ANT floral buds of stage 7 and younger. In ovules, ectopic expression of ANT results in increased growth of the proximal (funiculus) and distal (nucellus) elements of an ovule primordia but in decreased growth of the integuments arising from the chalazal region. In wild-type ovules, an initially broad ANT expression domain throughout the primordium becomes restricted to the central region of the primordium including the chalazal region and the distal part of the funiculus [Elliott et al., 1996]. Thus, the increased growth of the nucellus and funiculus in 35S:: ANT ovules is likely to result from ectopic expression of ANT in these regions of an ovule. The decreased growth of the outer integument in these ovules may result from some sort of integument-specific cosuppression of ANT. Alternatively, growth of the outer integument may requires a precise level of ANT expression. Differences in the levels and patterns of ANT expression in 35S:: ANT floral meristems and organs may also explain the inability of 35S::ANT to complement the ant ⫺6 mutation. Fig. 6. Two photon fluorescence microscopy of flower development in L-er, 35S::ANT, and ant ⫺5 flowers. L-er (A–C), 35S::ANT (D–F), and ant ⫺5 (G–I). A: L-er stage 3 flower, in which the sepal primordia have initiated. B: L-er stage 6 flower. C: L-er stage 7 flower. D: 35S::ANT stage 3 flower. E: 35S::ANT stage 6 flower. F: 35S::ANT stage 7 flower. G: ant ⫺5 stage 3 flower. The floral meristem is not as wide as a similar stage wild-type floral meristem. H: ant ⫺5 flower of approximately stage 6. While the carpel primordia are similar in size to those of wild type, the stamen primordia appear slightly thinner than stamen primordia of wild-type flowers. I: ant ⫺5 flower of approximately stage 7. Both the stamen and carpel primordia are thinner those of wild-type; growing more in length than width. Scale bars ⫽ 50 µm. ECTOPIC ANT EXPRESSION INCREASES FLORAL ORGAN GROWTH Comparison of ANT With Other Genes Involved in Regulating Cell Expansion The specificity of ANT ectopic expression on altering cell growth in flowers is similar to ectopic expression of another gene, NAP (NAC-like, activated by AP3/PI) [Sablowski and Meyerowitz, 1998]. This gene was identified as a target gene of the floral homeotic proteins APETALA3 (AP3) and PISTILLATA (PI) and is expressed in regions of petals and stamens during later floral stages (stage 8 to maturity). No NAP mutant has been isolated, but expression of NAP under the 35S promoter inhibits cell expansion in petals and stamens resulting in shorter organs. Thus NAP and ANT have opposite effects on cell growth in petals and stamens. NAP has been postulated to function in petals and stamens during the transition from organ growth resulting from cell division to that resulting from cell expansion [Sablowski and Meyerowitz, 1998]. Besides ANT, another member of the AP2/EREBP family of transcription factors has been shown to be involved in cell growth [Wilson et al., 1996]. The TINY gene seems to play a general role in promoting cell expansion. tiny plants are short in height, with stem epidermal cells that are reduced in length. tiny flowers have shorter stamens and pistils and rounder buds. Leaf epidermal cells from tiny have a different shape and are thicker than wild-type cells. Because tiny was identified as a dominant mutant in a screen using a Ds transposon containing a 35S promoter, the tiny phenotype is thought to arise from ectopic expression or overexpression of the gene [Wilson et al., 1996]. Thus, TINY and ANT appear to have opposite roles in cell growth and differ with regard to the organs which are affected and whether cell shape is altered. tiny, 35S:: NAP, and 35S::ANT plants all show reduced fertility [Sablowski and Meyerowitz, 1998; Wilson et al., 1996]. Few mutants or transgenes have been identified that increase organ growth. One mutant that causes increased growth of leaves and sometimes floral organs is revoluta (rev) [Talbert et al., 1995]. In the case of leaves, the increase in organ size is due to extra cell divisions, but the basis for the larger floral organs was not reported [Talbert et al., 1995]. Thus, it is unknown whether rev affects cell expansion in addition to cell division. The 35S::ANT transgene seems to be currently unique in its ability to confer increased organ growth that is specific to flowers. In addition, these plants exhibit few other phenotypes, none of which occurs during vegetative development. The ability to increase carpel and ovary size through the use of the 35S::ANT transgene might prove useful for producing larger yields in agriculture. Model for ANT Function ANT is required for floral organ initiation and growth [Baker et al., 1997; Elliott et al., 1996; Klucher et al., 1996; Schneitz et al., 1997]. The results presented in 235 this paper indicate that ANT is sufficient in flowers for organ growth but not organ initiation. The decreased number of floral organs in ant mutants result from a failure to initiate a normal number of organ primordia and not from abortion of organ primordia after initiation. Since ant floral meristems are smaller than wild type, the decrease in organ number may result from an insufficient number of meristematic cells for incorporation into organ primordia. The decreased size of ant floral organs results from changes in the number and orientations of cell divisions within developing floral organ primordia. Considering both the loss of function ant mutant phenotype as well as the 35S::ANT ectopic expression phenotype, the following model describes how ANT might control organ initiation and growth in flowers. The model proposes that ANT promotes cell growth. In the absence of ANT, cell growth is restricted. Studies in yeast and other systems have shown that cells must reach a critical size before dividing [Mitchison et al., 1997]. Cells below this size threshold are prevented from completing the cell cycle. Because of this close relationship between cell growth and cell proliferation, a decrease in cell growth can cause a reduction in cell division. The decreased numbers of cells in ant mutants is thus proposed to be a consequence of decreased cell expansion. Decreased cell division in ant floral meristems and young floral organ primordia ultimately leads to the presence of too few cells in young floral meristems, a reduction in organ initiation, and the development of smaller floral organs. In 35S::ANT plants, increased ANT function stimulates cell growth. This increased growth results in more cell divisions in sepals. In petals, stamens, and carpels increased cell growth is not accompanied by increased cell division. It remians unclear why the effects of increased growth are manifested in different ways in these different floral organs. Perhaps other factors involved in carrying out these growth processes are limiting in different organs. ACKNOWLEDGMENTS I thank Jannie Lee for doing the histone H4 in situs, Hajime Sakai for pHS-H4, Charles Gasser for ant ⫺5, ant ⫺6, ant ⫺7, and ant ⫺8 seeds, David Smyth for ant ⫺9 seeds and ANT cDNA, Dick Vogt for the use of his compound microscope and photography system and for help with two photon fluorescence microscopy, Madilyn Fletcher’s laboratory for the use of IPLab, Dana Dunkelberger and Shawn Vose for advice on the use of the SEM, and an anonymous reviewer for extremely helpful comments. REFERENCES Baker SC, Robinson-Beers K, Villanueva JM, Gaiser JC, Gasser CS. 1997. Interactions among genes regulating ovule development in Arabidopsis thaliana. Genetics 145:1109–1124. 236 KRIZEK Bechtold N, Ellis J, Pelletier G. 1993. In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. CR Acad Sci 316:1194–1199. Bowman JL, Smyth DR, Meyerowitz EM. 1991. Genetic interactions among floral homeotic genes of Arabidopsis. Development 112:1–20. Buttner M, Singh KB. 1997. Arabidopsis thaliana ethylene-responsive element binding protein (AtEBP), an ethylene-inducible, GCC box DNA-binding protein interacts with an ocs element binding protein. Proc Natl Acad Sci USA 94:5961–5966. Cleland RE. 1987. Auxin and cell elongation. In: Davis PJ, editor. Plant hormones and their role in plant growth and development. The Netherlands: Kluwer. p 132–148. Coen ES, Meyerowitz EM. 1991. The war of the whorls: genetic interactions controlling flower development. Nature 353:31–37. Coen ES, Romero JM, Doyle S, Elliott R, Murphy G, Carpenter R. 1990. Floricaula: a homeotic gene required for flower development in Antirrhinum majus. Cell 63:1311–1322. Depicker A, van Montagu M. 1997. Post-transcriptional gene silencing in plants. Curr Opin Cell Biol 9:373–382. Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQJ, Gerentes D, Perez P, Smyth DR. 1996. AINTEGUMENTA, an APETALA2like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 8:155–168. Fobert PR, Coen ES, Murphy GJP, Doonan J. 1994. Patterns of cell division revealed by transcriptional regulation of genes during the cell cycle in plants. EMBO J 13:616–624. Gaiser JC, Robinson-Beers K, Gasser CS. 1995. The Arabidopsis SUPERMAN gene mediates asymmetric growth of the outer integument of ovules. Plant Cell 7:333–345. Hanzawa Y, Takahashi T, Komeda Y. 1997. ACL5: an Arabidopsis gene required for internodal elongation after flowering. Plant J 12:863– 874. Heslop-Harrison J, Heslop-Harrison Y. 1970. Evaluation of pollen viability by enzymatically induced fluorescence: intracellular hydrolysis of fluorescein diacetate. Stain Tech 45:115–120. Hooley R. 1996. Plant steroid hormones emerge from the dark. Trends Genet 12:281–283. Jacobson DR, Moscovits T. 1991. Rapid, nonradioactive screening for activating ras oncogenic mutations using PCR primer-introduced restriction analysis (PCR-PIRA). PCR Methods Appl 1:146–148. Kagaya Y, Ohmiya K, Hattori T. 1999. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res 27:470–478. Kim Q-T, Tsukaya H, Uchimiya H. 1998. The CURLY LEAF gene controls both division and elongation of cells during the expansion of the leaf blade in Arabidopsis thaliana. Planta 206:175–183. Klimyuk VI, Carroll BJ, Thomas CM, Jones JDG. 1993. Alkali treatment for rapid preparation of plant material for reliable PCR analysis. Plant J 3:493–494. Klucher KM, Chow H, Reiser L, Fischer RL. 1996. The AINTEGUMENTA gene of Arabidopsis required for ovule and female gametophyte development is related to the floral homeotic gene APETALA2. Plant Cell 8:137–153. Krizek BA, Meyerowitz EM. 1996. Mapping the protein regions responsible for the functional specificities of the Arabidopsis MADS domain organ-identity proteins. Proc Natl Acad Sci USA 93:4063– 4070. Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. 1998. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature responsive gene expression, respectively, in Arabidopsis. Plant Cell 10:1391–1406. Ma H. 1994. The unfolding drama of flower development: recent results from genetic and molecular analyses. Genes Dev 8:745–756. Marzluff WF, Pandey NB. 1988. Multiple regulatory steps control histone mRNA concentrations. Trends Biochem Sci 13:49–52. Métraux J-P. 1987. Gibberellins and plant cell elongation. In: Davis PJ, editor. Plant hormones and their roles in plant growth and development. The Netherlands: Kluwer. p 296–317. Meyerowitz EM. 1997. Genetic control of cell division patterns in developing plants. Cell 88:299–308. Mitchison JM, Novak B, Sveiezer A. 1997. Size control in the cell cycle. Cell Biol Int 21:461–463. Müller A. 1961. Zur characterisierung der Blüten und Infloreszenzen von Arabidopsis thaliana (L.) Heynh. Kulturpflanze 9:364–393. Nakayama T, Iwabuchi M. 1993. Regulation of wheat histone gene expression. Crit Rev Plant Sci 12:97–110. Ohme-Takagi M, Shinshi H. 1995. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell 7:173–182. Okamuro JK, Caster B, Villarroel R, Montagu MV, Jofuku KD. 1997. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc Natl Acad Sci USA 94:7076– 7081. Regan SM, Moffatt BA. 1990. Cytochemical analysis of pollen development in wild-type Arabidopsis and a male-sterile mutant. Plant Cell 2:877–889. Reiser L, Fischer RL. 1993. The ovule and the embryo sac. Plant Cell 5:1291–1301. Riechmann JL, Meyerowitz EM. 1998. The AP2/EREBP family of plant transcription factors. Biol Chem 379:633–646. Robinson-Beers, Kay P, E. R, Gasser CS. 1992. Ovule development in wild-type Arabidopsis and two female-sterile mutants. Plant Cell 4:1237–1249. Running MP, Clark SE, Meyerowitz EM. 1995. Confocal microscopy of the shoot apex. In: Galbraith DW, Bohnert HJ, Bourque DP, editors. Methods in cell biology’’ San Diego: Academic Press. p 217–229. Sablowski RWM, Meyerowitz EM. 1998. A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3/PISTILLATA. Cell 92:93–103. Sakai H, Medrano LJ, Meyerowitz EM. 1995. Role of SUPERMAN in maintaining Arabidopsis floral whorl boundaries. Nature 378:199– 203. Schneitz K, Hulskamp M, Kopczak SD, Pruitt RE. 1997. Dissection of sexual organ ontogenesis: a genetic analysis of ovule development in Arabidopsis thaliana. Development 124:1367–1376. Schneitz K, Hulskamp M, Pruitt RE. 1995. Wild-type ovule development in Arabidopsis thaliana: a light microscopic study of cleared whole-mount tissue. Plant J 7:731–749. Sessions A, Yanofsky MF, Weigel D. 1998. Patterning the floral meristem. Semin Cell Dev Biol 9:221–226. Smyth DR, Bowman JL, Meyerowitz EM. 1990. Early flower development in Arabidopsis. Plant Cell 2:755–767. Stockinger EJ, Gilmour SJ, Thomashow MF. 1997. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc Natl Acad Sci USA 94:1035– 1040. Takahashi T, Gasch A, Mishizawa N, Chua N-H. 1995. The DIMINUTO gene of Arabidopsis is involved in regulating cell elongation. Genes Dev 9:97–107. Talbert PB, Adler HT, Parks DW, Comai L. 1995. The REVOLUTA gene is necessary for apical meristem development and for initiating cell divisions in the leaves and stems of Arabidopsis thaliana. Development 121:2723–2735. Vergani P, Morandinin P, Soave C. 1997. Complementation of a yeast Dpkc1 mutant by the Arabidopsis protein ANT. FEBS Lett 400:243– 246. Weigel D, Meyerowitz E. 1994. The ABCs of floral homeotic genes. Cell 78:203–209. Wilson K, Long D, Swinburne J, Coupland G. 1996. A dissociation insertion causes a semidominant mutation that increases expression of TINY, an Arabidopsis gene related to APETALA2. Plant Cell 8:659–671. Zhou J, Tang X, Martin GB. 1997. The Pto kinase conferring resistance to tomato bacterial speck disease interacts with proteins that bind a cis-element of pathogenesis-related genes. EMBO J 16:3207–3218.