Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Biology and consumer behaviour wikipedia , lookup
Behavioural genetics wikipedia , lookup
Gene desert wikipedia , lookup
Human genetic variation wikipedia , lookup
Gene therapy wikipedia , lookup
Gene nomenclature wikipedia , lookup
Pharmacogenomics wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Hardy–Weinberg principle wikipedia , lookup
Genome evolution wikipedia , lookup
Public health genomics wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Genetically modified crops wikipedia , lookup
Gene expression programming wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
Gene expression profiling wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Helitron (biology) wikipedia , lookup
Genomic imprinting wikipedia , lookup
Quantitative trait locus wikipedia , lookup
Genetic drift wikipedia , lookup
Genetic engineering wikipedia , lookup
Genome (book) wikipedia , lookup
Population genetics wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Designer baby wikipedia , lookup
History of genetic engineering wikipedia , lookup
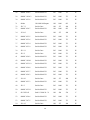
Vernalization Gene Architecture as a Predictor of Growth Habit in Barley By Douglas Heckart A thesis submitted to Oregon State University In partial fulfillment of the requirements for the degree of Bachelors of Science in Bioresource Research Applied Genetics and Crop and Soil Science Crop Breeding and Genetics Presented June 19, 2006 ©Copyright by Douglas L Heckart June 19, 2006 All rights reserved Bachelor of Science thesis of Douglas Heckart presented on June 19, 2006 Approved: Dr. Patrick Hayes Primary Faculty Mentor Date Dr. Thomas Chastain Secondary Faculty Mentor Date Ann Corey Research Advisor Date Dr. Anita Azarenko Director of Bioresource Research Program Date I understand that my thesis will become part of the permanent collection of Oregon Sate University Libraries. My signature below authorizes release of my thesis to any reader upon request. Douglas L. Heckart Date An Abstract of the Thesis of Douglas L Heckart for the degree of Bachelor of Science in Bioresource Resource Research and Crop and Soil Science presented on June 19, 2006. Title: Vernalization Gene Architecture as a Predictor of Growth Habit in Barley Abstract approved: _Dr. PatrickHayes___________________________________ Primary Research Mentor Date Winter hardiness in barley (Hordeum vulgare) is a trait targeted by breeding programs in order to expand the potential area of adaptation of fall-sown cereals. Vernalization requirement is an important factor in winter hardiness. A vernalization requirement is an extended period of low temperature required for transition from the vegetative to reproductive states. A model involving two vernalization genes that interact in an epistatic fashion to determine the vernalization phenotype has been proposed. To test this model, the vernalization phenotype was compared to the vernalization allele genotype for each entry in a breeding program nursery grown under fall- and spring-sown conditions. The number of growing degree days (GDD) from planting to Feekes growth stage 10.5 was recorded for each line in both experiments. Lines that did not reach Feekes 10.5 when spring-sown were assigned values of 1000 GDD. To calculate the growing degree day vernalization coefficient (GDD-VC), the fall sown GDD value was subtracted from the spring sown value for each line. The GDD-VC values ranged from 139 to 812. Facultative phenotypes were classified as <309 GDD-VC or less and winter phenotypes >409 and above. Genotypes were then assessed according to their vernalization allele configuration at the VRN-H1 and VRN-H2 loci. All lines had the “winter” allele at HvBM5A, the candidate gene for the VRN-H1 locus. Out of 54 lines, 46 had the “winter” allele at VRN-H2, where the ZCCT-H gene cluster is the candidate, classifying them as “winter” genotypes. Winter and facultative phenotypes were compared to winter and facultative genotypes. The phenotype was accurately predicted from genotype 94% of the time. Possible reasons for the <100% accuracy include residual heterogeneity and difficulty in classifying the growth habit of some lines under spring-sown conditions. Table of Contents Page Introduction 2 Methods and Materials 10 Results 14 Conclusions 19 Tables and figures 21 Literature cited 28 Acknowledgements 31 1 Introduction Barley (Hordeum vulgare subsp. vulgare) is a crop species of worldwide importance and an important genetic model for the Triticeae. Barley was one of the first domesticated crops originating from the Fertile Crescent approximately 10,000 years ago (Badr et al.,, 2000). This crop is important throughout the world because it has numerous end uses. In South America and parts of Asia, barley is traditionally used as a food crop. In the United States and Europe, barley is predominately used as an animal feed, although the most valuable use is as the primary source of malt for beer (Hayes, 2003). The inflorescence of barley has either one or three fertile florets per rachis node. The former leads to a “two-row” inflorescence and the latter to a “six-row” inflorescence”. These two inflorescence types define the major germplasm groups of barley. The division into the two-row and six-row germplasm groups is supported by numerous genetic diversity studies e.g. Matus et al., (2002). These genetic diversity studies were possible due to the abundant genetic resources available in barley. Barley is diploid (2n = 14), making it a model system for more genetically complex members of the Triticeae, such as hexaploid wheat. The genetic resources in barley include 48 comprehensive linkage maps (GrainGenes, wheat.pw.usda.gov/gg2/index.shtml), mapped QTLs, 348,620 Expressed Sequence Tags (www.harvest.ucr.edu) and 46,764 cloned mRNA (from “barleygene” NCBI). These genetic resources facilitate analyses of a range of traits, including the genes that define the major growth habit forms of barley. The growth habit forms of barley are spring, facultative, and winter. Agronomically speaking, spring forms are planted in the spring and harvested late summer; winter forms are planted in the fall and harvested the following summer; and 2 facultative forms can be planted in the fall or spring. Growth habit is important because it determines the range of adaptation. Winter and fall-sown facultative varieties generally yield more than spring-planted forms, provided that they have sufficient winter hardiness. However, a lack of winter hardiness limits the acreage of fall-sown cereals (Fowler et al., 2001). The ancestral growth habit of barley is the winter habit. The spring growth habit developed through loss of function of vernalization genes (Yan et al., 2004). In order to expand the potential area of adaptation of fall-sown cereals, winter hardiness has been the subject of intensive research in the Triticeae (Skinner et al., 2005). The three components of winter hardiness are low temperature tolerance, vernalization requirement, and photoperiod sensitivity (Skinner et al., 2005). The genetics of low temperature tolerance and photoperiod sensitivity were reviewed by Skinner et al., (2005) and Szucs et al., (2006). The focus of this research is on vernalization requirement. A vernalization requirement is an extended period of low temperature required for transition from the vegetative to the reproductive state. Without vernalization, genotypes that have a requirement will eventually flower, but the flowering time will be delayed by months and the flowering will be erratic (Karsai et al., 2001). The vernalization requirement ensures plants will be in a vegetative state during the winter, and maximum cold tolerance is achieved when plants are in a vegetative condition (Fowler et. al., 2001). An explicit relationship between growth habit and vernalization requirement was recently proposed by Von Zitzewitz et al., (2005) based on the cloning of candidate genes for the VRN-H1 and Vr-H2 loci in a diverse sample of barley germplasm. According to this model, winter forms require vernalization due to having the “winter” form of the allele at the VRN-H1 loci and dominant “winter” allele at the VRN-H2 loci. “Winter” 3 alleles refer to DNA sequences encoding a competent repressor of VRN-H1 at the VRNH2 locus and a competent binding site for the repressor at the VRN-H1 locus. Spring habit genotypes do not require vernalization due to a complete deletion of the gene encoding the repressor and/or a deletion of the repressor binding site in VRN-H1. Facultative genotypes lack the DNA sequence encoding the repressor but have a competent repressor binding site in VRN-H1. Facultative genotypes are often sensitive to short photoperiod, which accomplishes the same effect as vernalization: it maintains plants in a vegetative state during the winter (Mahfoozi et. al., 2001). This comprehensive model for the genetics of vernalization requirement is built on nearly forty years of prior research. Takahashi and Yasuda (1970) proposed a three locus model to explain the genetic basis of spring and winter growth habit. The loci were named “Spring habit, (Sh) and were assigned linkage map positions on chromosomes 4 (Sh1), 5 (Sh3), and 7 (Sh2). Variation in Sh3 (now Vrn3), is observed only in rare barley germplasm from extremely high or low altitudes (Yasuda et al., 1993). Barley chromosome numbering nomenclature has changed such that 4 = 4H; 5 = 1H; and 7 = 5H. Furthermore, the loci are now designated as “Vernalization, Vrn” and the locus numbering has also changed. The renumbering is based on the cloning of candidate genes in Triticum monococcum (Yan et al., 2003) and barley (Dubcovsky et al., 2004; Von Zitzewitz et al.,, 2005). The current nomenclature and chromosome assignments for barley are: VRN-H1 (5H), VRN-H2 (4H) and Vrn-H3 (1H). Vrn-H3 has not yet been cloned, but since allelic variation is not seen in most germplasm, the genetic control of vernalization is thought to be best explained by a two-locus model in most cases. 4 The genetic basis of the two-locus epistatic interaction in barley was described in detail by Von Zitzewitz et al., (2005). The candidate genes of interest are the ZCCT-H gene family at the VRN-H2 locus and the HvBM5A gene at the VRN-H1 locus. The ZCCT-H gene family consists of three physically linked genes (Yan et. al., 2004). The VRN-H2 complex locus encodes a flowering repressor. The HvBM5A gene encodes a meristem identity gene. In the first intron of this gene, there is a binding site for the repressor encoded by VRN-H2. To recapitulate the relationship of growth habit and the genetics of vernalization, winter types require vernalization because they have the VRNH2 gene that encodes the repressor and they have Vrn-H1 alleles that have a binding site for the repressor. Facultative genotypes do not require vernalization because even though they have VRN-H1 alleles with the repressor binding site, they are completely lacking the VRN-H2 gene that encodes the repressor. Spring types do not require vernalization because they have VRN-H1 alleles that do not have a repressor binding site; the VRN-H2 gene may or may not be present. These genetic models for vernalization requirement require accurate measurement of the vernalization requirement phenotype. The vernalization requirement phenotype can be calculated from data collected on plant material grown under controlled or field environments. In either case, the basic measure of vernalization requirement is the difference between the growth and/or development of plants with and without vernalization. Controlled environments have been used for measuring vernalization requirement in oat (Holland et al.,, 2002) and barley (Karsai et al., 2005). In the latter, the vernalization requirement was calculated as the difference, in number of days until flowering, between unvernalized and vernalized plants grown under the same light 5 intensity, photoperiod duration, and temperature. The principal drawbacks to controlled environments are high cost and the limited number of plants that can be assayed. An advantage, and a drawback, to controlled environments is that they cannot reflect the complex environmental cues plants receive under field conditions. Under field conditions, vernalization requirement can be measured as the difference in the number of days to flowering between spring-sown and fall-sown experiments. As described by Francia et al., (2003), the data from the two sowing dates can be standardized by using the number of days from April 1 to flowering (heading) rather than the number of days from planting. Genotypes that do not flower under spring sown conditions are assigned a high value of 1000 days. A problem with using calendar days to heading is that it does not take into account the role of temperature and its effects on plant growth and development. A solution is to use the difference in the number of growing degree days (GDD) between the spring and fall plantings (T. Chastain, personal communication, 2006). Regardless of whether the plant material is grown under controlled environment or field conditions, and whether or not GDD or calendar days are used, there are “maturity per se” genes that can influence the time elapsed between emergence and flowering (Laurie, 1997) An extreme example of the effect of such maturity genes is that of rice transformed with the maturity gene LEAFY from Arabidopsis. Transgenic plants were 30 days earlier to flower than the wild type (He et al.,, 2000). Another complication is that plants may flower without vernalization but the flowering pattern may be so erratic that it would be agronomically unacceptable to consider such plants as “spring” or “facultative”. An alternative measure of vernalization requirement that should avoid the confounding 6 effects of maturity per se genes and erratic flowering is the final leaf number (FLN). Fowler et al., (2001) have used this measure successfully with plants grown with hydroponics under controlled environment conditions. This measure of the vernalization phenotype, however, would be costly to implement with field grown material., Finally, spring-sown genotypes can simply be assigned a rating based on an integrative, subjective scale of growth habit at a given time point. The advantage of such an approach is that it is rapid and may be suitable for agronomic classification, if not for genetic analysis. In the Oregon State University Barley Breeding Program, spring-sown plots from the winter and facultative breeding program are rated on a 1 – 9 scale, where 1 = maturity comparable to the spring habit check variety, 5 = normal growth and development but late maturity, and 9 = only vegetative growth. Understanding the genetics of vernalization requires scrupulous phenotyping, as described in the preceding section, and it also requires cost-effective and reliable genotyping. There is a range of techniques available for determining differences in DNA sequence between varieties. While complete sequencing of the gene in each variety is possible, it is too expensive and may be unnecessary. For example, the candidate gene for VRN-H1, HvBM5A, is 11kb (spring), 17kb (winter) and the key region that differentiates winter and spring alleles is 435bp (Von Zitzewitz et al., 2005). In the case of the candidate for VRN-H2, the genes of interest (ZCCT-Ha and ZCCT-Hb) form the tandemly linked ZCCT gene family of 1.7kb each. The spring allele is null: the gene family is completed deleted (Dubcovsky et al., 2005). Therefore, the Polymerase Chain Reaction (PCR) can be used with gene-specific primers for the critical region of HvBM5A and to determine the presence or absence of the ZCCT gene family (Von Zitzewitz, 7 2005). Primers for the HvBM5A gene were designed to amplify the repressor binding site that distinguished winter and facultative (presence0 from spring (absence) genotypes. In the case of the VRN-H2 locus, the gene-specific primers function also as a dominant marker since an amplicon is obtained only when the ZCCT gene family is present. Dominant markers are problematic because the absence of an amplicon may be due to absence of the gene, or to failure of the PCR. Therefore, in the case of dominant markers, the assays may need to be repeated in order to have complete confidence in the case of the null allele. The two-locus epistatic genetic model proposed by Von Zitzewitz et al., (2005) was based on comparison of gene sequences between genotypes whose vernalization requirement had been previously determined in controlled environment and field tests. Although the model is compelling, it remains to be validated. There are three approaches to validation, each with advantages and disadvantages. One approach uses transgenic technology to either constitutively or transiently over-express and under-express target genes (Loukoianov et al., 2005). This approach requires considerable time and expense, since only a limited number of barley genotypes are amenable to transformation. An alternative genetic approach that has the advantage of assaying candidate genes in a native genomic context is monitoring phenotype and genotype in progeny segregating for alleles at one or both Vrn loci. Szucs et al., (personal communication) have taken this approach and determined Vrn allele type and vernalization requirement in three F2 populations derived from crosses of contrasting parental genotypes. The third approach, and that taken with this project, is to determine Vrn allele type and vernalization requirement in an array of breeding program germplasm derived from crosses of parents 8 known to differ in Vrn allele type and growth habit. In this project 46 breeding lines in the Oregon Elite Line Trial (ORELT) were derived from crosses involving four parents. Strider and Kold are winter varieties (VRN-H1 recessive,VRN-H2 dominant); 88Ab536 is facultative (VRN-H1 recessive,VRN-H2 recessive); and Orca is a spring type (VRN-H1 dominant, VRN-H2 recessive). In this project, these lines were genotyped for Vrn alleles and phenotyped for Vrn requirement in order to answer the question “can the two-locus Vrn genetic model be used to predict the growth habit of experimental lines in a breeding program?” In addition, two previously uncharacterized varieties from the Pacific Northwest – Hundred (Washington) and Eight-twelve (Idaho) – were included in the genotyping and phenotyping assays. 9 Methods and Materials Germplasm A total of 427 lines in the Oregon State University Barley Breeding Project, were phenotyped for this research. Subsequently, this research focused on one breeding nursery – the Oregon Elite Line Trial (ORELT). The varieties and experimental lines in this nursery, and their pedigrees, are shown in Table 1. There were 54 entries in the trial; Strider was repeated twice, so there are 53 unique entries. There were six varieties (88Ab536, Kold, Strider, Eight twelve, and Maja) and 47 experimental lines. The single replicate ORELT nursery was planted in the fall and in the spring at the Hyslop Agronomy Farm, near Corvallis, Oregon. The soil type at this location is Woodburn silt loam with a 0 – 3% slope. The fall-sown nursery was planted on October 14, 2004. Each entry was represented by one 4.6 meter long, six-row plot. The spring sown-nursery was planted on March 9, 2005. Each entry was represented by one 1.6 meter long, one-row plot. Fertilization and weed control were in accordance with recommended practice for the location. Supplemental irrigation was applied only to the spring-sown nursery. Phenotyping The growth stage was recorded on 211 fall-sown plots and 216 spring sown plots. However, as the study focused on the ORELT, the subsequent information is specific to the 54 entries that were the basis of this research. Growth staging: The Feekes growth stage for each entry was recorded every two weeks (Large, 1954). No further measurements were taken after Feekes 10.5, when the inflorescence was fully emerged. 10 Growing degree day vernalization coefficient: The GDD values used in this experiment were obtained in from the Oregon State University Hyslop Agronomy farm website (http://www.ocs.orst.edu/corv_dly.html). The GDD values were calculated each day. The high and low temperatures were averaged and the base temperature was subtracted to produce the daily GDD value. The base temperature used in this calculation was 10°C. The number of GDD from planting to Feekes 10.5 was calculated for each entry in the fall and spring-sown experiments. Growing degree days Entries that failed to reach Feekes 10.5 in the spring-sown nursery were assigned a GDD value of 1000. The vernalization requirement was calculated as the difference in these GDD values between the spring and fall sown experiments. This is referred to as the Growing Degree Day Vernalization Coefficient (GDD-VC). An example of this calculation is shown in Table 2. The spring-sown ORELT was also rated for “growth habit” when the spring variety “Baronesse” was at Feekes 10.5. A 1 – 9 scale was used to characterize growth habit, where 1 = spring growth habit and maturity phenotype comparable to “Baronesse” and 9 = a completely vegetative growth habit and maturity phenotype comparable to the winter check “Strider”. Genotyping Genotyping was performed at two locations: the USDA/ARS regional genotyping lab at Pullman, Washington and the Oregon State University barley genotyping facility. DNA was extracted from independent seed samples at each location. DNA was extracted from a single plant using a QIAGEN DNeasy miniprep kit (Qiagen, Valencia, CA) at OSU. These genomic DNA samples were used for assay of the candidate genes for the VRN-H1 and VRN-H2 loci. 11 At Oregon State University, primers were designed to test for the presence or absence of ZCCT-Ha and ZCCT-Hb. These two genes are very similar and the PCR amplifies a 307bp fragment for ZCCT-Ha and a 273bp fragment for ZCCT-Hb. The forward primer sequence was 5’ CCT AGT TAA AAC ATA TAT CCA TAG AGC 3’ and the reverse primer sequence was 5’ GAT CGT TGC GTT GCT AAT AGT G 3’. The winter allele has an expected size of 307bp + 273bp and the spring allele an expected size of 0bp. For VRN-H1, primers were designed to amplify the intron 1 region of HvBM5A that is hypothesized to be the binding site for the repressor encoded by VrnH-2 (Von Zitzewitz et al.,, 2005). The forward primer sequence was 5’ TGA GGG TAT GAG TGG CGC TAG 3’and the reverse primer sequence was 5’ TCT CAT AGG TTC TAG ACA AAG CAT AG 3’. The winter allele has an expected size of 435bp and the spring allele an expected size of 0bp. The PCR amplicons were separated on 1% agarose and stained with ethidium bromide. At the USDA/ARS lab and OSU barley lab, Corvallis OR, the presence or absence of the ZCCT and HvBM5A genes in each ORELT line was evaluated. The assays performed at the two laboratories were considered to be independent replicates. In the case of the dominant alleles at HvBM5A, the allele scoring between the two labs was compared directly. In the case of the ZCCTa and ZCCTb dominant markers, results from the two labs were compared and the presence of an amplicon at either of the two labs was considered to be diagnostic of the “winter” allele. The ZCCT-H genotyping assay was repeated again at Corvallis and produced the same allele configuration. The predictive power of the genotyping was assessed by assigning a growth habit classification (winter, facultative, or spring) to each genotype based on the nomenclature 12 of Von Zitzewitz et al., (2005). The growth habit assignments were then compared with the vernalization requirement phenotype, as measured by GDD. The number of correct predictions was divided by the total to determine the percentage of accurate predictions. Data were recorded in Excel spreadsheets and this software was used for all calculations and construction of phenotypic frequency distributions. 13 RESULTS Growing conditions in both the fall-sown and spring-sown field trials were adequate for scoring the phenotypes recorded in this study. No significant winter injury was observed in the fall-sown trials. High winds and heavy spring rains led to significant lodging in the fall-sown trials, but these conditions occurred after Feekes 10.5 and therefore, did not affect the estimation of growth stages. The number of GDD from planting to Feekes 10.5 in fall and spring-sown trials, and calculated GDD vernalization coefficients (GDD-VC) for the check varieties and the experimental line (Kab 47), replicated across multiple experiments are shown in Table 3. Standard errors for 88Ab536, Kab 47, and Strider were quite high because the growth stage data were taken at two-week intervals. For instance, even if an entry was close to 10.5 at the date of recording, it was not recorded as having reached that stage until the date of the next reading. In the case of 88Ab536, Feekes 10.5 was reached at 122, 188, and 188 GDD in the Oregon Elite Line Trial (ORELT), the Winter Barley Advanced Yield Trial (WBADV), and the Winter Barley Preliminary Yield Trial (WBPYT), respectively. When the Feekes growth stages were recorded for entries in these three nurseries on April 30, 2005, 88Ab536 in the ORELT nursery had attained Feekes 10.5, whereas in the WBADV and WBPYT nurseries it was staged as 10.4. Only a few dyas rather than two weeks were needed for 88Ab536 to attain Feekes 10.5 in WBADV and WBPYT. By way of comparison, Ms. Ann Corey, Senior Research Assistant with the OSU Barley Project, took daily notes on heading date in the same experiments and recorded a Julian heading date of 99 days for 88Ab536 in each of the three experiments. For consistency, the fall-sown GDD value for the ORELT was used for calculating the GDD-VC for each 14 of the check varieties and Kab 47, rather than the mean value. This value is shown in parentheses in Table 3. Under fall-sown conditions, there was a large range in the number of GDD from planting to Feekes 10.5. There was a difference of 606 GDD between the earliest (88Ab536 and Kab 47) and latest (Hundred and Kold) varieties. All check varieties except Kold, Strider, and Hundred reached Feekes 10.5 under spring-sown conditions. According to the spring-sown GDD data, Kold, Strider, and Hundred are the three varieties that would be considered to have a vernalization requirement because they did not flower within an agronomically acceptable time frame. These varieties had the largest values GDD-VCs (812, 715, and 715 respectively). By way of comparison, the spring habit variety Orca had a GDD-VC of 206. The other check varieties, and Kab 47, had GDD-VCs between those of Orca and Kold. Thirty-five experimental lines did not reach Feekes 10.5 in the spring-sown experiment (Figure 1). These entries with GDD-VCs greater or equal to those of Strider, Kold, and Hundred can be considered to have a vernalization requirement. Five entries, including the variety Eight twelve, had intermediate GDD-VCs of 409 and would be too late maturing for agronomic performance under spring-sown conditions and are therefore also considered to also have a vernalization requirement. Genotypes with GDD-VCs between 139 and 302 are considered facultative. The results from the amplification of the VRN-H1 critical region with allelespecific primers were in complete agreement between the USDA/ARS and OSU labs. All genotypes showed the expected 435bp product, which is the “winter” type allele according to Von Zitzewitz et al., (2005). The fact that the previously uncharacterized 15 winter varieties Hundred and Eight twelve have the winter allele at this locus is further evidence for the role of the VRN-H1 critical region as a binding site for the repressor encoded by VRN-H2 (Von Zitzewitz et al.,, 2005). All experimental lines in the ORELT are descended from crosses among Strider, 88Ab536, Kold, and Orca (see pedigrees in Table 1). Of these parents, all have been genotyped, except for Orca, and all have the winter allele at VRN-H1. The VRN-H1 allele architecture of Orca can be inferred based on parentage and phenotype. The parents of Orca are Calicuchima and Bowman. Calicuchima, a parent of Orca, does not have a vernalization requirement and was genotyped and identified to have the VRN-H2 gene and a unique “spring” allele at VRNH1 (P. Szucs, personal communication). Bowman is a spring variety from North Dakota and it is unlikely that it has a winter allele at VRN-H1. Therefore, it is reasonable to hypothesize that Orca has a spring allele at VRN-H1. Fourteen ORELT entries have Orca in their pedigree, yet all of these lines have the Strider (winter) allele at VRN-H1. This is probable evidence of stringent selection against the inferred Orca allele at this locus. The Strider/Orca lines were developed by J. Von Zitzewitz for his Bio Resource Research (BRR) undergraduate thesis at Oregon State University. His goal was to combine winter hardiness (from Strider) with Barley Yellow Dwarf Virus (BYDV) resistance from Orca. Only the lines surviving a low temperature event at Corvallis in December 1998 were advanced, and it is likely that winter survival was associated with the Strider allele at VRN-H1. According to Skinner et al., (2005), the VRN-H1 locus is associated with low temperature tolerance and is a candidate for the low temperature tolerance QTL Fr-H1. The linked cluster of HvCBF loci is a candidate for the Fr-H2 QTL. Thus, selection for winter survival in the 16 segregating progeny of Strider/ Orca crosses would have led to fixation of the Strider alleles at loci in this region of the genome. There was not complete agreement between labs for the VRN-H2 locus. At the USDA/ARS lab, no amplification was obtained for ORELT lines 17, 23, 29, 41, 48, 52, 53 and at OSU no amplification was obtained for ORELT lines 26 and 27. Possible explanations for these discrepancies between laboratories are: (i) PCR failure, (ii) sample contamination, and (iii) heterogeneous samples. PCR failure is a problem with dominant markers, such as the primers for VRN-H2, since the ZCCT gene family is either present or completely deleted. Sample contamination is always a possibility, but in each laboratory standard operating procedures are designed to minimize the likelihood of this happening. Entries in the ORELT were at the F6, or more advanced generation at the time of DNA extraction. However, some within-line heterogeneity is always possible. Unless winter breeding lines are routinely screened under spring-planted conditions, it is possible that “winter phenotype” lines are actually mixtures of winter and facultative types. DNA extractions are performed on a single plant from a single seed representing each variety or selection. In the case of heterogeneous varieties or lines, the random selection of the seed for DNA extraction could lead to contrasting conclusions regarding the presence or absence of VRN-H2. The characterization of growth habit based on spring-sown phenotype would depend on the percentage of seeds within the seed lot positive for VRNH2. PCR failure and/or heterogeneous seed lots are the most likely explanations for the discrepancies, since the VRN-H2 assay involves a dominant marker. For the purposes of this project, the presence of a PCR product at one or both labs was considered to be diagnostic of the presence of VRN-H2. By this criterion, the OSU data would be correct 17 for the seven lines that failed to amplify at the USDA/ARS lab and the USDA/ARS data would be correct for the two lines that failed to amplify at the OSU lab. This hypothesis was tested by performing new, independent, PCR reactions at OSU. All lines that previously amplified at OSU, but not at USDA/ARS, were positive for VRN-H2 and the two lines that were negative at OSU, remained negative for this allele. These genotyping results were then considered in the context of the phentoyping results. Since all ORELT entries have the winter allele at VRN-H1, the genetic model to account for vernalization requirement reduces to allelic variation at the VRN-H2 locus and all entries in the experiment can be classified as winter or facultative, per the definition of Von Zitzewitz et al., (2006). This growth habit classification, where “w” = winter and “f” = facultative is shown for each entry in the ORELT, together with the fall and spring-sown GDD values, and the GDD-VC in Table 4. With the exception of three entries – ORELT 18, 23, and 24 - all genotypes with GDD-VCs > 400 are classified as winter and those with GDD-VCs < 400 are classified as facultative. Therefore, the VRNH2 allele type is an excellent predictor of growth habit, but it failed in 3/54 cases. ORELT 18, 23, and 24 were lines with GDD-VCs of 236, 236, and 139 respectively: based on phenotype they are clearly facultative. However, 18 and 24 were scored as VRN-H2 positive at the USDA/ARS lab and 24 was scored as VRN-H2 positive at the OSU lab. 18 Conclusions The model proposed by Von Zitzewitz et al., (2005) has essentially been validated by this study. The GDD-VC phenotype was predicted by the Vrn allele phenotype in 51 out of 54 cases. This accuracy level is quite acceptable and indicates that the Vrn phenotype is a suitable target for marker assisted selection (MAS). However, there remains the question of why the correlation was not perfect. There are several possible explanations for the discrepancy. These include residual heterogeneity, DNA contamination, or ambiguity in rating the phenotype. The USDA/ARS lab has been contacted to determine if reserve tissue is available for re-extraction of DNA. Residual heterogeneity will be assessed in the summer of 2006, as the ORELT is planted for a second time for assessment of vernalization requirement. Residual heterozygosity is hypothesized to account for the two lines (26 and 27) that were winter habit according to their GDD-VC’s and have the VRN-H2 allele according to the USA/ARS data, but not the OSU data. Ambiguity in scoring growth stages under spring planted conditions may account for some discrepancies. For reasons that are not understood, many winter genotypes show erratic flowering under spring-sown conditions. The plant remains in an overall vegetative state but one or two inflorescences are formed. Technically, the plant reaches Feekes 10.5 when these few inflorescences are at the appropriate stage. Three ORELT entries are VRN-H2 positive, but had GDD-VC’s of 236,236, and 139 respectively. These same entries were rated 4, 7, and 7, respectively, for growth habit under spring-sown conditions by Dr. Pat Hayes. This is a subjective rating scale where 1 = flowering at the same time as the local spring check (Baronesse) and 9 = completely 19 vegetative. These intermediate ratings confirm that the spring growth habit was not entirely normal and the erratic growth habit could result from the presence of VRN-H2. There are two recommendations that can be made for future research in this and for Vrn MAS. Genotyping can be improved by augmenting the VRN-H2 genotyping with a codominant assay. This can be done by using a linked gene that shows complete linkage disequilibrium with VRN-H2. The HvSnf2 gene is good candidate for this type of assay (Von Zitzewtiz et al., 2005). This gene is distal to ZCCT and is present in all “spring”, “winter” and “facultative” genotypes. Phenotyping methods can also be improved. There were ambiguities between checks in different nurseries. These differences were often misleading due to the two week period between dates when plants were evaluated. It was possible for a line to reach Feekes 10.4 on a measurement date and 10.5 several days later, yet the date recorded would have been two weeks later. This problem can be resolved by evaluating plants daily. In some instances there are lines that flower under spring-sown conditions, but not in an agronomically acceptable fashion. In these cases the FLN could be measured and compared to FLN of spring and winter checks. 20 Table 1. Varieties and experimental lines, and their pedigrees, in the Oregon Elite Line nursery planted at Corvallis, Oregon in the fall of 2004 and spring of 2005. ORELT No 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 Variety or Selection 88Ab 536 Kold Strider Kab 47 Hundred Eight-Twelve Maja J2-5-1 StabBC 42-3-2 J1-8-17 J2-5-4 J2-12-2 Stab 113/Kab 43-1 J2-5-12 J1-13-1 Stab 113/Kab 50-22 StabBC 50-7-3 J2-6-19 StabBC 182-6-5 Stab 47/Kab 51-20 J2-15-1 StabBC 50-7-6 StabBC 50-9-1 StabBC 50-7-1 Stab 47/Kab 51-9 StabBC 50-9-2 Stab 47/Kab 51-7 StabBC 42-4-5 StabBC 182-4-2 StabBC 42-3-8 Strider J2-17-2 StabBC 42-4-7 J1-20-2 StabBC 42-3-5 StabBC 42-3-6 StabBC 42-3-4 StabBC 42-3-9 J2-5-11 J2-5-2 StabBC 42-4-2 StabBC 182-4-1 Pedigree Ne 76129/Morex/Morex 1285/Astrix OR 1860164/ Steptoe Kold/88Ab536 Strider/88Ab536 Strider/Orca Strider/88Ab 536 Strider/Orca Strider/Orca Strider/Orca Stab 113/Kab 43 Strider/Orca Strider/Orca Stab 113/Kab 50-22 Strider/88Ab 536 Strider/Orca Strider/88Ab 536 Stab 47/Kab 51-20 Strider/Orca Strider/88Ab 536 Strider/88Ab 536 Strider/88Ab 536 Stab 47/Kab 51-9 Strider/88Ab 536 Stab 47/Kab 51-7 Strider/88Ab 536 Strider/88Ab 536 Strider/88Ab 536 OR 1860164/ Steptoe Strider/Orca Strider/88Ab 536 Strider/Orca Strider/88Ab 536 Strider/88Ab 536 Strider/88Ab 536 Strider/88Ab 536 Strider/Orca Strider/Orca Strider/88Ab 536 Strider/88Ab 536 21 43 44 45 46 47 48 49 50 51 52 53 54 StabBC 42-3-10 StabBC 42-4-8 StabBC 42-3-7 StabBC 42-4-3 J1-8-16 StabBC 42-3-3 J2-5-3 StabBC 42-3-11 S113/K50-21 StabBC 42-4-6 StabBC 50-7-5 J1-14-2 Strider/88Ab 536 Strider/88Ab 536 Strider/88Ab 536 Strider/88Ab 536 Strider/Orca Strider/88Ab 536 Strider/Orca Strider/88Ab 536 Stab 113/Kab 50 - 21 Strider/88Ab 536 Strider/88Ab 536 Strider/Orca 22 Table 2. An example of calculation of the growing degree day vernalization coeffiecient (GDD-VC). Growing degree days from planting to Feekes growth scale 10.5 for fall planting subtracted from GDD value for spring planting. GDD to heading Variety Spring - Fall = GDD-VC 88Ab536 424 - 122 = 302 Strider 1000 - 188 = 23 812 Table 3.Growing degree day vernalization coefficients (GDD-VCs) for the check varieties and experimental selection (Kab 47) that were replicated in fall and spring-sown field trials at Corvallis, Oregon in the 2004-2005 crop year. Growing degree day vernalization Spring coefficient Variety or Fall sown sown (GDD-VC)* selection GDD GDD 88 AB 536 166 (122)** + 22 424 302 EightTwelve 188*** 597 409 Hundred 285 1000 715 Kab 47 166 (188)+22 424 236 Kold 285 1000 715 Maja 188 424 236 Orca 113 319**** 206 Strider 212 (188)+22 1000 812 * The GDD-VC was calculated as (Spring-sown GDD - Fall-sown GDD) ** ORELT nursery values are shown in parentheses for the entries that did not have identical GDD values in all nurseries. ***If no standard error indicated, the number of GDD was the same in all experiments. **** One observation only 24 Table 4. Growing degree day vernalization coefficients (GDD-VCs) and growth habit classification, based on VRN-H1 and VRN-H2 allele genotypes for commercial varieties and experimental lines in the Oregon Elite Lines Trial (ORELT) in the 2004-2005 crop year. ORELT Variety or no. Selection 1 88Ab 536 Fall Spring GDD-VC growth habit Pedigree GDD GDD classification Ne 76129/Morex/Morex 122 424 302 F 2 Kold 1285/Astrix 285 1000 715 W 3 Strider OR 1860164/Steptoe 188 1000 812 W 4 Kab 47 Kold/88Ab536 188 424 236 F 5 Hundred 285 1000 715 W 6 Eight-Twelve 188 597 409 W 7 Stab 113 Strider/88Ab536 188 424 236 F 8 J2-5-1 Strider/Orca 285 1000 715 W 9 StabBC 42-3-2 Strider/88Ab 536 285 1000 715 W 10 J1-8-17 Strider/Orca 285 1000 715 W 11 J2-5-4 Strider/Orca 188 1000 812 W 12 J2-12-2 Strider/Orca 188 1000 812 W 13 Stab 113/Kab 43-1 Stab 113/Kab 43 188 424 236 F 14 15 J2-5-12 J1-13-1 Strider/Orca Strider/Orca 285 188 1000 1000 715 812 W W 16 Stab 113/Kab 50-22 Stab 113/Kab 50-22 285 424 139 F 17 StabBC 50-7-3 Strider/88Ab 536 285 1000 715 W 18 J2-6-19 Strider/Orca 188 424 236 W 19 StabBC 182-6-5 Strider/88Ab 536 285 1000 715 W 20 Stab 47/Kab 51-20 Stab 47/Kab 51-20 188 424 236 F 21 J2-15-1 Strider/Orca 188 1000 812 W 22 StabBC 50-7-6 Strider/88Ab 536 188 1000 812 W 23 StabBC 50-9-1 Strider/88Ab 536 188 424 236 W 24 StabBC 50-7-1 Strider/88Ab 536 285 424 139 W 25 Stab 47/Kab 51-9 Stab 47/Kab 51-9 188 424 236 F 26 StabBC 50-9-2 Strider/88Ab 536 285 1000 715 W 27 Stab 47/Kab 51-7 Stab 47/Kab 51-7 188 1000 812 W 25 28 StabBC 42-4-5 Strider/88Ab 536 285 1000 715 W 29 StabBC 182-4-2 Strider/88Ab 536 285 1000 715 W 30 StabBC 42-3-8 Strider/88Ab 536 285 1000 715 W 31 Strider OR 1860164/Steptoe 188 1000 812 W 32 J2-17-2 Strider/Orca 188 597 409 W 33 StabBC 42-4-7 Strider/88Ab 536 188 1000 812 W 34 J1-20-2 Strider/Orca 188 597 409 W 35 StabBC 42-3-5 Strider/88Ab 536 285 1000 715 W 36 StabBC 42-3-6 Strider/88Ab 536 285 1000 715 W 37 StabBC 42-3-4 Strider/88Ab 536 285 1000 715 W 38 StabBC 42-3-9 Strider/88Ab 536 285 1000 715 W 39 J2-5-11 Strider/Orca 285 1000 715 W 40 J2-5-2 Strider/Orca 285 1000 715 W 41 StabBC 42-4-2 Strider/88Ab 536 285 1000 715 W 42 StabBC 182-4-1 Strider/88Ab 536 285 1000 715 W 43 StabBC 42-3-10 Strider/88Ab 536 285 1000 715 W 44 StabBC 42-4-8 Strider/88Ab 536 285 1000 715 W 45 StabBC 42-3-7 Strider/88Ab 536 285 1000 715 W 46 StabBC 42-4-3 Strider/88Ab 536 285 1000 715 W 47 J1-8-16 Strider/Orca 188 597 409 W 48 StabBC 42-3-3 Strider/88Ab 536 285 1000 715 W 49 J2-5-3 Strider/Orca 285 1000 715 W 50 StabBC 42-3-11 Strider/88Ab 536 285 1000 715 W 51 S113/K50-21 Stab 113/Kab 50 - 21 188 424 236 F 52 StabBC 42-4-6 Strider/88Ab 536 285 1000 715 W 53 StabBC 50-7-5 Strider/88Ab 536 285 1000 715 W 54 J1-14-2 Strider/Orca 188 597 409 W 26 Figure 1. Growing degree day vernalization coefficient (GDD-VC) for the eight check varieties and 46 experimental lines in the Oregon Elite Line Trial (ORELT) based on fall and spring-sown field trials at Corvallis, Oregon in the 2004-2005 crop year. 35 30 Number of Lines 25 20 15 10 5 0 139 236 302 409 Vrn requirement GDD-VC 27 715 812 Literature Cited Badr, A., K. Muller, R. Schafer-Pregl, H. El Rabey, S. Effgen, H.H. Ibrahim, C. Pozzi, W. Rohde, and Salamini. 2000. On the origin and domestication history of barley (Hordeum vulgare). Mol. Biol. Evol. 17(4):499-510. Borner, A., G. Buck-Sorlin, P.M. Hayes, S. Malyshev, and V. Korzun. 2002. Molecular mapping of major genes and quantitative trait loci determining lowering time in response to photoperiod in barley. Plant Breeding 12: 129-132. Dubcovsky, J. C. Chen, and L. Yan. 2005. Molecular characterization of the allelic variation at the VRN-H2 vernalization locus in barley. Mol. Breeding. 15:395-407. Fowler, D.B., G. Breton, A.E. Limin, S. Mahfoozi, and F. Sarhan. 2001. Photoperiod and temperature interactions regulate low-temperature-induced gene expression in barley. Plant Physiol. 127:1676-1681. Fowler, D.B., G. Breton, A.E. Limin, J.T. Ritchie. 1999. Low-temperature tolerance in Cereals: Model and Genetic Interpretation. Crop Sci. 39:626-633. Francia, E., F. Rizza, L. Cattivelli, A.M. Stanca, G. Galiba, B. Tóth, P.M. Hayes, J.S. Skinner, N. Pecchioni. 2004. Two loci on chromosome 5H determine low temperature tolerance in the new ‘winter’ x ‘spring’ (‘Nure’ x ‘Tremois’) barley map. Theor. Appl. Genet. 108: 670-680. Hayes, P.M., A. Castro, L. Marquez-Cedillo, A. Corey, C. Henson, B.L. Jones, J. Kling, D. Mather, I. Matus, C. Rossi, and K. Sato. 2003. Genetic Diversity for Quantitatively Inherited Agronomic and Malting Quality Traits. In R. Von Bothmer, H. Knupffer, T. van Hintum, and K. Sato (ed.). Diversity in Barley. Elsevier Science Publishers, Amsterdam. He, Z., Q. Zhu, T. Dabi, D. Li, D. Weigel, and C. Lamb. 2000. Transformation of rice with the Arabidopsis floral regulator LEAFY causes early heading. Transgenic Res. 9:223-227. Holland, J.B., V.A. Portyanko, D.L. Hoffman, and M. Lee. 2002. Genomic regions controlling vernalization and photoperiod responses in oat. Theor. Appl. Genet. 105:113-126. Karsai, I., K. Meszaros, L. Lang, P,M. Hayes, and Z. Bedo. 2001. Multivariate analysis of traits determining adaptation in cultivated barley. Plant Breeding 120:217-222. 28 Karsai, I., P. M. Hayes, J. Kling, I. A. Matus, K. Meszaros, L. Lang, Z. Bedo , and K. Sato. 2004. Genetic variation in component traits of heading date in Hordeum vulgare subsp. spontaneum accessions characterized in controlled environments. Crop Sci. 44:1622–1632. Karsai, I., P. Szűcs, , K. Meszaros, T. Filichkin, P. M. Hayes, L. Lang, and Z. Bedo. 2005. The VRN-H2 locus is a major determinant of flowering time in a facultative × winter growth habit barley (Hordeum vulgare L.) mapping population. Theor. Appl. Genet. 110:1458-1466. Large, E.C. 1954. Growth stages in cereals. Illustrations of the Feekes’ scale. Plant Path. 3:128-129. Laurie, D. 1997. Comparative genetics of flowering time. Plant Mol Bio. 35:167-177. Loukoianov, A., L. Yan, A. Blechl, A. Sanchez and J. Dubcovsky. 2005. Regulation of VRN-1 vernalization genes in normal and transgenic polyploid wheat.Plant Physiol. 2005. 138(4):2364-2373 Mahfoozi, S., A. E. Limin, P. M. Hayes, P. Hucl, and D. B. Fowler. 2000. Influence of photoperiod response on the expression of cold hardiness in wheat and barley. Can. J. Plant Sci. 80:721-724. Mahfoozi, S., A. E. Limin and D. B. Fowler. 2001. Influence of Vernalization and Photoperiod Responses on Cold Hardiness in Winter Cereals. Crop Sci 41:1006-1011 Skinner J.S., J. von Zitzewitz, P. Szűcs, L. Marquez-Cedillo, T. Filichkin, K. Amundsen, E. Stockinger, M.F. Thomashow, THH Chen, and P.M. Hayes. 2005. Structural, Functional, and Phylogenetic Characterization of a Large CBF Gene Family in Barley. Plant Mol. Bio. 59:533-551. Szűcs, P., I. Karsai, J. von Zitzewitz, K. Mészáros, L.L.D. Cooper, Y.Q. Gu, T.H.H. Chen, P.M. Hayes, and J.S. Skinner. 2006. Positional relationships between photoperiod response QTL and photoreceptor and vernalization genes in barley. Theor. Appl. Genet. 112:1277-1285. von Zitzewitz, J., P. Szűcs, J. Dubcovsky, L. Yan, E. Francia, N. Pecchioni, A. Casas, T.H.H. Chen, P. M. Hayes, and J. Skinner. 2005. Structural and functional characterization of barley vernalization genes. Plant Mol. Bio. 59:449-467. Yan, L., A. Loukoianov, G. Tranquilli, M. Helguera, T. Fahima, and J. Dubcovsky. 2003. Positional cloning of the wheat vernalization gene VRN1. PNAS. 100 (10): 6263-6268. 29 Yan, L., A. Loukoianov, A. Blechl, G. Tranquilli, W. Ramakrishna, P. SanMiguel, J.L. Bennetzen, V. Echenique, and J. Dubcovsky. (2004) The wheat VRN2 gene is a flowering repressor down-regulated by vernalization Science 303 (5664) : 1640-1644 Yan, L., J. von Zitzewitz, J. Skinner, P.M. Hayes, and J. Dubcovsky. 2005. Molecular characterization of the duplicated meristem identity genes HvAP1a and HvAP1b in barley. Genome. 48:905-912. 30 Acknowledgements I would like to sincerely thank Dr. Patrick Hayes, Head of the Oregon State Barley Project. His knowledge, persistence, and patience have helped me enormously throughout this project. I would like to thank Dr. Peter Szucs for all the time and expertise with molecular genetics that he put into this project. He always had time for questions and was eager to help. I would like to thank Ann Corey for the help and guidance with the field portion of this project. I would like to thank others in the OSU Barley Project including Tanya Filichkina and Kelly Richardson for always making time to answer laboratory questions. I would like to thank Tom Chastain for his help with this project and for being an excellent advisor and mentor. I would like to thank Wanda Crannell for the guidance throughout the Bioresource Research degree program and for keeping me motivated during difficult times. I would like to thank the Crop and Soil Science Department at OSU for making this academic experience not only interesting and fun, but a family affair. Thank you. 31